


Abstract
Recent clinical data indicate that tumor hypoxia negatively affects the treatment outcome of both radiotherapy and surgery in various cancers, emphasizing the need for noninvasive detection of tumor hypoxia. Several studies have shown an increased uptake of FDG in hypoxic regions of xenografts, suggesting the use of PET with FDG as a potential technique. In this study, we examine the mechanism underlying the hypoxia-induced increase of FDG uptake in the human breast carcinoma cell line MCF7. Methods: The uptake of 3H-FDG into MCF7 cells was determined after incubation under hypoxic (0% oxygen) or normoxic conditions, with or without redox agents, for varying time periods. In addition, the effects of the redox agents on the glucose transporter activity and the hexokinase activity were determined independently, and the effects of hypoxia on glucose transporter protein and hexokinase levels were assessed. Results: A more than twofold increase (2.53 ± 0.79; P < 0.005) in 3H-FDG uptake was observed under hypoxic conditions, but no changes in the cellular levels of glucose transporter proteins or hexokinase were observed. A reducing agent, dithiothreitol (DTT), also caused an increase in 3H-FDG uptake but failed to affect uptake under hypoxic conditions. This indicates that the mechanisms by which hypoxia and DTT affect 3H-FDG uptake might be the same. The oxidizing agent p-chloromercuribenzenesulfonic acid (pCMBS) had no effect on 3H-FDG uptake under normoxic conditions but counteracted the effect of hypoxia. DTT caused an increase in glucose transporter activity, whereas it had no effect on hexokinase activity. pCMBS had no effect on either glucose transporter activity or hexokinase activity. Conclusion: The hypoxia-induced increase in 3H-FDG uptake in MCF7 cells is the result, in part, of an increase in glucose transporter activity resulting from the modification (reduction) of thiol group(s) in the glucose transport protein(s). Modulation of hexokinase activity is probably not involved in the hypoxia-induced increase in 3H-FDG uptake in these cells.
Clinical data indicate that the presence of hypoxic regions negatively affects the treatment outcome of both radiotherapy and surgery in a variety of tumors (1–4). Thus, there is significant potential for therapeutic benefit if one could noninvasively diagnose tumor hypoxia and, correspondingly, use these data to define high-risk populations and modify therapy. PET imaging with 18F-FDG has been suggested as a potential technique for this purpose (5).
Several in vivo and in vitro studies indicate that the uptake of FDG is increased in hypoxic cells or prenecrotic (and probably hypoxic) areas in xenografts (5–8). The mechanisms underlying this increased uptake remain unclear. In one of the in vitro studies, a hypoxia-induced increase in the cellular level of glucose transporters (GLUTs) has been suggested as a possible explanation (5).
It has also been suggested that, under hypoxic conditions, the cellular redox status shifts to a more reduced state. This might result in modification (reduction) of thiol groups of cysteine residues in proteins, potentially altering their activity (9,10). Here we present data suggesting that this redox modulation is (in part) the mechanism underlying the hypoxia-induced increase in FDG uptake, that is, increased GLUT activity as a result of reduction of protein-thiol groups.
In animal tumor models (and probably in human tumors) both chronic and transient hypoxia occurs. Chronic hypoxia pertains to tumor cells that remain under hypoxic conditions for long durations as a result of their distance from the blood supply. Transient hypoxia is the result of sporadic or intermittent constriction of tumor blood vessels and can render tumor regions that were previously oxic acutely hypoxic (11,12). The experiments presented here were performed under conditions of acute hypoxia (up to 4 h) and will be more representative for tumor areas that experience transient hypoxia.
MATERIALS AND METHODS
Induction of Hypoxia
Hypoxia was induced using a modification of the method described by Ling et al. (13,14). For experiments, 1 × 106 cells were seeded as a monolayer into 15-cm2 glass flasks in medium containing 25 mmol Hepes (pH 7.4). To induce hypoxia, the flasks were sealed with soft rubber stoppers through which 19-gauge stainless-steel needles were inserted for equilibration with humidified N2 (<0.002% O2). After the 1-h equilibration period, during which the cells were maintained at 0°C to prevent uptake of 3H-FDG, the cells were rapidly returned to physiological temperatures (by immersing the flasks in a 37°C waterbath for 2 min) and then placed in a 37°C incubator. No uptake of FDG was observed during the equilibration period. Control experiments showed that the incubation at 0°C and subsequent rewarming to 37°C had no effect on FDG uptake in normoxic cells (data not shown). Oxygen measurements inside the culture flasks, using an Oxylite-4000 pO2 meter (Oxford Optronics, Oxford, UK), were performed to confirm hypoxia. In addition, radiation survival experiments were performed using hypoxic and normoxic cells. The oxygen enhancement ratios (OERs) observed in these experiments (n = 3) were between 2.8 and 3.0, indicating that the cells were radiobiologically hypoxic (data not shown).
3H-FDG Uptake
For the measurement of 3H-FDG uptake, cells were incubated with 1.5 μCi of 3H-FDG (60 Ci/mmol; American Radiolabeled Chemicals Inc., St. Louis, MO) in 1 mL medium for various periods. Subsequently, the cells were rinsed with cold phosphate-buffered saline (PBS) and lysed with 1 mL 0.3N NaOH/1% sodium dodecyl sulfate (SDS). The lysate was transferred into a liquid scintillation vial and neutralized with 3N HCl (0.1 mL), and the protein content was determined (using the bicinchoninic acid protein assay kit; Pierce, Rockford, IL). 3H-FDG activity was determined by liquid scintillation counting, and expressed as dpm/mg protein. Statistical analysis of the experimental results was performed using a paired t test.
