


Abstract
Human somatostatin (SS) receptor (sst)–positive tumors can be visualized by gamma camera scintigraphy after the injection of [111In-diethylenetriaminepentaacetic acid (DTPA)-d-Phe1] octreotide. Uptake of [111In-DTPA-d-Phe1]octreotide is dependent on sst-mediated internalization of the radioligand by the tumor cells. Human sst-positive tumors frequently express multiple sst subtypes. In vitro studies have demonstrated that the 5 sst subtypes (sst1–5) differentially internalize sst-bound ligand. The present study was performed to evaluate the role of sst2 in vivo in determining the uptake of [111In-DTPA-d-Phe1]octreotide, as well as of the more “universal” ligand [111In-DTPA]SS-14, by sst-positive organs expressing multiple sst subtypes. Methods: Wild-type and sst2 knockout mice (n = 4 per treatment group) were injected intravenously with 1 MBq (0.1 μg) [111In-DTPA-d-Phe1]octreotide or [111In-DTPA]SS-14. After 24 h, the animals were sacrificed and radioactivity in the organs under investigation was determined. In addition, the sst subtype messenger RNA (mRNA) expression pattern in these organs was determined by reverse transcriptase polymerase chain reaction (RT-PCR) analysis. Results: RT-PCR analysis demonstrated the presence of all 5 sst subtype mRNAs in the adrenals and pituitary of wild-type mice but no sst2 in the knockout mice. The thymus expressed mRNA for sst2 and sst4 mRNA in wild-type mice, whereas no sst2 was detected in knockout mice. In wild-type mice, the in vivo uptake values (in percentage injected dose per gram of tissue) of [111In-DTPA-d-Phe1]octreotide for the pituitary, adrenals, pancreas, and thymus amounted to 1.2 ± 0.2, 0.26 ± 0.03, 0.18 ± 0.03, and 0.30 ± 0.05, respectively, in wild-type mice. Compared with wild-type mice, sst2 knockout mice had dramatically lower uptake values in these organs—lower by 97%, 83%, 96%, and 94%, respectively (P < 0.01 vs. wild type). Comparable differences in the uptake of radioactivity between wild-type and knockout mice were found using [111In-DTPA]SS-14 as the radiotracer. Interestingly, in some organs expressing sst2 mRNA (liver, muscle, and peripheral blood mononuclear cells), no specific binding of [111In-DTPA-d-Phe1]octreotide or [111In-DTPA]SS-14 to sst in vivo was found, suggesting that the sst2 protein expression level was very low in these tissues. Conclusion: The uptake of [111In-DTPA-d-Phe1]octreotide and [111In-DTPA]SS-14 in sst-positive organs is determined predominantly by sst2.
Human somatostatin (SS) receptors (ssts) are expressed in a variety of organ systems, including the brain, anterior pituitary gland, pancreas, thyroid gland, and gastrointestinal tract (1). Tumors arising from sst-positive cells in these organs contain a high density of ssts (2–5). This high density of ssts on human tumors forms the basis for the successful and wide use of radiolabeled SS-analogs in the detection of human sst-positive tumors and their metastases by sst scintigraphy (4,6). Moreover, preliminary phase I clinical trials using the β-radiation–emitting SS-analogs, such as [90Y-dodecanetetraacetic acid (DOTA),Tyr3]octreotide and [90Y-DOTA]lanreotide for sst-targeted radiotherapy of neuroendocrine tumors have been performed with promising results (7–12). The long residence of tumor-associated radioactivity after the administration of radiolabeled SS-analogs suggests that the radioligand is internalized after binding to its sst. In vitro studies have indeed confirmed sst-mediated internalization of radiolabeled SS-analogs by sst-expressing cell lines and primary cultures of human sst-positive tumors (13–17). Internalization of [111In-diethylenetriaminepentaacetic acid (DTPA)-d-Phe1]octreotide in vivo is also evidenced by our observations in rats, in which uptake of radioactivity in sst-positive organs, such as the pituitary gland and the pancreas, after the injection of the radiopharmaceutical can be prevented by injection of an excess of unlabeled octreotide up to 10 min after injection but not 20 min after injection. At that time, all radioactivity present in the sst-positive tissues probably reflects internalized radioligand (18). This process of sst-mediated internalization of radioisotopes coupled to SS-analogs will bring the radioisotope closer to its target, that is, the nucleus of the tumor cell. The currently used radiotherapeutics [90Y-DOTA,Tyr3]octreotide, [90Y-DOTA]lanreotide, and [177Lu-DOTA,Tyr3]octreotate (19) bind with high affinity to sst subtype 2 (sst2), with moderate affinity to sst subtype 5 (sst5), and with low affinity to sst subtype 3 (sst3). The binding affinity to sst subtypes 1 (sst1) and 4 (sst4) is very low (20). The majority of human sst-positive tumors frequently express multiple sst subtypes (3,5). The