


Abstract
The goal of this study was to determine whether myocardial glucose uptake after repetitive ischemia differs in response to coronary occlusion–reperfusion versus supply–demand ischemia induced by dobutamine. Although glucose metabolism is increased after myocardial ischemia, the metabolic effect of supply–demand ischemia induced by dobutamine may increase glucose metabolism within remote myocardium. This would make it difficult to discriminate postischemic from remote myocardium with glucose tracers. Methods: Eighteen swine with a hydraulic occluder and flow probe on the circumflex artery underwent repetitive ischemia. In group 1 (n = 9), the circumflex artery was occluded, whereas in group 2 (n = 9), circumflex flow was decreased by 30% before dobutamine (40 μg/kg/min intravenously). Each pig underwent 15 min of ischemia, twice per day for 5 d. Echocardiography and PET to determine myocardial glucose (18F-FDG) uptake were performed after final ischemia, and tissue was later analyzed for activation of Akt, p38 mitogen-activated protein, and adenosine monophosphate (AMP) kinase. Results: Wall thickening in the circumflex region was lower than in remote regions in both groups. 18F-FDG uptake in the circumflex region was similar in groups 1 and 2 (0.22 ± 0.03 and 0.23 ± 0.04 μmol/min/g, respectively; not statistically significant). In the remote region, 18F-FDG uptake was lower than in the circumflex region in group 1 (0.14 ± 0.03 μmol/min/g; P < 0.05) but was similar to that in the circumflex region in group 2 (0.20 ± 0.03 μmol/min/g; not statistically significant). AMP kinase activity in the remote region was significantly lower than in the circumflex region in group 1 but was similar to that in the circumflex region in group 2. Conclusion: Unlike repetitive coronary artery occlusion–reperfusion, repetitive supply–demand ischemia with dobutamine alters glucose uptake within the remote myocardium, possibly as a result of AMP kinase activation. Clinically, these data suggest that 18F-FDG studies have a limited role in discriminating postischemic from remote myocardium after dobutamine stress.
Ischemic reperfused myocardium demonstrates a sustained increase in glucose uptake when compared with remote tissue (1–3). In the absence of reperfusion, myocardium in the distribution of coronary artery disease also demonstrates a transient increase in glucose uptake relative to remote myocardium, after either exercise (4) or dipyridamole stress (5). Although dobutamine stress would be expected to induce the same response within postischemic myocardial tissue, sustained effects of adrenergic receptor stimulation may affect glucose metabolism in the remote myocardium independent of insulin-mediated changes in phosphoinositol 3-kinase (6,7). Understanding the effects of dobutamine on glucose metabolism in remote tissue is important for defining the utility of glucose metabolic imaging studies for discriminating postischemic from remote myocardial regions.
In the present study, we hypothesized that postischemic myocardial glucose metabolism is similar in models of repetitive occlusion–reperfusion and repetitive supply–demand mismatch induced by high-dose dobutamine. We speculated that glucose metabolism in the remote myocardium differs between models, however. PET was used to estimate changes in myocardial glucose transport and phosphorylation with the glucose analog 18F-FDG during fasting conditions, in 2 swine models of ischemia.
MATERIALS AND METHODS
This study was performed under the guidance of the animal care committee at the VA Medical Center, Minneapolis, MN, and conformed with Guide for the Care and Use of Laboratory Animals (8).
Experimental Protocol
Domestic pigs (30–35 kg) of either sex were used for these studies. After an overnight fast, they were sedated with xylazine (2 mg/kg intramuscularly) and Telazol (combination of teletamine HCl and zolazepam HCl; Fort Dodge Laboratory, Ft. Dodge, IA) (4 mg/kg intramuscularly), ventilated, and anesthetized with isoflurane (1%). The left neck and chest were prepped and draped, and lines were placed in the external jugular vein and internal carotid artery and subcutaneously tunneled through the skin. A left thoracotomy was performed in the 5th intercostal space, and the circumflex artery was dissected free and instrumented with a Doppler flow probe (2.5 mm) and hydraulic occluder. Incisions were closed, and sterile dressings were applied. The animals were allowed to recover for 72 h, and a prophylactic antibiotic (cephazolin, 1 g intramuscularly) was given twice a day for 2 d.