GLUT Activity
[3H]3-O-methylglucose ([3H]3-O-MG) transport into cells was used as a measure for the glucose transporting capability of cells (15). The [3H]3-O-MG uptake in cells was assayed as described for 3H-FDG uptake, except for the tracer and incubation time. [3H]3-O-MG (80 Ci/mmol) was from New England Nuclear, Boston, MA.
Hexokinase Activity
Total hexokinase activity was measured using the method of Viñuela et al. (16) as modified by Waki et al. (15). The cells were scraped off, suspended in solution I (50 mmol triethanolamine, 5 mmol MgCl2;pH 7.6), and homogenized. Supernatants, containing free and mitochondria-bound hexokinase, were obtained by centrifuging the homogenates at 1000 g at 4°C for 5 min and were analyzed as follows. Three milliliters of solution I containing 0.5 mmol glucose, 5 mmol adenosine triphosphate (ATP), 0.25 mmol reduced nicotinamide adenine dinucleotide phosphate (NADPH), and 6 U glucose-6-phosphate dehydrogenase were preincubated at 20°C for 5 min. The supernatants were then added and the mixture further incubated at 20°C. After 5 and 20 min, samples of the reaction mixture were taken and the absorption measured at 340 nm. The differences in absorbance were calculated and the hexokinase activity determined from an NADPH standard curve. One unit of hexokinase activity will phosphorylate 1 μmol of glucose per minute at pH 7.6 and 20°C. Aliquots were removed to measure the protein content of the lysates, and the activity was expressed as milliU/mg protein.
Western Analysis
Total protein extracts of cells incubated under hypoxic conditions, and control cells were prepared in cold SDS sample buffer (2% SDS, 10% glycerol, and 50 mmol tris[hydroxymethyl]aminomethane [Tris]; pH 6.8) containing protease (0.5 mmol phenylmethylsulfonyl flouride [PMSF] and 1X Complete [Boehringer GmbH, Mannheim, Germany]) and proteasome (10 μmol MG132) inhibitors. The levels of hypoxia-inducible factor (HIF)-1α, HIF-1β, hexokinase II, GLUT1, and GLUT3 proteins were analyzed by separation of the proteins by SDS-polyacrylamide gel electrophoresis followed by Western analysis using standard protocols (17,18). AntiHIF-1α and antiHIF-1β antibodies were from Novus Biologicals (Littleton, CO), antihexokinase II antibody was from Santa Cruz Biotechnology Inc. (Santa Cruz, CA), and antiGLUT1 and antiGLUT3 antibodies were from Alpha Diagnostics International (San Antonio, TX). HRP-conjugated second antibodies (Boehringer Mannheim, Indianapolis, IN) and the Enhanced Chemoluminescence system (Amersham Life Science, Piscataway, NJ) were used to visualize the proteins.
RESULTS
Hypoxia-Induced Increase of 3H-FDG Uptake
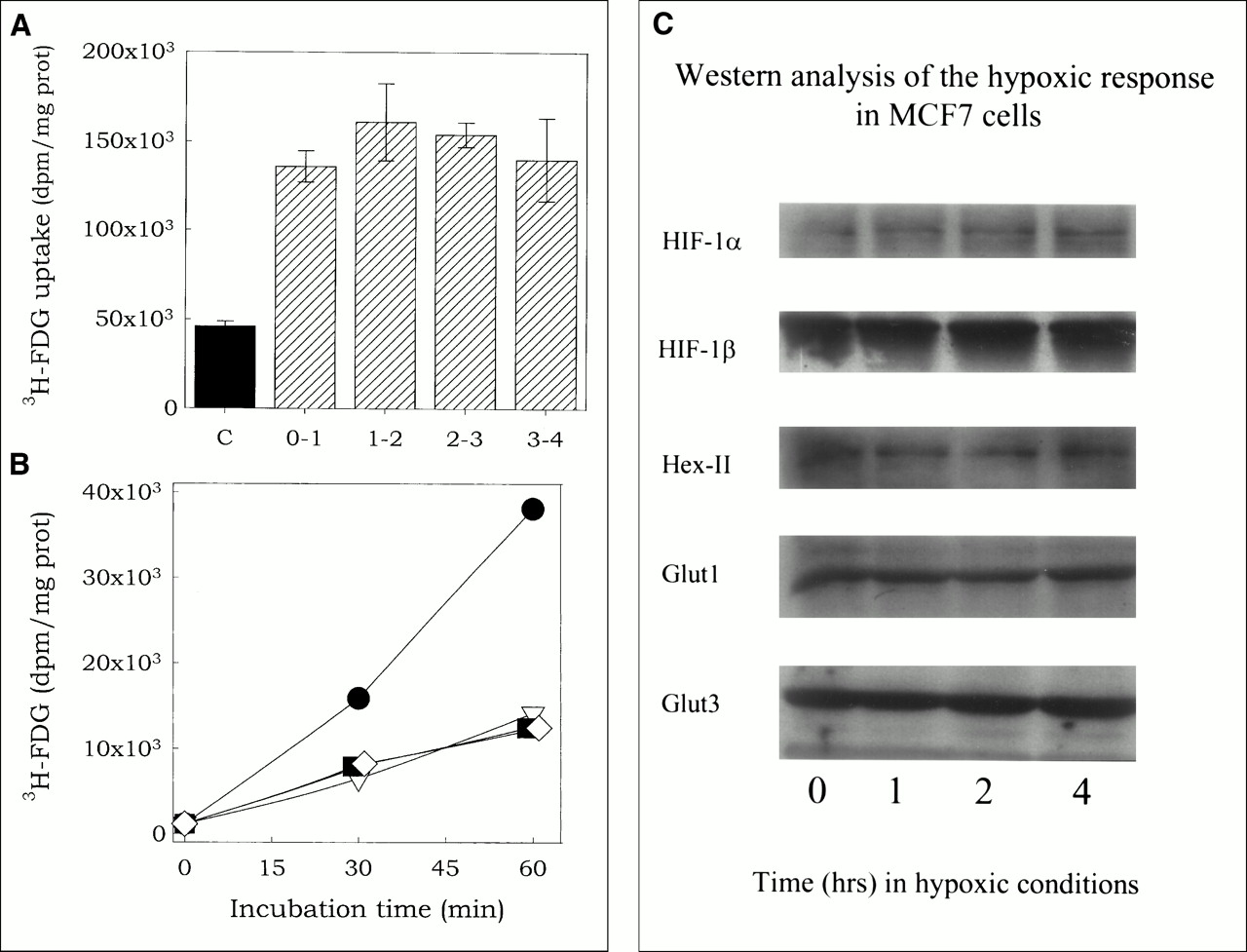
3H-FDG uptake was significantly higher in MCF7 cells incubated under hypoxic condition (<0.002% O2) than in oxygenated cells (21% O2). The increase in 3H-FDG uptake at 1 hr, averaged over nine experiments, was 2.53 ± 0.79-fold (P < 0.005). Figure 1A presents the results from a representative experiment on 3H-FDG uptake, showing an increased uptake in hypoxic cells (hatched bars) compared with oxygenated cells (solid bar). No differences in the rate of uptake were observed during the first 4 h of incubation under hypoxic conditions. Other data (not shown) indicated that the increase is observable after 15 min of incubation (earliest timepoint tested).