expression of multiple sst subtypes by human tumors makes it important to evaluate which sst subtypes are responsible for the uptake of radioactivity after the injection of radiolabeled SS-analogs. On the basis of the high binding affinity of [90Y-DOTA,Tyr3]octreotide and [90Y-DOTA]lanreotide to sst2 (20), as well as on the basis of a correlation between the relative uptake values of [111In-DTPA-d-Phe1]octreotide and sst2 messenger RNA (mRNA) expression (21,22), it can be suggested that this sst subtype plays an important role. Nevertheless, the involvement of sst5 and sst3 cannot be fully excluded. In fact, in vitro studies have demonstrated that both sst5 and sst3 internalize radiolabeled SS more efficiently than does sst2 (23). In order to determine whether sst5 and sst3 play a role in sst-mediated uptake of radiolabeled octreotide as well, we performed tissue distribution studies of uptake of radioactivity in sst-positive tissues after the injection of [111In-DTPA-d-Phe1]octreotide in wild-type and sst2 knockout mice. Uptake values were compared with the sst subtype expression pattern in these organs. For comparison, [111In-DTPA]SS-14 was used as a “universal” radiotracer.
MATERIALS AND METHODS
Animals
C57 black wild-type and sst2 knockout mice were kindly provided by Dr. James Schaeffer (Merck Laboratories). Establishment of these mice was described in detail previously (24). The animals were bred and maintained under specific pathogen-free conditions with free access to γ-irradiated pellet food (Hope Farms) and water acidified to pH 2.2 in the animal facilities of the Erasmus University Rotterdam. The experimental protocol was approved by the Animal Welfare Committee of the Erasmus Medical Center.
Preparation of Organs for RNA Isolation
After the mice were killed by CO2 asphyxiation, peripheral blood was collected by cardiac puncture into polypropylene tubes containing 100 international units of heparin (Leo Pharmaceuticals B.V.). Peripheral blood mononuclear cells (PBMC) were isolated by Ficoll gradient (density, 1.07 kg/L; Pharmacia) centrifugation according to the manufacturer’s protocol. PBMC from 2 mice were pooled, and aliquots of 106 cells were collected. The aliquots were lysed as described in detail elsewhere (25) and were stored at −80°C until RNA isolation. Within minutes after the mice were killed, the following organs were removed and snap-frozen in liquid nitrogen: brain, pituitary, pancreas, adrenals, thymus, spleen, liver, kidneys, and soft tissue (muscle). These organs were then stored at −80°C until further processing. For RNA isolation, the tissues were ground to powder, while frozen, using a mortar and pestle. Tissue powder was stored in ribonuclease-free containers at −80°C.
Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) Studies
RT-PCR was performed as described previously (25). Briefly, poly A+ mRNA was isolated using Dynabeads Oligo (dT)25 (Dynal AS) from powdered tissue (10 mg) or 106 cells. Complementary DNA (cDNA) was synthesized using the poly A+ mRNA captured on the Dynabeads Oligo (dT)25 as solid-phase and first-strand primer. Twenty microliters of the cDNA were used for each amplification by PCR using primer sets specific for mouse sst1–5, for SS, and for hypoxanthine-guanine phosphoribosyl transferase (hprt) as a control, as listed in Table 1. Mouse genomic DNA served as a positive control for sst1–5. For SS and hprt, cDNA of mouse brain RNA was used. The PCR was performed in a DNA thermal cycler with a heated lid (Perkin Elmer Cetus Instruments). After an initial denaturation at 94°C for 5 min, the samples were subjected to 40 cycles of denaturation at 94°C for 1 min, annealing for 2 min at 60°C, and extension for 1 min at 72°C. After a final extension for 10 min at 72°C, 10-μL aliquots of the resulting PCR products were analyzed by electrophoresis on 1.5% agarose gels stained with ethidium bromide. The identities of the products were confirmed by direct sequencing using an ABI Prism 3100 Genetic Analyzer (Applied Biosystems) according to the manufacturer’s protocol.
Primers Used for RT-PCR Analysis
sst-Binding Studies
Determination of the binding affinity profiles of SS-28, SS-14, and [DTPA]SS-14 for sst1–5 was performed by sst autoradiography on cryostat sections of membrane pellets of cells stably expressing sst1 or sst5 (CHO-K1) and sst2, sst3, and sst4 (CCL39 cells), as described in detail previously (20). Briefly, 20-μm-thick cryostat sections were incubated with [125I-Leu8,d-Trp22,Tyr25]SS-28 using increasing concentrations of unlabeled peptide ranging from 0.1 to 1,000 nmol/L. Concentrations resulting in 50% inhibition of specific binding (IC50) were calculated after quantification of the autoradiographic signal, as described in detail elsewhere (26).