Before ischemia, the animals were sedated with Telazol (4 mg/kg intravenously). Two groups of 9 animals were assigned: In group 1 (occlusion–reperfusion group), the circumflex artery was completely occluded for 15 min, followed by full reperfusion. In group 2 (supply–demand ischemia group), mean circumflex flow by Doppler flow probe was decreased by ∼30% of baseline, and dobutamine (40 μg/kg/min) was infused intravenously for 15 min. Coronary flow was kept constant throughout the 15 min of dobutamine infusion and, by gradual release of the hydraulic occluder after 15 min, was allowed to return to normal without a reactive hyperemic phase. Preliminary data from pilot studies showed that this dose of dobutamine increases blood flow in remote myocardium by ∼2-fold, resulting in a relative flow reduction in the circumflex region of ∼60%–70% during dobutamine stress. In all animals, regional myocardial ischemia was induced twice per day for 5 d, during continuous monitoring of heart rate, blood pressure, and electrocardiograms.
Animals fasted on the final day. One hour after the final episode of ischemia, they were ventilated and anesthetized with isoflurane (1%). Regional myocardial function was measured with 2-dimensional echocardiography (model 5500; Hewlett-Packard Co., Palo Alto, CA), and wall thickening was measured at the right parasternal short-axis view in the posterior (circumflex) and anterior (remote) walls. Wall thickening was computed as the difference between end-systolic and end-diastolic wall thickness and expressed as a percentage of end-diastolic thickness, using a Hewlett-Packard software program. End-diastole and end-systole were defined as the onset of the QRS configuration and the frame with the smallest chamber size, respectively. The animals were transferred to the PET suite and positioned on the table. Images were acquired with an ECAT 953B/31 (CTI, Knoxville, TN/Siemens Medical Systems, Inc., Hoffman Estates, IL). Myocardial blood flow was measured with 13N-ammonia (555 MBq), infused intravenously over 20 s during acquisition of dynamic scans. The scanning protocol consisted of one 30-s, twelve 10-s, two 30-s, three 60-s, and one 900-s frames. Fifty minutes later, 18F-FDG (222 MBq) was infused, and dynamic scans were acquired over the next 40 min. The 18F-FDG scanning protocol included twelve 10-s, six 30-s, four 60-s, three 120-s, three 300-s, and one 600-s frames. After the PET scan, the animals were returned to the surgical suite and a midline sternotomy was performed. Transmural biopsies were obtained from the circumflex and remote regions using a modified variable-speed drill. Specimens were transferred to liquid nitrogen–cooled 2-methyl butane and stored for later analysis. The animals were then sacrificed.
PET Analysis
Multiple circular regions of interest (ROI) were chosen from 5–10 transverse planes from the initial myocardial blood flow images and were saved for analysis of the blood flow and 18F-FDG scans. The ROIs were selected from the postischemic (circumflex) and remote (left anterior descending) regions from each plane and were placed in the center of the myocardial wall to avoid spillover and partial-volume effects. For analysis of the studies, ∼5 ROIs were obtained in both the circumflex and the remote territories, and a circular region was obtained from the largest portion of the left ventricular cavity for arterial input. Time–activity curves for each ROI were obtained from both the dynamic 13N-ammonia scans and the 18F-FDG scans.
For estimation of regional myocardial blood flow, we applied a 3-compartment model (9). Compared with other methods, the advantage of this model is that the partial-volume and spillover effects are both implicitly accounted for by the introduction of a total blood volume parameter into the model equation. This approach has been proven to be independent of the region size provided that the ROI contains only blood and myocardial tissue. Arterial input function was derived from an ROI carefully placed in the center of the left ventricle, which minimizes the spillover effect from the myocardium to the blood pool. Metabolite correction for the input function was not performed because of its negligible effects. Myocardial blood flows were obtained by a nonlinear least squares fitting to the model equation using the input function and 18 tissue samples acquired in the first 6 min of the dynamic scan.