Hypoxia-induced increase in 3H-FDG uptake. (A) 3H-FDG uptake in cells under hypoxic (0% O2; hatched bars) or normoxic (21% O2; solid bar [C]) conditions. Cells were preincubated under hypoxic conditions for various periods (0–3 h), 3H-FDG was added without disrupting hypoxic conditions (by injection through the stopper), and uptake was determined 1 h later. For example, in column labeled 3-4, 3H-FDG was added after 3 h and uptake determined at 4 h after the induction of hypoxia. In this experiment, uptake of 3H-FDG was 3.21 (±0.25)-fold higher in hypoxic cells than in normoxic cells. (B) Cells were incubated for 1 h under hypoxia, followed by 0-h (squares) or 1-h (diamonds) incubation in air and then assessed for 3H-FDG uptake as function of time under normoxic conditions. 3H-FDG uptake in cells incubated under hypoxic (circles) and normoxic (triangles) conditions was measured for comparison. (C) Western analysis of the cellular HIF-1α, HIF-1β, hexokinase II, GLUT1 and GLUT3 levels in cells incubated under hypoxic conditions for various periods.
Figure 1B shows the effect of reoxygenation of hypoxic cells on 3H-FDG uptake. Compared with the rate in hypoxic cells, the rate of 3H-FDG uptake in aerated cells that were first incubated in hypoxia (circles) for 1 h followed by 0 (squares) or 1 h (diamonds) at normoxic conditions was significantly lower, and, in fact, was the same as that for cells incubated under normoxic conditions (triangles) without the hypoxia exposure. This rapid decay of the effect of hypoxia, in combination with the rapid appearance of the hypoxic effect, indicates that hypoxia-induced increase in 3H-FDG uptake is not mediated through de novo synthesis of enzymes involved in 3H-FDG uptake or retention. Western analysis showed no increase in the levels of the hypoxia-inducible proteins HIF-1α, HIF-1β, GLUT1, GLUT3, or hexokinase II in our MCF7 cells during incubation under hypoxic conditions for up to 4 h (Fig. 1C). The absence of HIF-1α accumulation during hypoxia is not the result of an HIF-1α deficiency, because a 4-h incubation of the MCF7 cells with 10 μmol 2-2 dipyridyl (an iron chelator) induces a strong increase in the cellular HIF-1α level (data not shown). The lack of p53 accumulation during hypoxia (not shown) is probably the result of the failure of hypoxia to induce HIF-1α accumulation in this cell line, because a strong increase in the cellular p53 level is observed within 4 h of treatment of the cells with ionizing radiation (data not shown). Thus, in these MCF7 cells, hypoxia induces a rapid increase in 3H-FDG uptake, which is independent of de novo protein synthesis.
In principle, an enhancement of the rate of 3H-FDG uptake with a fixed cellular GLUT and hexokinase content can result from translocation of GLUTs from sites within the cell to the plasma membrane or increased activity of the existing enzymes through either an increase in affinity or activation of “dormant” or “masked” enzymes (19). Translocation of GLUTs from intracellular sites to the plasma membrane does not seem to be involved in the mechanism underlying the observed hypoxia-induced increase in 3H-FDG uptake, because the presence of Brefeldin A, an inhibitor of GLUT translocation to the plasmamembrane, does not interfere with the effect of hypoxia on 3H-FDG uptake (data not shown). Therefore, an increase in the activity of the GLUTs or hexokinase as a result of post-translational modification or of a hypoxia-induced change in the cellular redox status seems to be the more likely mechanism underlying the hypoxia-induced increase in 3H-FDG uptake.
Effect of Redox Agents on 3H-FDG Uptake
By adding dithiothreitol (DTT, a thiol reducing agent) or p-chloromercuribenzenesulfonic acid (pCMBS, an oxidizing agent) to the medium we altered the cells’ redox status to study the influence of redox on 3H-FDG uptake. As shown in Figure 2, the effect of DTT on 3H-FDG uptake is concentration-dependent, negligible at 0.1 or 1 mmol but inducing a 1.54 ± 0.44-fold (P < 0.05; n = 5) increase at 10 mmol. Preincubation (1 hr) of the cells with 10 mmol DTT did not enhance this effect. In contrast, pCMBS had no effect on 3H-FDG uptake (Fig. 3). These data suggest that the reduction of protein thiol groups might be at least part of the mechanism underlying the hypoxia-induced increase in 3H-FDG uptake.