Radiolabeling of [DTPA-d-Phe1]Octreotide and [DTPA,d-Ala1,d-Trp8,Tyr11]SS-14 ([DTPA]SS-14)
[DTPA-d-Phe1]octreotide, [DTPA]SS-14, and 111InCl3 (DRN 4901, 370 MBq/mL in HCl, pH 1.5–1.9) were obtained from Mallinckrodt. [DTPA]SS-14 was synthesized by one of us. [DTPA-d-Phe1]octreotide and [DTPA]SS-14 were 111In-labeled (labeling efficiency > 98%) as described in detail previously (27). Quality controls were performed as described elsewhere (27).
[111In-DTPA-d-Phe1]Octreotide Tissue Distribution Studies
Male mice under ether anesthesia were injected via the penis vein with 1 MBq (0.1 μg) of [111In-DTPA-d-Phe1]octreotide or [111In-DTPA]SS-14. This had been previously determined as being the optimal mass of peptide resulting in the highest uptake values (not shown). To determine specific binding to sst in vivo, a second group of mice was injected with the radiopharmaceutical in combination with 100 μg of unlabeled octreotide (Novartis Pharma) or SS-14 (Sigma Chemical Co.), respectively. The injection volume was kept constant at 0.2 mL per mouse. The mice were sacrificed by CO2 asphyxiation 24 h after injection of the radiopharmaceutical. Within minutes after the mice were killed, peripheral blood and organs of interest were removed, weighed, and collected into polyurethane tubes. Afterward, tissue and blood radioactivity was counted using an LKB-1282-Compugamma system (Wallac). Specific binding was