For the determination of myocardial glucose uptake, regional time–activity curves were generated from each of the myocardial ROIs and from the left ventricular cavity as arterial input. Patlak plots were generated from the time–activity curves from the left ventricular cavity and each myocardial ROI. Values were then averaged for the postischemic circumflex and remote regions. The model has been previously described and is based on a 3-compartment model (10). Briefly, the Patlak plot defines a constant (K) that incorporates the forward (k1) and reverse (k2) rate constants from plasma to tissue and the phosphorylation (k3) constant. The formula is expressed as K = (k1 × k2)/(k2 + k3). The dephosphorylation constant (k4) is assumed to be zero. The metabolic rates of glucose uptake were determined from arterial plasma glucose samples, and the calculations assumed a lumped constant of 0.67. Although fasting conditions were expected to cause a low metabolic rate of glucose uptake, they avoided high insulin levels, which may cause an underestimation in glucose uptake based on its effects on the lumped constant. The latter is the factor relating differences in transport and phosphorylation between deoxyglucose analogs and glucose.
Tissue Analysis
Immunoprecipitation was performed for all activity kinase assays. Equivalent amounts of protein were diluted with 2% sodium dodecylsulfate Laemmli sample buffer. Sodium dodecylsulfate–polyacrylamide gel electrophoresis was performed, and gels were transferred to polyvinylidene fluoride membranes (Bio-Rad Laboratories Inc., Hercules, CA). Membranes were blocked with 5% milk in tris-buffered saline in 0.1% polysorbate for 2 h at 25°C, followed by an overnight incubation in primary antibodies (phospho-specific Akt serine 473, protein kinase substrate, activating transcription factor 2 [ATF2] substrate, and phospho-specific mitogen-activated protein kinase–activated protein kinase 2 [MAPKAPK2]) at 4°C. After a brief wash in tris-buffered saline in 0.1% polysorbate, membranes were incubated with an antirabbit-linked horseradish peroxidase antibody for 1 h at 25°C. Phosphorylation state or activities were detected with chemiluminescence using the ECL Plus kit (Amersham Biosciences, Piscataway, NJ). Quantification was done with an imaging densitometer. Values from the postischemic circumflex region were normalized to the remote region and were expressed as arbitrary units.
For glycogen, tissue was dissolved in 30% KOH, precipitated with ethanol, and hydrolyzed using amyloglucosidase. The glucose residues were measured with a nicotinamide adenine dinucleotide phosphate–linked spectrophotometric method by use of glucose 6-phosphate dehydrogenase and hexokinase. For enzyme activities, frozen tissue was pulverized to a fine powder in a liquid nitrogen–cooled stainless steel percussion mortar. Tissue was then homogenized in 60% glycerol, 10 mmol/L ethylenediaminetetraacetic acid, 50 mmol/L KF, at pH 7.0, and 1:2 (wt/vol) in a Potter-Elvehjem homogenizer. Homogenization was completed at 0°C using 10 mmol/L ethylenediaminetetraacetic acid, 50 mmol/L KF, at pH 7.0, and 1:10 (wt/vol). The homogenate was centrifuged at 12,000g for 10 min at 4°C, and the supernatant was used for assay. Frozen tissue was homogenized, and hexokinase activity was estimated by determining the rate of nicotinamide adenine dinucleotide phosphate production measured over 20 min at 340 nm and 30°C.
Immunohistochemical slides were prepared from transmural sections of frozen samples, cut on a cryostat microtome, mounted on Superfrost Plus slides (Erie Scientific Co., Portsmouth, NH), air dried for 2 min, and stored at −80°C. The slides were fixed in methanol at −20°C for 20 min and were blocked with 1% bovine serum albumin in tris-buffered saline in 0.1% polysorbate. For co-localization of glucose transporter (GLUT) 4 with nuclei, the slides were incubated for 2 h at 37°C with polyclonal rabbit GLUT 4 antisera. Secondary fluorescein-labeled donkey antirabbit IgG and propidium iodide were used, and the slides were incubated at 37°C for 1 h followed by a 1-h wash in triethanolamine-buffered saline. They were mounted with Vectashield (Vector Laboratories, Burlingame, CA) and examined with confocal microscopy.