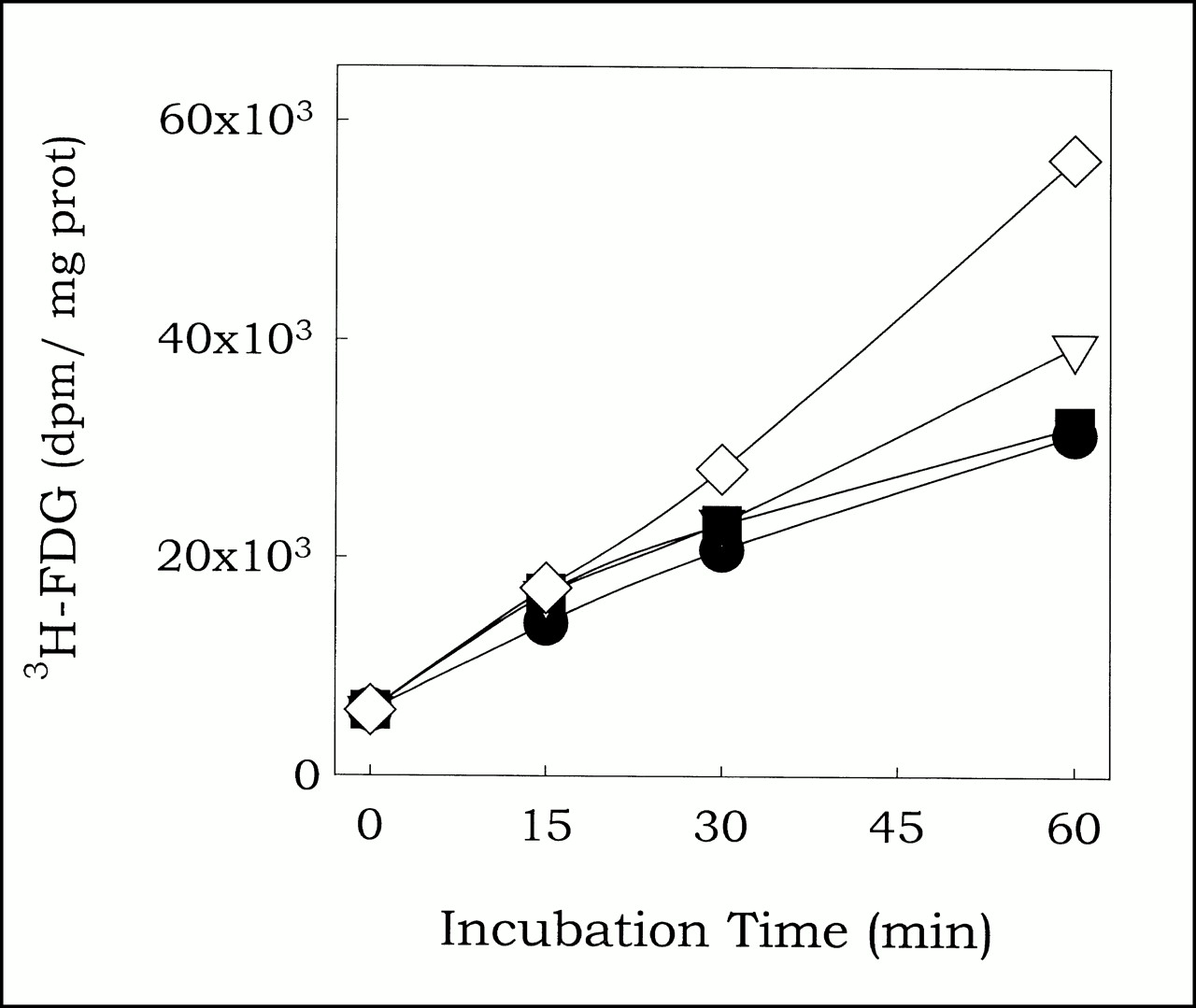
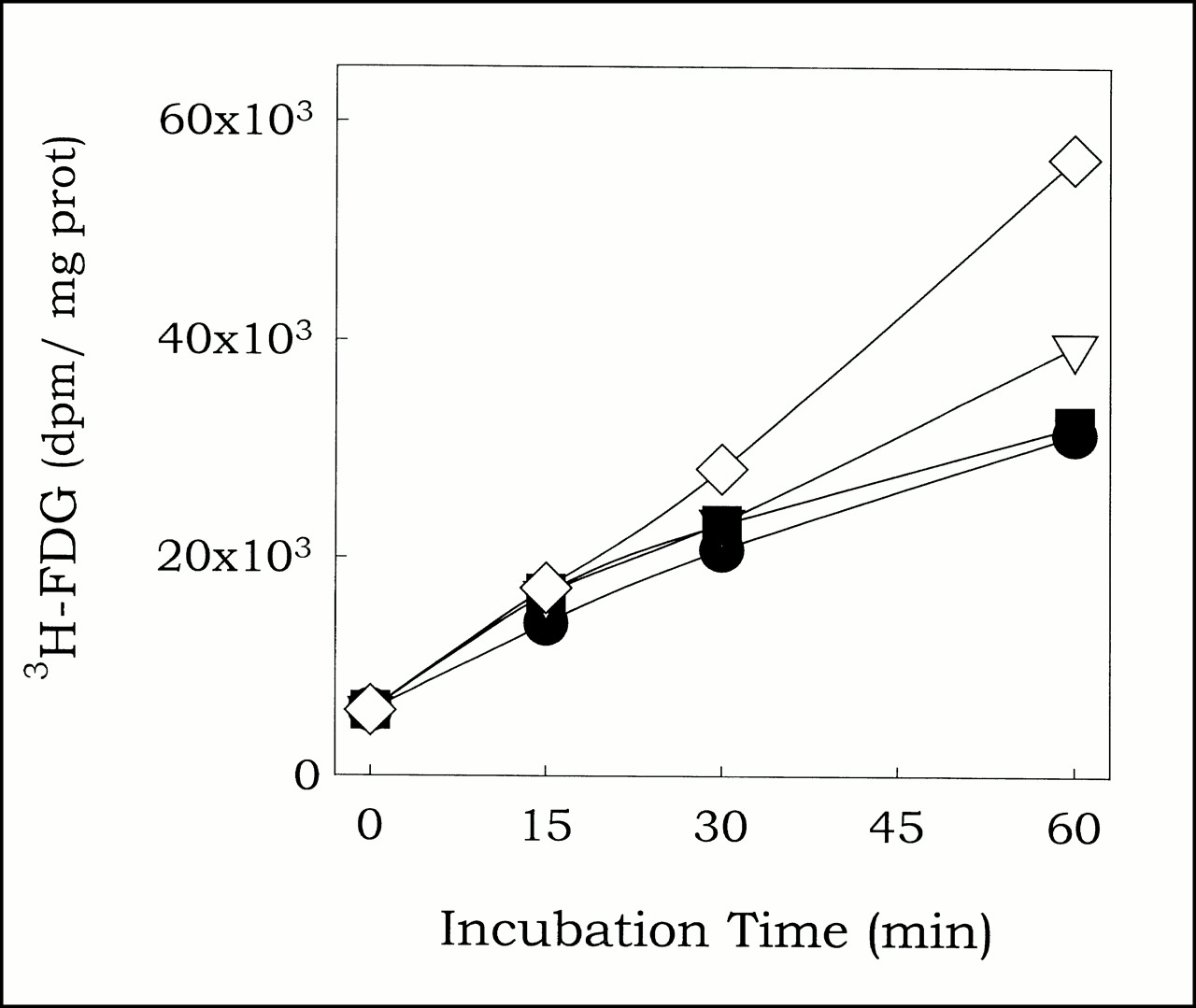
To assess effect of modulation of the redox status on 3H-FDG uptake, cells were incubated under normoxic conditions with 3H-FDG and graded doses of DTT for various periods. Subsequently, cells were lysed and 3H-FDG uptake determined. Relative to control (0 mmol, circles), DTT concentrations of 0.1 mmol (triangles) and 1 mmol (squares) had no effect