determined by calculating the difference between uptake of radioactivity in the absence of competitor (total binding) and uptake of radioactivity in the presence of 100 μg of unlabeled octreotide or SS-14 (nonspecific binding), as described previously (28). Statistical analysis was performed using ANOVA followed by the Newman–Keuls multiple-comparison test. A P value of <0.05 was considered statistically significant.
RESULTS
mRNA Expression in Cells and Tissues
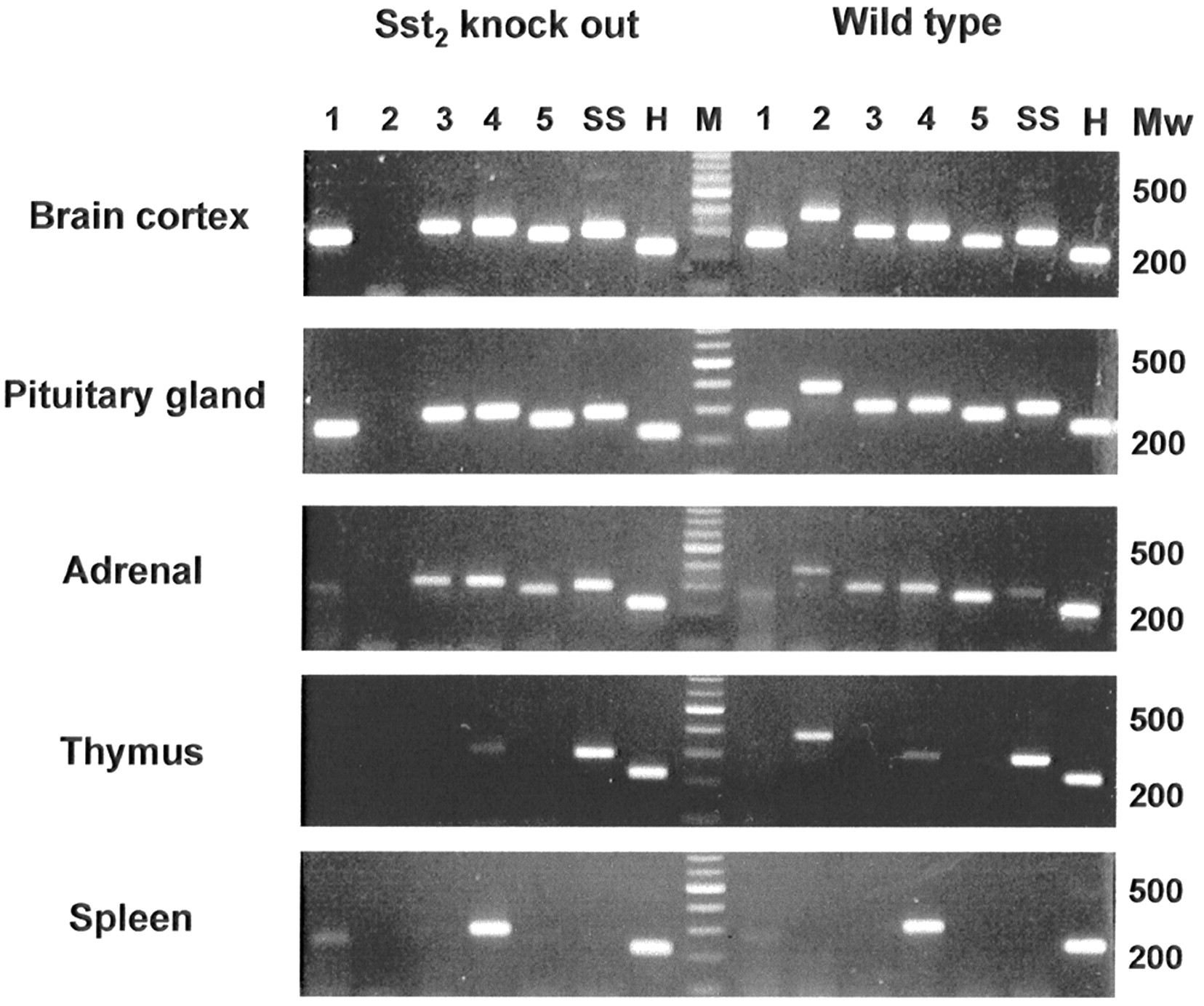
Table 2 shows the mRNA expression of sst1–5 and SS in the various organs. All 5 sst subtypes were expressed in the brain, pituitary gland, and adrenals, whereas the knockout mice lacked sst2 mRNA expression (Fig. 1; Table 2). In addition, the thymus expressed sst2 and sst4 mRNAs (Fig. 1; Table 2). Table 2 shows that sst1 and sst4 mRNAs are expressed in the spleen and kidney; sst1, sst2, sst4, and sst5 mRNAs in the liver; and sst1, sst2, and sst4 mRNAs in muscle and PBMC. Neither of these organs showed sst2 mRNA expression in the knockout mice. Compared with wild-type mice, the sst2 knockout mice expressed no compensatory sst mRNAs. SS mRNA expression was detected in the brain cortex, pituitary gland, adrenals, thymus, and muscle (soft tissue).

Expression of SS and sst1–5 mRNAs in different tissues of wild-type and sst2 knockout mice, as determined by RT-PCR. All mock-reverse-transcribed samples and all controls with no added template showed no PCR products. Marker lanes (M) contain 100-bp DNA ladder. H = hprt; Mw = molecular weight.