Statistical Analysis
Results are expressed as mean ± SE. Differences between the postischemic circumflex and remote regions were tested at the P < 0.05 level of significance with a paired Student t test.
RESULTS
Systemic hemodynamics at baseline and during the final period of ischemia are shown in Table 1. At baseline, there were no differences in heart rate or blood pressure. During ischemia, however, dobutamine in group 2 animals increased heart rate and systolic blood pressure by ∼40%–50%. One hour after the final period of ischemia, regional wall thickening was lower in the circumflex region than in the remote region in both groups (Table 1).
Systemic Hemodynamics at Baseline and During Final Episode of Ischemia and Regional Myocardial Wall Thickening by Echocardiography 1 Hour After Final Episode of Ischemia
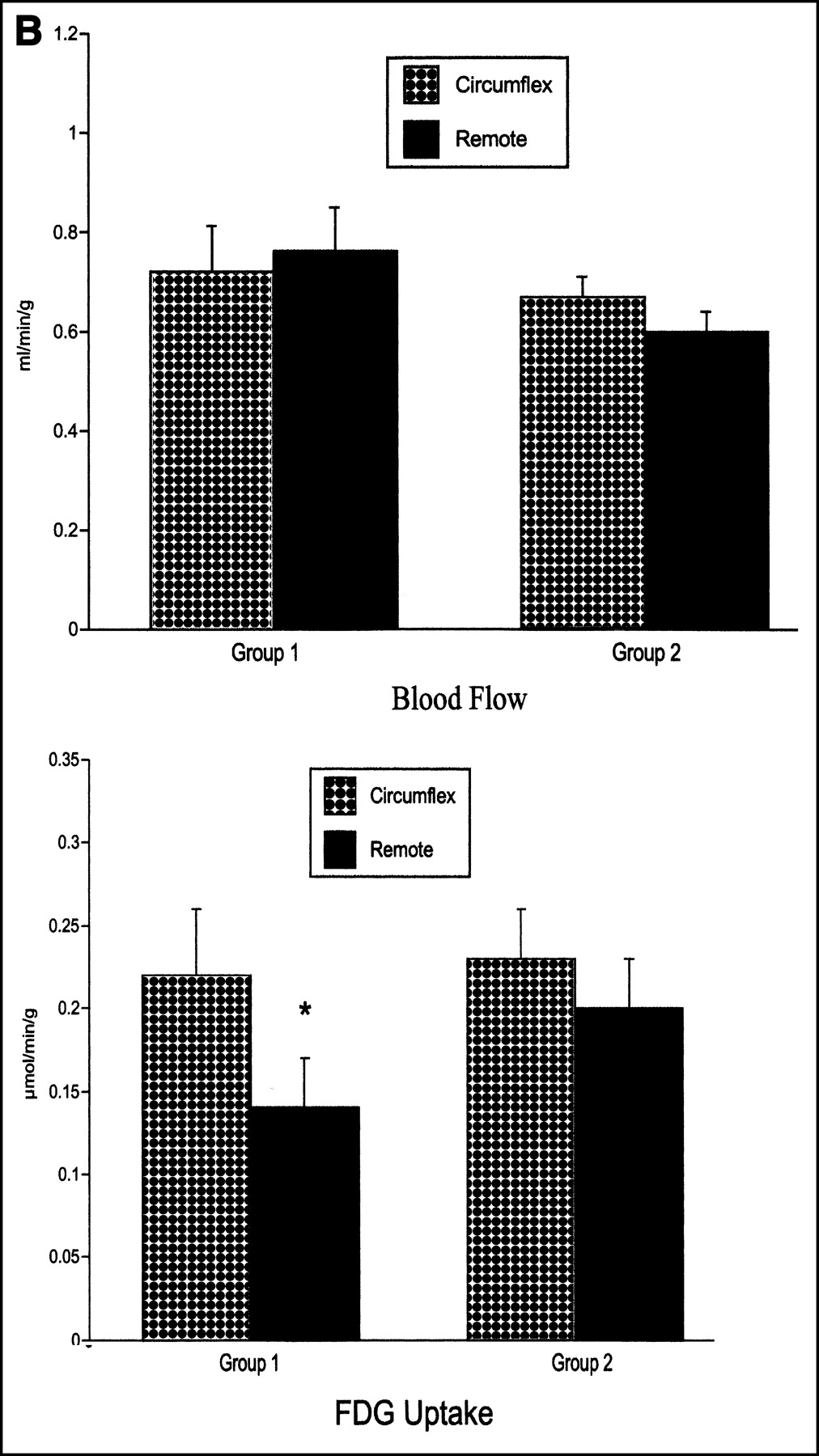
Myocardial blood flow in the postischemic circumflex region was similar to that in the remote region in both groups (Fig. 1). The metabolic rate of myocardial glucose uptake in the postischemic circumflex region was similar in both models of repetitive ischemia (0.22 ± 0.03 μmol/min/g