on 3H-FDG uptake, whereas 10 mmol DTT (diamonds) induced 1.54 ± 0.44-fold increase (P < 0.05, n = 5).
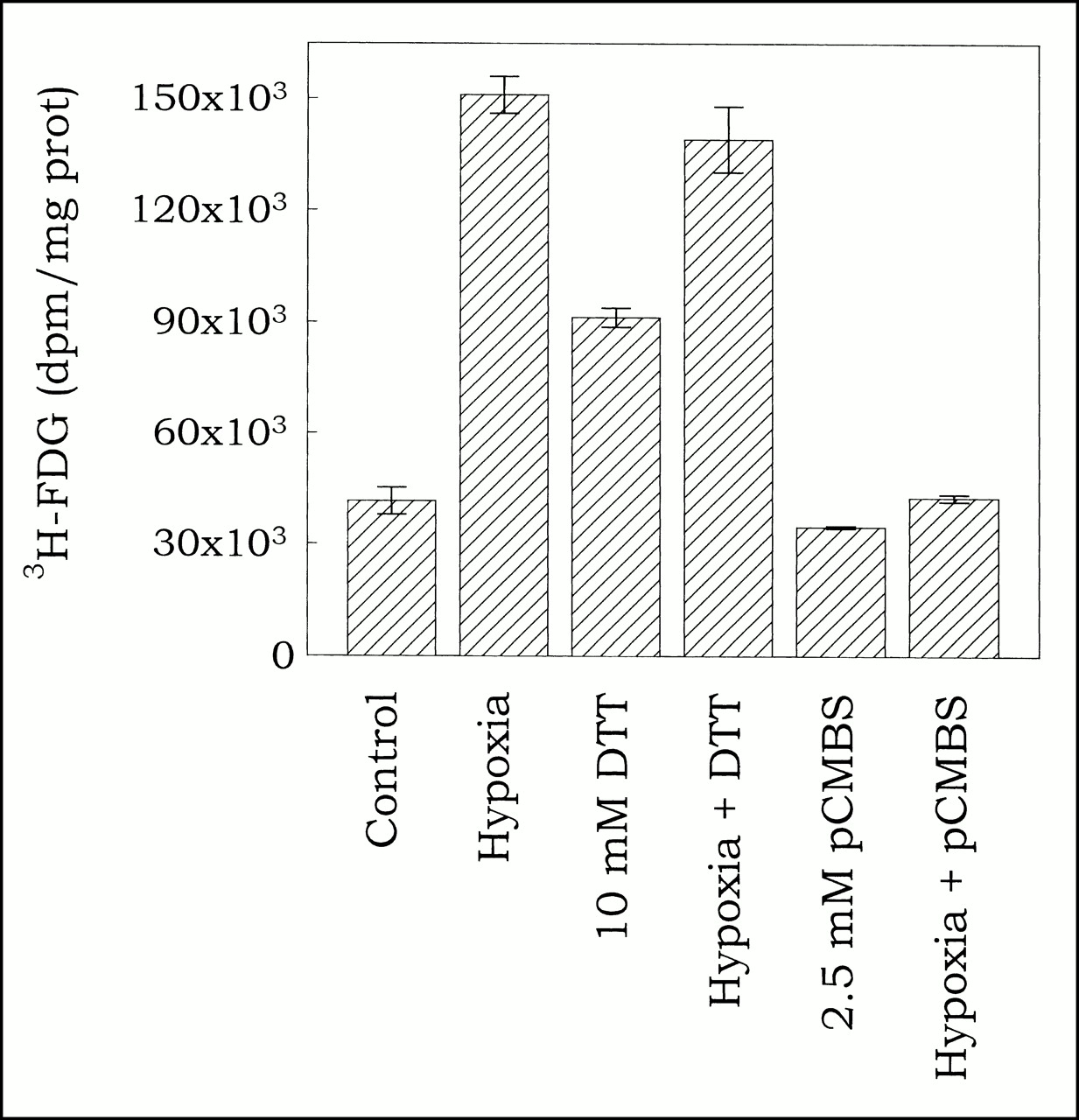
To test hypothesis that DTT and hypoxia affect 3H-FDG uptake through same mechanism, effect of combining hypoxia with DTT or pCMBS was determined. Uptake was determined in cells incubated for 1 h with 3H-FDG under normoxic or hypoxic conditions with or without 10 mmol DTT or 2.5 mmol pCMBS.