sst and SS mRNA Expression Pattern in Mouse Tissues as Determined by RT-PCR
Biodistribution of [111In-DTPA-d-Phe1]Octreotide
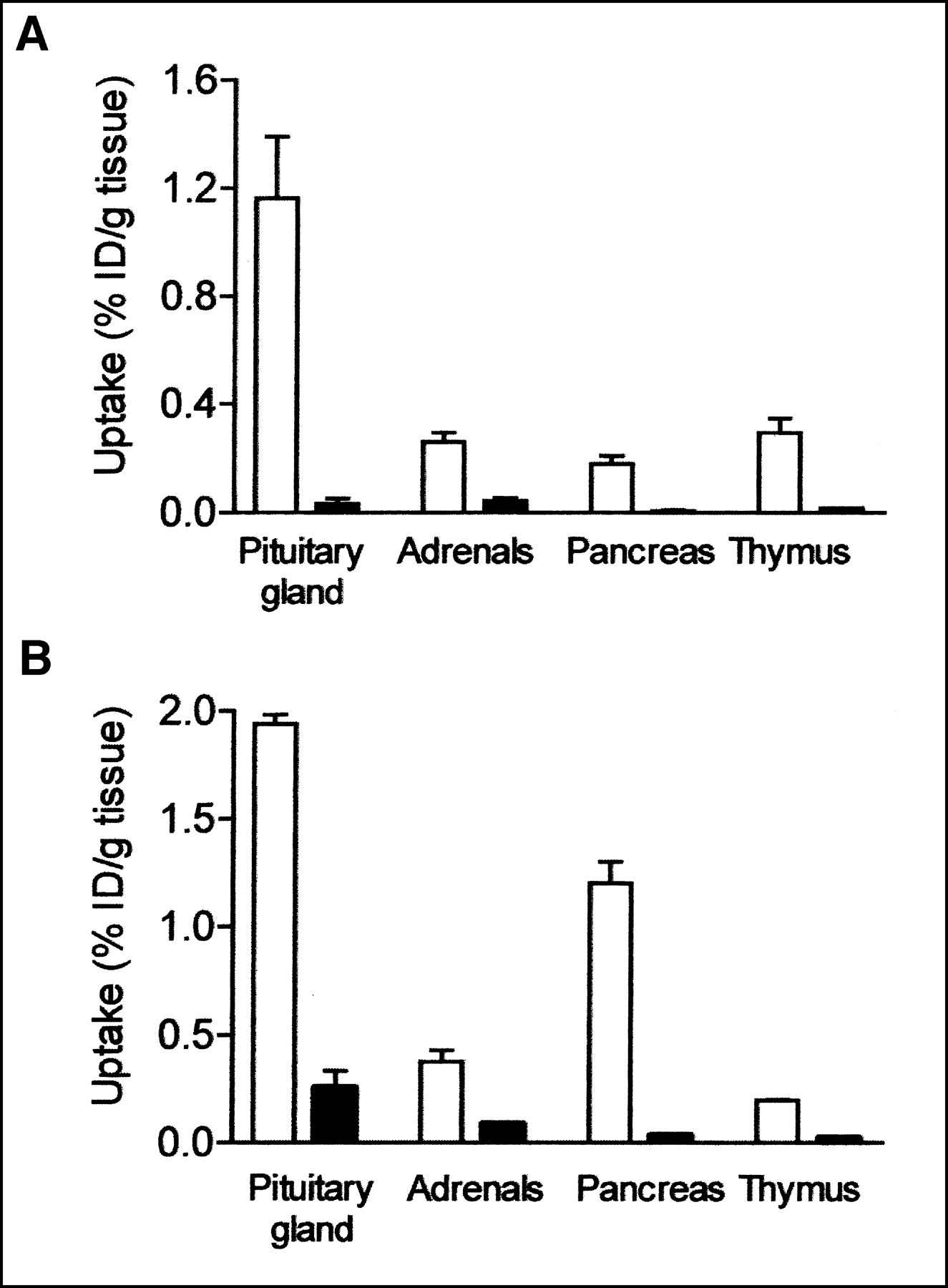
Table 3 shows the uptake values of radioactivity after the injection of [111In-DTPA-d-Phe1]octreotide (percentage injected dose per gram of tissue) in wild-type and sst2 knockout mice. Specific binding to sst as determined by a more than 50% displacement of uptake in the presence of excess unlabeled octreotide was found in the pituitary gland, adrenals, pancreas, and thymus. Interestingly, despite the presence of sst2 mRNAs, as detected by RT-PCR in the liver, soft tissue, and blood, these organs did not show specific binding of [111In-DTPA-d-Phe1]octreotide. In sst2 knockout mice, uptake values in the pituitary gland, adrenals, pancreas, and thymus were considerably reduced—by 97%, 83%, 96%, and 94%, respectively (P < 0.01 vs. wild type). Figure 2A shows the uptake of radioactivity in the pituitary gland, adrenals, pancreas, and thymus of wild-type and knockout mice. No specific binding was found in the spleen, liver, kidneys, soft tissue, or blood (Table 3). In some of these organs, uptake was even slightly higher when 100 μg of unlabeled octreotide was coinjected with the radiopharmaceutical.