and 0.23 ± 0.04 μmol/min/g from groups 1 and 2, respectively; not statistically significant). The metabolic rate of myocardial glucose uptake in the remote myocardium was lower than in the circumflex region in group 1 pigs (0.14 ± 0.03 μmol/min/g; P < 0.05 vs. circumflex region) but was similar to that in the circumflex region in group 2 pigs (0.20 ± 0.03 μmol/min/g) (Fig. 1). Plasma substrates, glycogen, and hexokinase activity were similar in both groups (Table 2).


(A) Transaxial images of myocardial blood flow (13N-ammonia; MBF) and 18F-FDG are shown in same plane, during basal, fasting condition, 2 h after repetitive ischemia. Arrows indicate postischemic circumflex region. Group 1 (n = 9) underwent complete circumflex artery occlusion with reperfusion, whereas group 2 (n = 9) underwent supply–demand ischemia, induced by 30% reduction in circumflex artery flow before infusion of dobutamine (40 μg/kg/min intravenously). All animals underwent 15 min of ischemia, twice per day for 5 d. (B) In group 1, myocardial 18F-FDG was higher in postischemic circumflex region than in remote regions, whereas in group 2, 18F-FDG was homogeneously distributed. Data are expressed as mean and SEM. *P < 0.05 versus circumflex region of same group.