Interaction of DTT/pCMBS and Hypoxia
To further examine the mechanism(s) underlying the effect of hypoxia and DTT on 3H-FDG uptake (presumed to be the reduction of protein thiol), we assessed the effect of combining DTT with hypoxia and pCMBS with hypoxia. If DTT and hypoxia act through different mechanisms, their effects on 3H-FDG uptake should be additive (as has been shown for the effects of insulin and hypoxia on glucose uptake [19]). On the other hand, if DTT and hypoxia act through the same mechanism, their combined effect on 3H-FDG uptake should be much less than the additive effect of the separate treatments. As shown in Figure 3, this is indeed the case. Hypoxia by itself induced a 3.63 ± 0.12-fold increase in 3H-FDG uptake compared with normoxic cells, whereas 10 mmol DTT induced a 2.20 ± 0.06-fold increase. Compared with hypoxia alone, the combination with DTT did not induce an additional increase in 3H-FDG uptake (3.35 ± 0.21-fold compared with normoxic cells or 0.92 ± 0.12-fold compared with hypoxic cells). The effect of hypoxia plus DTT is identical to the effect of hypoxia by itself, indicating that their cellular targets might be the same. On the other hand, pCMBS, which by itself had no effect on 3H-FDG uptake, almost completely inhibited the effect of hypoxia, suggesting that its mechanism of action (oxidation of protein thiol groups) is antagonistic to that of hypoxia. These results provide strong support for the hypothesis that reduction of protein thiol groups is at least part of the mechanism underlying the hypoxia-induced, protein-synthesis-independent increase in 3H-FDG uptake.
Effect of DTT on GLUT and Hexokinase
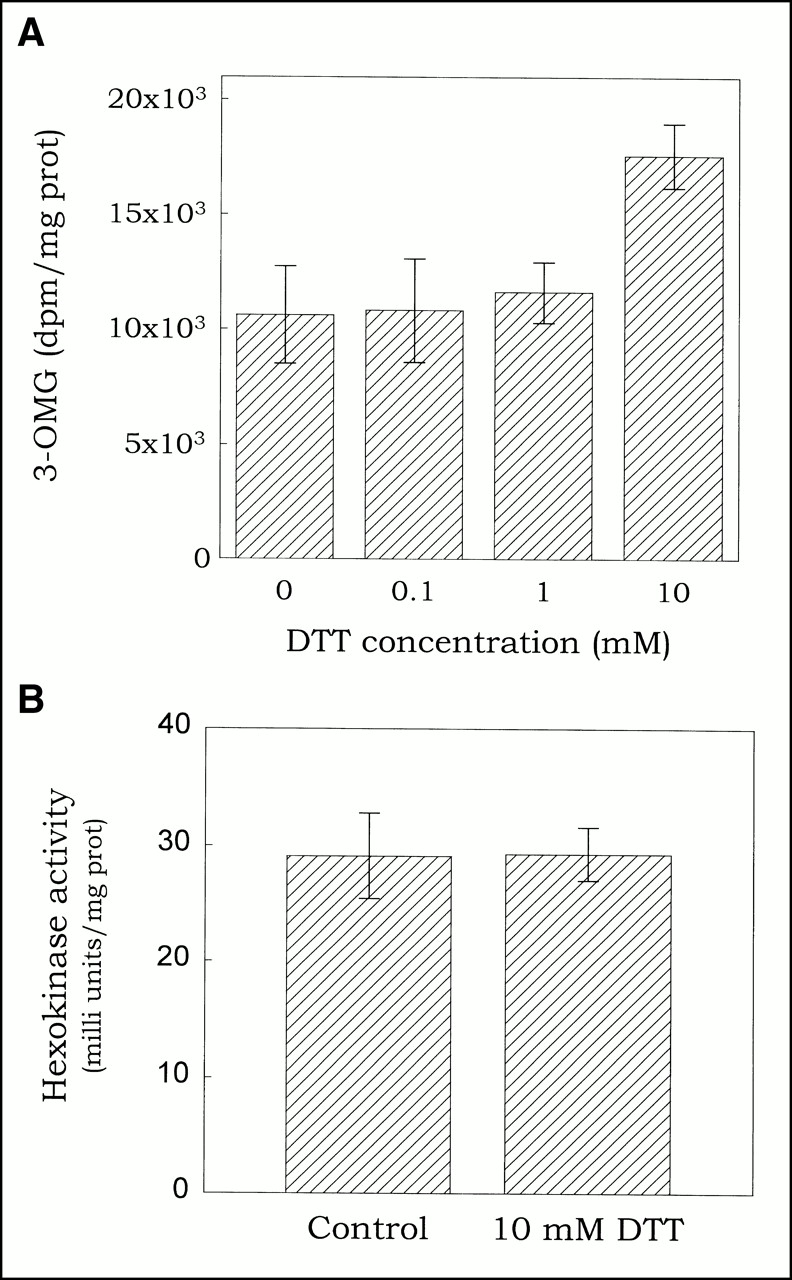
Cellular uptake of 3H-FDG occurs in two steps: translocation across the plasma membrane by GLUTs and phosphorylation by hexokinase. To determine which of these is affected by redox changes, we studied the effect of DTT on GLUT activity and hexokinase activity. Because cytochalasin B has been shown to compete with DTT for interaction with GLUT 1 (20), total, rather than cytochalasin B inhibitable, 3-O-methylglucose (3-OMG) uptake was used as a measure for GLUT activity. As shown in Figure 4A, the effect of DTT on GLUT activity was concentration-dependent, with no effect at 0.1 or 1 mmol but inducing a 1.47 ± 0.18-fold (P = 0.1; n = 2) increase in GLUT activity at 10 mmol. Thus, the effect of DTT on GLUT activity seems to mirror its effect on 3H-FDG uptake, both in concentration dependence and magnitude, suggesting that the two effects are related.

Effect of dithiothreitol on glucose transporter activity and hexokinase activity. (A) Cells were preincubated for 30 min with graded doses of DTT, and glucose transporter activity determined as 3-OMG uptake during 2 min. (B) Hexokinase captivity was determined spectrophotometrically in cell lysates using a glucose-6-phosphate dehydrogenase coupled assay.
In contrast, DTT (at 10 mmol) had no effect on hexokinase activity (Fig. 4B) and pCMBS (up to 2.5 mmol) had no significant effect on GLUT activity (data not shown). Thus, DTT (and probably hypoxia) seems to increase 3H-FDG uptake through modification of the GLUT activity.
DISCUSSION
Our data show that acute hypoxia increases 3H-FDG uptake in MCF7 cells. This effect develops rapidly on hypoxia induction, decays quickly on return to normoxic conditions, and is independent of de novo protein synthesis.