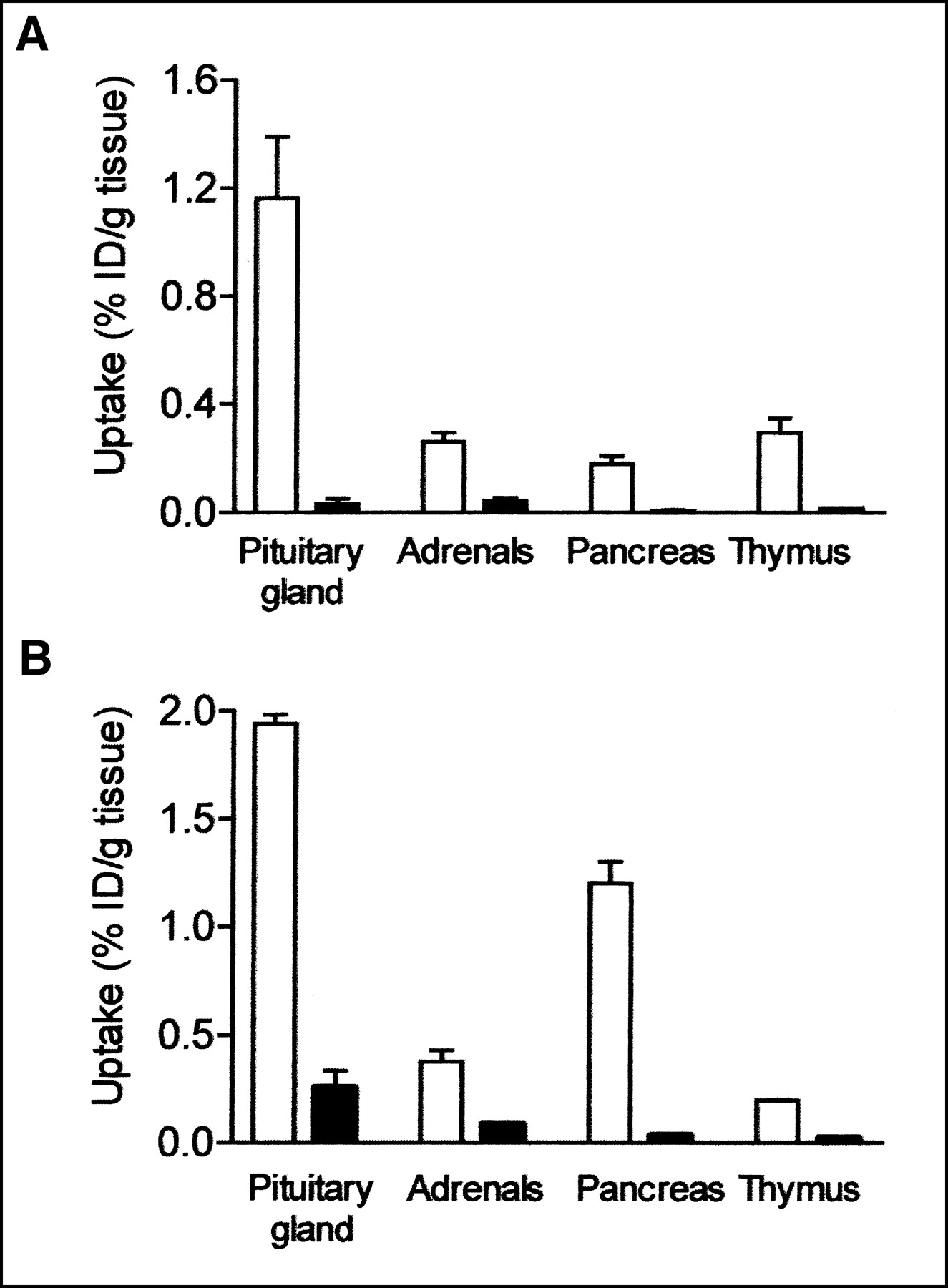
Uptake values of radioactivity after injection of [111In-DTPA-d-Phe1]octreotide (A) and [111In-DTPA]SS-14 (B), expressed as percentage injected dose per gram of tissue (% ID/gram) in pituitary gland, adrenals, pancreas, and thymus of wild-type mice (white bars) and sst2 knockout mice (black bars) (n = 4).
Tissue Distribution of [111ln-DTPA-d-Phe1]Octreotide in Wild-Type and sst2 Knockout Mice
Biodistribution of [111In-DTPA]SS-14
We also investigated tissue distribution after the injection of 1 MBq of the more universal ligand [111In-DTPA]SS-14 in wild-type and sst2 knockout mice. The [DTPA]SS-14 molecule showed high-affinity binding to sst2, sst3, and sst4—affinity comparable to that of SS-28 and SS-14. The binding affinity of [DTPA]SS-14 to sst1 and sst5 was slightly lower (Table 4). Using this radioligand, we observed a very low uptake of radioactivity in the knockout mice, comparable with the findings using [111In-DTPA-d-Phe1]octreotide. Table 5 shows the uptake values after the injection of [111In-DTPA]SS-14. Uptake values were significantly reduced in the pituitary gland (86% decrease), adrenals (76% decrease), pancreas (97% decrease), and thymus (88% decrease) of knockout mice. Figure 2B compares radioactivity uptake in the pituitary gland, adrenals, pancreas, and thymus. No specific binding of [DTPA]SS-14 was found in the spleen, liver, kidneys, soft tissue, and blood of either wild-type or sst2 knockout mice (Table 5), although these organs expressed mRNAs of several sst subtypes to which [DTPA]SS-14 binds with high affinity (Table 4). In agreement with the observations with [111In-DTPA-d-Phe1]octreotide, we found that the uptake values in organs with a very low uptake of radioactivity, as well as in sst2 knockout mice, were even slightly higher when an excess amount of 100 μg of unlabeled SS-14 was coinjected with the radiopharmaceutical.