Plasma Substrates, Glycogen, and Hexokinase Activity After Final Episode of Ischemia
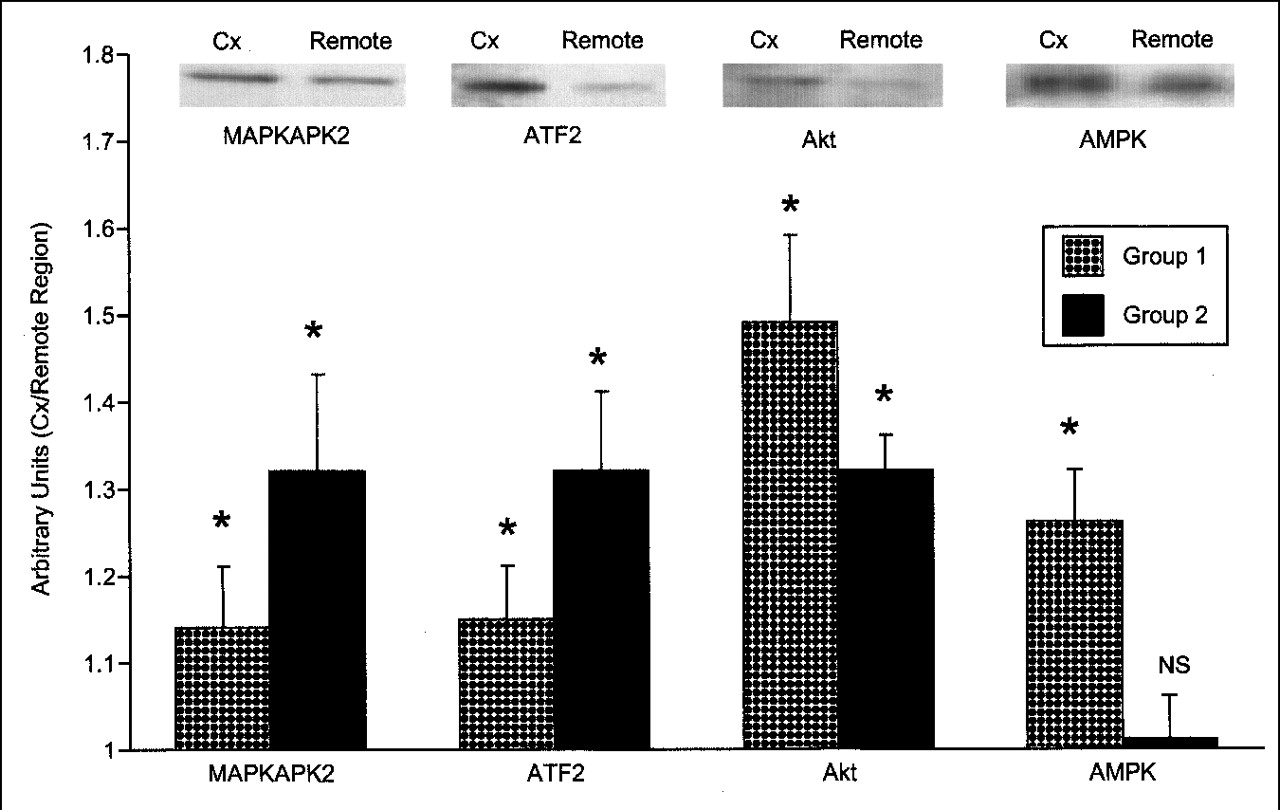
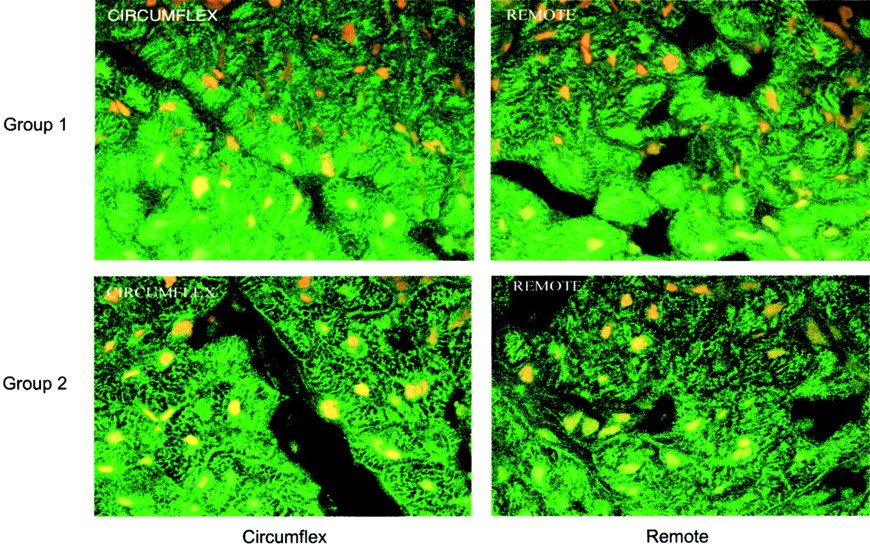
Activities for p38 mitogen-activated protein (MAP) kinase (MAPKAPK2 and ATF2 substrate assay), Akt, and adenosine monophosphate (AMP) kinase in the postischemic circumflex region were statistically compared with the corresponding remote region from each animal. Values from the circumflex region were then normalized to the remote regions and were expressed in arbitrary units. As shown in Figure 2, the postischemic circumflex region was higher than the remote region for all assays, except for the AMP kinase activity from group 2 pigs. No qualitative differences in regional myocardial GLUT 4 translocation were detected by immunohistochemistry in either group 1 or group 2 animals (Fig. 3). These data suggest that the sustained glucose metabolic effects of dobutamine in the remote myocardial region may relate to increased rates of transport through AMP kinase.