Although the MCF7 cells used in this study were not HIF-1α deficient, hypoxia failed to induce accumulation of HIF-1α. Consequently, no increase in the cellular levels of p53 (p53 accumulation is the result of stabilization of this protein by HIF-1α) or of the HIF-1α-regulated proteins GLUT 1, GLUT 3, or hexokinase II was observed. HIF-1α accumulation could be observed after treatment of the cells with the iron chelator 2-2 dipyridyl, indicating that the absence of hypoxia-induced HIF-1α accumulation is probably the result of a defect in the hypoxia-sensing signal transduction pathway in these cells.
Experiments conducted with the reducing agent DTT yielded results indicating that alterations in the cellular redox status may underlie the hypoxia-induced increase in 3H-FDG uptake. Furthermore, this increase appeared to be the result of an increase in GLUT activity, with no change in hexokinase activity. Others have reported that hypoxia shifts the cellular redox status to a more reduced state, resulting in the reduction of protein thiol groups (9,12). These observations suggest that hypoxia-induced modification (reduction) of the cysteine residues in the GLUT may be the mechanism underlying the hypoxia-induced increase in 3H-FDG uptake. The same mechanism has previously been suggested for the hypoxia-induced inhibition of K+ and Ca2+ channels (9). That the oxidizing agent pCMBS counteracts the effect of hypoxia on 3H-FDG uptake provides further evidence for the role of redox modulation of protein thiol groups in this phenomenon.
There is a second mechanism by which protein thiol groups may be reduced under hypoxic conditions. Oxygen (or reactive oxygen species [ROS]) can react directly with protein thiol groups. It has been shown that the activity of several transcription factors can be regulated by this mechanism, at least in vitro (21). In addition, it has been shown that membrane protein activity, such as glutamate and dopamine transport in rat striatal synaptosomes, can be modulated through interaction with oxygen or ROS (22,23). Thus, the reduced amount of oxygen and ROS (and the resulting shift of protein-thiol groups to a more reduced state) under hypoxic conditions may mediate the observed increase in 3H-FDG uptake.
An enhancement of transmembrane 3H-FDG transport without translocation of GLUTs to the plasma membrane may result either from increased affinity of GLUTs existing in the plasma membrane or activation of dormant or “masked” GLUTs preexisting in the plasma membrane (19). Currently, we cannot distinguish between these possibilities. Further studies on the kinetics of 3H-FDG uptake under hypoxic conditions will provide the answer to these questions, because the first possibility will result in a decreased Km value, whereas the second possibility will result in an increased Vmax for 3H-FDG transport.
Our data with DTT also indicate that the hypoxia-induced increase in 3H-FDG uptake is the result of increased GLUT activity, rather than hexokinase activity, suggesting that GLUT activity is rate-limiting for 3H-FDG and glucose uptake in MCF7 cells. This is in agreement with the data of Waki et al. (15), that among a panel of 16 tumor cell lines and one normal cell line, 2-deoxyglucose (2DG) uptake is governed by GLUT activity rather than hexokinase activity.