Binding Affinity Profiles of SS-28, SS-14, and [DTPA]SS-14 for Human sst1–5 Receptors
Tissue Distribution of [111ln-DTPA]SS-14 in Wild-Type and sst2 Knockout Mice
DISCUSSION
Human sst-positive tumors show a high uptake of [111In-DTPA-d-Phe1]octreotide with sst scintigraphy (6). The majority of human neuroendocrine sst-positive tumors express multiple sst subtypes (3,5,29). The role of the individual sst in determining the uptake of radiolabeled SS-analogs is still unclear, however. Because [111In-DTPA-d-Phe1]octreotide is an sst2-preferring ligand (20), it is assumed that this sst subtype plays an important role in determining the uptake of radioactivity by sst-positive tumors via internalization of this radioligand (30). However, some studies have demonstrated significant uptake of [111In-DTPA-d-Phe1]octreotide in tumors lacking sst2. sst scintigraphy revealed tumors in patients with thyroid tumors, which lacked sst2 mRNA expression. However, analysis of sst subtype expression demonstrated the presence of the 4 other subtypes (31,32). Moreover, we recently described low but significant uptake of [111In-DTPA-d-Phe1]octreotide in a patient with a thymoma (33). In vitro studies demonstrated the absence of sst2A, sst2B, and sst5 and a predominant expression of sst3 in the thymoma tissue (33). These data suggest that sst subtypes other than sst2 could be involved in the uptake of [111In-DTPA-d-Phe1]octreotide in vivo.
In the present study, we evaluated the role of sst2 in determining the uptake of [111In-DTPA-d-Phe1]octreotide by using wild-type and sst2 knockout mice (24). In addition, we used a universal ligand, [111In-DTPA]SS-14, which binds with a relatively high affinity to the 5 sst subtypes, to investigate the role of the other sst subtypes in this respect. RT-PCR analysis of several organs of interest, that is, the pituitary gland, adrenals, thymus, spleen, liver, kidneys, soft tissue, and blood, showed that all but the spleen and kidney expressed sst2 mRNA in wild-type animals whereas the sst2 knockout animals lacked sst2 mRNA. In addition, the other 4 sst subtype mRNAs were variably expressed in these tissues. With respect to the sst1, sst3, sst4, and sst5 expression patterns, no qualitative differences between wild-type and sst2 knockout mice were observed, suggesting that no compensatory sst mRNAs are expressed after knockout of sst2 expression. However, it cannot be excluded that there are changes in sst subtype expression levels in sst2 knockout mice, compared with wild-type animals.
After the injection of [111In-DTPA-d-Phe1]octreotide, we found that uptake of radioactivity was virtually absent in sst-positive organs in sst2 knockout mice, compared with the significant uptake of radioactivity in the pituitary gland, adrenals, thymus, and pancreas in the wild-type animals. This suggests a crucial role for sst2 in determining the uptake of [111In-DTPA-d-Phe1]octreotide in normal sst-positive organs. In these organs, sst3 and sst5 apparently do not play a role in this respect, although [111In-DTPA-d-Phe1]octreotide has a relatively high binding affinity to sst5. In order to evaluate the role of the other sst subtypes in the sst-mediated uptake in vivo, we used a universal ligand, that is, [111In-DTPA]SS-14, in this mouse model as well. Surprisingly, significant uptake of radioactivity in sst-expressing organs was found only in wild-type mice, not in sst2 knockout mice. This finding indicates that sst2 is the most important, if not the crucial, sst subtype for the uptake of radiolabeled SS and SS-analogs in vivo in mice. Data on humans need to be proved, however. A possible explanation may be that the sst2, via physical interaction (e.g., heterodimerization) with one or more of the other sst subtypes, has a modulatory effect on sst-mediated internalization of these sst subtypes. Such heterodimerization of sst subtypes in vitro has previously been shown to modify the capacity of a particular sst subtype to internalize sst-bound agonists (34,35). In addition, in line with the observed crucial role of sst2, it has been shown that uptake of [111In-DTPA-d-Phe1]octreotide correlates with sst2 expression levels at the mRNA (22) as well as with sst2 protein level (21) in neuroblastoma and carcinoid tumors, respectively.