Activation of p38 MAP (MAPKAPK2 and ATF2 substrate assays), Akt, and AMP kinase are shown for groups 1 (n = 9) and 2 (n = 9) from tissue harvested after sacrifice. Values from postischemic circumflex region were normalized to remote regions and expressed as arbitrary units. Representative Western gels are shown from group 1 animals. Of note, activities of kinases were higher in postischemic circumflex region from all groups, except for AMP kinase from group 2. These findings show that dobutamine-activated AMP kinase is higher within remote regions from group 2. Data are expressed as mean and SEM. *P < 0.05, circumflex versus remote region of same group. AMPK = AMP kinase; Cx = circumflex.

Immunohistochemical staining of myocardial GLUT 4 transport protein (green) and nuclei (yellow) (×200) are shown from postischemic circumflex and remote regions from groups 1 and 2. No differences in GLUT 4 translocation were apparent.
DISCUSSION
The principal findings of this study are 2-fold: First, during fasting, postischemic myocardial glucose uptake after repetitive coronary artery occlusion is similar to that of repetitive supply–demand ischemia induced by high-dose intravenous dobutamine. Second, myocardial glucose uptake in the remote myocardium is higher after repetitive supply–demand ischemia, demonstrating that dobutamine induces a sustained metabolic effect on remote myocardial tissue. This may relate to a sustained effect on glucose transport through AMP kinase activation. Clinically, these data suggest that glucose metabolic studies with 18F-FDG kinetics have a limited role in discriminating postischemic from remote myocardium after dobutamine stress testing.
In the present study, animals were kept fasting to increase the likelihood of detecting differences in 18F-FDG kinetics between postischemic and remote myocardium. Plasma substrates, tissue glycogen, and hexokinase activity did not differ between groups 1 and 2 and therefore cannot explain the observed differences in 18F-FDG uptake in the remote myocardium. In anesthetized animals, regional myocardium subjected to sustained ischemia and complete reperfusion demonstrates increased glucose uptake relative to remote myocardium (1–3). Although the mechanism of the enhanced glucose uptake still remains speculative, reactive hyperemia does not appear to be a prerequisite. In patients, for instance, after a brief episode of either exercise stress (4) or pharmacologic stress with dipyridamole (5), myocardium in the distribution of severe coronary artery disease also shows a transient increase in glucose uptake, when compared with remote myocardium. In experimental models of regional myocardial ischemia and reperfusion, differences in 18F-FDG uptake within postischemic and remote myocardium are more striking at 24 h than early after reperfusion (2,3). The mechanism for this remains unclear but may represent either increased workload in remote regions early after reperfusion or an effect of high catecholamines after the prolonged anesthesia in animal preparations. The latter concept would be consistent with the present findings, which indicate that adrenergic stimulation can attenuate differences in 18F-FDG uptake in postischemic and remote tissue, by virtue of changes in the remote myocardium.
An alternative explanation for the relatively increased uptake of 18F-FDG at 24 h after reperfusion is that hexokinase activity may be activated 24 h after reperfusion and not early after reperfusion. In a similar model of ischemia–reperfusion induced by 10 min of repetitive ischemia, we found that hexokinase activity in the postischemic region exceeded that in the remote regions 24 h after reperfusion, at a time when 18F-FDG uptake and glycogen storage were also higher than in remote regions (11). The values for the hexokinase activity were nearly twice as high as those of the present study, despite similar conditions of fasting. In the present study, hexokinase and glycogen levels were similar in the postischemic and remote regions. It is possible that differences between studies relate to the temporal differences in recovery, relative to either glycogen or regional wall thickening.