That GLUT activity was increased by DTT does not shed insight as to which GLUT is responsible for the redox-sensitive 3H-FDG uptake increase. It has been reported that in MCF7 cells only GLUT 1 and GLUT 3 are expressed (24). However, our result seems to argue against a role for GLUT 1, because it has been reported that GLUT 1 activity is inhibited by DTT (25,26) as well as by pCMBS (27) at concentrations similar to or lower than those used in our experiments. No data are available on the effects of these agents on GLUT 3 activity.
The kinetics for the hypoxia-induced increase in 3H-FDG uptake and our postulate of the underlying mechanism are different from those observed by Clavo et al. (5,7) in human melanoma and human adenocarcinoma cell lines. In their cell lines, 1.5-h incubation in hypoxia was required before an increase in 3H-FDG uptake (attributed to an increased membrane expression of GLUT 1) could be detected. Whereas the results of Clavo et al. (5,7) suggest a role for GLUT 1 in the mechanism underlying the hypoxia-induced increase in 3H-FDG uptake, our results seems to argue against GLUT 1 involvement in MCF7 cells. It is possible that differences in GLUT species expressed in the various cell lines can account for the disparity in results.
CONCLUSION
3H-FDG uptake is increased in MCF7 cells under conditions of acute hypoxia. This increase is not associated with a detectable increase in the cellular levels of GLUT protein(s) or hexokinase. Experiments using redox modulators (either alone or in combination with hypoxia) indicate that the hypoxia-inducible increase in 3H-FDG uptake is most likely the result of an increase in GLUT activity resulting from the modification (reduction) of thiol group(s) in the glucose transport protein(s). Modulation of hexokinase activity is probably not involved in the hypoxia-induced increase in 3H-FDG uptake in these cells.
Acknowledgments
This work was partially supported by U.S. Army Grant BC980411.
Footnotes
Received Mar. 3, 2000; revision accepted Jul. 28, 2000.
For correspondence or reprints contact: Paul Burgman, PhD, Department of Medical Physics, Memorial Sloan-Kettering Cancer Center, 1275 York Avenue, New York, NY 10021.
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Concomitant [18F]F-FAZA and [18F]F-FDG Imaging of Gynecological Cancer Xenografts: Insight into Tumor Hypoxia
- GOT1 primes the cellular response to hypoxia by supporting glycolysis and HIF1{alpha} stabilisation
- Relationships between SUVmax of lung adenocarcinoma and different T stages, histological grades and pathological subtypes: a retrospective cohort study in China
- Glucose-lactate metabolic cooperation in cancer: insights from a spatial mathematical model and implications for targeted therapy
- Preclinical Assessment of Efficacy of Radiation Dose Painting Based on Intratumoral FDG-PET Uptake
- Preclinical Kinetic Analysis of the Caspase-3/7 PET Tracer 18F-C-SNAT: Quantifying the Changes in Blood Flow and Tumor Retention After Chemotherapy
- The Reverse Warburg Effect and 18F-FDG Uptake in Non-Small Cell Lung Cancer A549 in Mice: A Pilot Study
- Biological Correlation of 18F-FDG Uptake on PET in Pulmonary Neuroendocrine Tumors
- PET Imaging of Tumor Hypoxia Using 18F-Fluoroazomycin Arabinoside in Stage III-IV Non-Small Cell Lung Cancer Patients
- Tumor Microenvironment-Dependent 18F-FDG, 18F-Fluorothymidine, and 18F-Misonidazole Uptake: A Pilot Study in Mouse Models of Human Non-Small Cell Lung Cancer
- Correlation Between 18F-FDG Uptake on PET and Molecular Biology in Metastatic Pulmonary Tumors
- Biologic Correlation of 2-[18F]-Fluoro-2-Deoxy-D-Glucose Uptake on Positron Emission Tomography in Thymic Epithelial Tumors
- High 18F-FDG Uptake in Microscopic Peritoneal Tumors Requires Physiologic Hypoxia
- 18F-FDG Uptake in Lung, Breast, and Colon Cancers: Molecular Biology Correlates and Disease Characterization
- Chemotherapy Response Monitoring of Colorectal Liver Metastases by Dynamic Gd-DTPA-Enhanced MRI Perfusion Parameters and 18F-FDG PET Metabolic Rate
- Impact of Angiogenesis-Related Gene Expression on the Tracer Kinetics of 18F-FDG in Colorectal Tumors
- In vivo19F Magnetic Resonance Spectroscopy and Chemical Shift Imaging of Tri-Fluoro-Nitroimidazole as a Potential Hypoxia Reporter in Solid Tumors
- Combining Radioimmunotherapy with Antihypoxia Therapy 2-Deoxy-D-Glucose Results in Reduction of Therapeutic Efficacy
- Positron Emission Tomography-Based Molecular Imaging in Human Cancer: Exploring the Link between Hypoxia and Accelerated Glucose Metabolism
- Hypoxia and Glucose Metabolism in Malignant Tumors: Evaluation by [18F]Fluoromisonidazole and [18F]Fluorodeoxyglucose Positron Emission Tomography Imaging
- Cellular Origin and Molecular Mechanisms of 18F-FDG Uptake: Is There a Contribution of the Endothelium?
- Analysis of the Regional Uptake of Radiolabeled Deoxyglucose Analogs in Human Tumor Xenografts
- Enhancing Targeted Radiotherapy by Copper(II)diacetyl- bis(N4-methylthiosemicarbazone) Using 2-Deoxy-D-Glucose
- Repetitive Supply-Demand Ischemia with Dobutamine Increases Glucose Uptake in Postischemic and Remote Myocardium
- Effect of Respiratory Gating on Quantifying PET Images of Lung Cancer