Although we did not perform quantitative measurements of sst subtype mRNA levels, wild-type and sst2 knockout mice have been shown to express comparable sst1, sst3, sst4, and sst5 mRNA levels in the brain (36). How can we explain, then, the uptake of [111In-DTPA-d-Phe1]octreotide in particular patients with tumors lacking sst2 expression? One possible explanation is a relatively high expression level of one of the other sst subtypes (e.g., sst3 or sst5), which could be involved on the basis of its binding, though at low affinity, to [111In-DTPA-d-Phe1]octreotide (20). A high expression level, in combination with the high capacity of human sst3 and sst5 to internalize after agonist binding (23), could explain the (low) tumoral uptake of radioactivity after the injection of [111In-DTPA-d-Phe1]octreotide in patients with sst2-negative tumors (32,33). Moreover, other factors, including the capacity of the cell to retain internalized radioactivity, can play a role as well.
In our present study, we observed that several organs, that is, the spleen, liver, kidneys, soft tissue, and blood, do not show specific binding in vivo of either [111In-DTPA]SS-14 or [111In-DTPA-d-Phe1]octreotide, irrespective of the presence of sst2 mRNA or mRNAs of one or more of the other sst subtypes. In some of these organs, uptake values were even slightly higher when 100 μg of unlabeled octreotide were coinjected with the radiopharmaceutical. Because this effect was also observed in the sst2 knockout mice, this slight increased uptake might be due to a systemic effect of the pharmacologic amount of unlabeled octreotide, for example, via sst5. Apparently, such a systemic effect is evident only when the uptake values are very low. The absence of specific binding in the spleen, liver, kidneys, soft tissue, and blood is unlikely to be caused by endogenous SS, competing with binding of the radioligand for sst plays a role, since these organs do not express a significant amount of SS mRNA. This indicates that, in these organs, sst protein levels are probably not high enough to result in specific binding and uptake of radioactivity in vivo. It also means that care should be taken, when drawing conclusions on the significance of the expression of a particular sst subtype, to consider its involvement in the complex processes of sst-mediated uptake of radiolabeled SS-analogs in vivo.
CONCLUSION
The present study demonstrates that the expression of sst2 is crucial for the uptake of [111In-DTPA-d-Phe1]octreotide, as well as of a radiolabeled universal SS-ligand, in normal sst-positive organs in vivo. In addition, the expression of sst2 mRNA, as determined by RT-PCR, does not necessarily result in a significant uptake of radioactivity in all organs, suggesting that sst2 protein levels in these organs are too low to result in specific binding in vivo. This may also explain why in humans the spleen, but not the thymus, is visualized through sst scintigraphy whereas both organs express sst2 mRNAs.
Footnotes
Received Feb. 11, 2002; revision accepted Mar. 28, 2003.
For correspondence or reprints contact: Leo J. Hofland, PhD, Department of Internal Medicine (Room Bd240), Erasmus Medical Center, Dr. Molewaterplein 40, 3015 GD Rotterdam, The Netherlands.
E-mail: L.hofland{at}erasmusmc.nl
REFERENCES
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The Antiproliferative Role of Lanreotide in Controlling Growth of Neuroendocrine Tumors: A Systematic Review
- Evaluating digestive neuroendocrine tumor progression and therapeutic responses in the era of targeted therapies: state of the art
- Frequency and characterization of gastro-entero-pancreatic neuroendocrine tumor patients with high-grade of uptake at somatostatin receptor scintigraphy
- Nuclear medicine techniques for the imaging and treatment of neuroendocrine tumours
- Targeting Somatostatin Receptors: Preclinical Evaluation of Novel 18F-Fluoroethyltriazole-Tyr3-Octreotate Analogs for PET
- Indium-111-Pentetreotide Scintigraphy and Somatostatin Receptor Subtype 2 Expression: New Prognostic Factors for Malignant Well-Differentiated Endocrine Tumors
- Novel insights in somatostatin receptor physiology
- The analysis of quantitative expression of somatostatin and dopamine receptors in gastro-entero-pancreatic tumours opens new therapeutic strategies
- In Vitro and in Vivo Antitumor Effects of Cytotoxic Camptothecin-Bombesin Conjugates Are Mediated by Specific Interaction with Cellular Bombesin Receptors
- Increased Uptake of 111In-Octreotide in Idiopathic Pulmonary Fibrosis
- MicroPET Imaging of Gene Transfer with a Somatostatin Receptor-Based Reporter Gene and 94mTc-Demotate 1
- The multi-ligand somatostatin analogue SOM230 inhibits ACTH secretion by cultured human corticotroph adenomas via somatostatin receptor type 5
- Detection of Liver Metastases From Endocrine Tumors: A Prospective Comparison of Somatostatin Receptor Scintigraphy, Computed Tomography, and Magnetic Resonance Imaging
- Induction of Apoptosis with Hybrids of Arg-Gly-Asp Molecules and Peptides and Antimitotic Effects of Hybrids of Cytostatic Drugs and Peptides