Repetitive ischemia was induced over 5 d. Over this period, it is unlikely that the total content of the myocardial glucose transporters is increased (11,12). Prolonged, moderate ischemia does provide a strong stimulus for increasing glucose uptake, presumably by enhancing the translocation of the GLUT transporters from cytoplasmic stores to active sites on the sarcolemma (13). Because this process occurs in the absence of insulin (i.e., during fasting conditions), the signal involved with this process acts independently of phosphoinositol 3-kinase (14). Within ischemic-preconditioned myocardium, the mitogen-activated protein kinase, p38 MAP kinase, phosphorylates its major substrate MAPKAPK2 and increases glucose transport by facilitating the translocation of GLUT 4 (7). In the present study, p38 MAP kinase activation was higher in the postischemic region in both groups, using assays for both MAPKAPK2 phosphorylation and ATF2 substrate. Similarly, Akt activation was higher in the postischemic circumflex regions from both models, but neither could explain the differences in 18F-FDG uptake within the remote regions. The differences in myocardial 18F-FDG uptake in these 2 models of repetitive ischemia is most likely a result of the sustained metabolic effects of dobutamine on glucose transport in the remote region. AMP-activated protein kinase has been shown to increase glucose uptake in response to adrenergic stimulation, though the effects were more dependent on α-receptor than β-receptor stimulation (6). In the present study, AMP kinase activation in the remote region was lower than in the postischemic region of group 1 animals but was similar to that in the postischemic region of group 2 animals. AMP kinase activity in the postischemic region of group 1 animals was ∼30% higher than in remote regions—a magnitude similar to that of the regional differences in 18F-FDG uptake. The absence of regional differences in AMP kinase activity in group 2 animals provides a plausible mechanism for the finding that myocardial 18F-FDG uptake was more homogeneously distributed after the supply–demand ischemia induced by dobutamine.
Whether intracellular translocation of the GLUT transporters is a prerequisite for a sustained increased in glucose uptake after ischemia is not clear. In the present study, qualitative estimates of GLUT 4 translocation by immunohistochemistry did not differ between the postischemic circumflex and remote regions from either model of ischemia. Previous models of postischemic myocardium have shown that a persistent increase in glucose uptake is evident at a time when translocation of the GLUT transporters is not present (15). One mechanism for a sustained increase in glucose uptake in the absence of GLUT translocation is that ischemia or reperfusion can increase the activity of existing membrane-bound GLUT protein by reducing thiol groups within the glucose transport protein (16). Therefore, glucose transport activity can be increased, despite a normal content of membrane-bound protein transporter.
We used a lumped constant of 0.67 to adjust for differences in rates of transport and phosphorylation between molecules of glucose and deoxyglucose (10). Although prior studies have shown that this constant varies when plasma concentrations of insulin and substrate are altered (17,18), animals were kept fasting in the present study to provide a more stable metabolic environment. More recent studies suggest that a lumped constant of unity might be more accurate, when differences in arterial input between the plasma and whole-blood compartments are considered (19). Although use of a different lumped constant would change the absolute measurements of myocardial glucose uptake, the magnitude of the regional differences in the present study would not be different.
CONCLUSION
After repetitive supply–demand ischemia with dobutamine, myocardial glucose metabolism differs from that of repetitive occlusion–reperfusion, by virtue of alterations within the remote myocardium. The sustained metabolic effects of dobutamine may be a result of activation of AMP kinase. Clinically, these data suggest that 18F-FDG studies have a limited role in discriminating postischemic from remote myocardium after dobutamine stress, even in fasting conditions.
Acknowledgments
This work was supported in part by a Merit Review Grant from the U.S. Department of Veterans Affairs.
Footnotes
Received Apr. 4, 2002; revision accepted Aug. 8, 2002.
For correspondence or reprints contact: Edward O. McFalls, MD, PhD, Cardiology (111C), VA Medical Center, 1 Veterans Dr., Minneapolis, MN 55417.
E-mail: mcfal001{at}tc.umn.edu




{kind=link}
{kind=link}
{kind=link}
{kind=link}