


Abstract
For solid tumors and metastatic lesions, tumor vascularity is a critical factor in assessing response to therapy. Here we report the first example, to our knowledge, of 64Cu-labeled vascular endothelial growth factor 121 (VEGF121) for PET of VEGF receptor (VEGFR) expression in vivo. Methods: VEGF121 was conjugated with 1,4,7,10-tetraazadodecane-N,N′,N′′,N′′′-tetraacetic acid (DOTA) and then labeled with 64Cu for small-animal PET of mice bearing different sized U87MG human glioblastoma xenografts. Blocking experiments and ex vivo histopathology were performed to confirm the in vivo results. Results: There were 4.3 ± 0.2 DOTA molecules per VEGF121, and the VEGFR2 binding affinity of DOTA-VEGF121 was comparable to VEGF121. 64Cu labeling of DOTA-VEGF121 was achieved in 90 ± 10 min and the radiolabeling yield was 87.4% ± 3.2%. The specific activity of 64Cu-DOTA-VEGF121 was 3.2 ± 0.1 GBq/mg with a radiochemical purity of >98%. Small-animal PET revealed rapid, specific, and prominent uptake of 64Cu-DOTA-VEGF121 in small U87MG tumors (high VEGFR2 expression) but significantly lower and sporadic uptake in large U87MG tumors (low VEGFR2 expression). No appreciable renal clearance of 64Cu-DOTA-VEGF121 was observed, although the kidney uptake was relatively high likely due to VEGFR1 expression. Blocking experiments, immunofluorescence staining, and western blot confirmed the VEGFR specificity of 64Cu-DOTA-VEGF121. Conclusion: Successful demonstration of the ability of 64Cu-DOTA-VEGF121 to visualize VEGFR expression in vivo may allow for clinical translation of this radiopharmaceutical for imaging tumor angiogenesis and guiding antiangiogenic treatment, especially patient selection and treatment monitoring of VEGFR-targeted cancer therapy.
- vascular endothelial growth factor
- Flk-1/KDR (vascular endothelial growth factor receptor 2)
- PET
- tumor vasculature
- 64Cu
Vascular endothelial growth factor A (VEGF-A) plays a central role in both normal vascular tissue development and tumor neovascularization (1). VEGF-A is an endothelial cell-specific angiogenic protein expressed in various human tumors (2). Through alternative splicing of RNA, VEGF may exist as at least 7 different molecular isoforms, having 121, 145, 148, 165, 183, 189, or 206 amino acids (3). These isoforms differ not only in their molecular weight but also in their biologic properties, such as the ability to bind to cell-surface heparin sulfate proteoglycans. VEGF121 is a soluble, nonheparin-binding variant that exists in solution as a disulfide-linked homodimer containing the full biologic and receptor-binding activity of the larger variants (1,4).
The angiogenic actions of VEGF are mediated primarily via two closely related endothelium-specific receptor tyrosine kinases, Flt-1 (VEGF receptor 1 [VEGFR1]) and Flk-1/KDR (VEGFR2) (5). Both are largely restricted to vascular endothelial cells and are overexpressed on the endothelium of tumor vasculature, yet they are almost undetectable in the vascular endothelium of adjacent normal tissues (2). All of the VEGF-A isoforms bind to both VEGFR1 and VEGFR2. VEGF and its receptors are overexpressed in a variety of solid tumor biopsy specimens, and overexpression of VEGFR2 or VEGF-A has been implicated as a poor prognostic marker in various clinical studies (6–8). Agents that prevent VEGF-A binding to its receptors (9), antibodies that directly block VEGFR2 (10), and small molecules that inhibit the kinase activity of VEGFR2 (11,12), and thereby block growth factor signaling, are all under active development. The contribution of VEGF-A to cancer progression has been highlighted by the recent approval of the humanized anti-VEGF monoclonal antibody bevacizumab (Avastin; Genentech) for first-line treatment (13). Recently, VEGFR2-targeted delivery of toxins also showed impressive antitumor vascular-ablative effects (14,15).
Recombinant human VEGF121 was labeled with 111In for identification of ischemic tissue in a rabbit model of unilateral hindlimb ischemia created by femoral artery excision. Only marginal difference was observed between the ischemic hindlimb and the contralateral hindlimb (16). 123I-VEGF165 was also reported as a potential tumor marker (17). Despite the high receptor affinity of this radiopharmaceutical, the biodistribution in A2508 melanoma tumor-bearing mice indicated low tumor-to-background ratio, most likely due to the low metabolic stability of the compound (18). VEGF121 has been labeled with 99mTc through an “adapter/docking” strategy for imaging 4T1 murine mammary carcinoma tumors. Low tumor signal (∼2–4 percentage injected dose per gram [%ID/g]) and poor tumor-to-background contrast were obtained (19). A recombinant protein composed of VEGF165 fused through a flexible polypeptide linker (GGGGS)3 to the n-lobe of human transferrin (hnTf) has also been reported for imaging tumor angiogenesis, and the tumor uptake was modest (20). In all of these reports, radiolabeled VEGF molecules were used for SPECT with limited success. To date, no radiolabeled VEGF has been developed for PET for any type of disease, although a few radiolabeled anti-VEGF antibodies have been reported (21,22). PET has several advantages over SPECT, including 10-fold greater sensitivity, and the increasing implementation of clinical PET and PET/CT scanners can facilitate the translation of novel PET radiopharmaceuticals to the clinic (23).
Successful development of VEGF-based PET could serve as a paradigm for assessment of cancer therapeutics targeting tumor angiogenesis. The ability to noninvasively visualize and quantify tumor VEGFR expression levels could provide new opportunities to document tumor angiogenesis status, more appropriately select patients considered for antiangiogenic treatment, and monitor antiangiogenic treatment efficacy more effectively. Herein we report, to our knowledge, the first example of 64Cu (half-life, 12.7 h; 39% β−; 17.4% β+)-labeled VEGF121 for PET of tumor angiogenesis and VEGFR expression.
MATERIALS AND METHODS
All commercially available chemical reagents were used without further purification. 1,4,7,10-Tetraazadodecane-N,N′,N″,N″′-tetraacetic acid (DOTA) was purchased from Macrocyclics, Inc. 1-Ethyl-3-[3-(dimethylamino)propyl]carbodiimide, N-hydroxysulfonosuccinimide, and Chelex 100 resin (50–100 mesh) were purchased from Aldrich. Water and all buffers were passed through a Chelex 100 column before use in radiolabeling procedures to ensure heavy metal–free conditions. PD-10 columns were purchased from GE Healthcare. Female athymic nude mice were supplied by Harlan at 4–5 wk of age. 64Cu was obtained from the University of Wisconsin, Madison.
VEGF121 Preparation
The gene for VEGF121 was cloned by polymerase chain reaction from human umbilical vein endothelial cells, sequenced, and inserted into the pET-32 vector (Novagen) downstream from the cleavable His6 tag sequence. Bacterial host cells were transformed and selected under antibiotic resistance, and positive clones were selected for optimal protein expression. Cells were lysed by sonication. The lysate was ultracentrifuged at 40,000 rpm for 90 min at 4°C. The supernatant was carefully collected, adjusted to 40 mmol/L Tris-HCl (pH 8.0) and 300 mmol/L NaCl, and run through an immobilized metal affinity column (cobalt). The resin was washed with 40 mmol/L Tris-HCl (pH 8.0), 0.3 mol/L NaCl, and 20 mmol/L imidazole buffer and eluted with buffer containing 300 mmol/L imidazole. After pooling fractions containing the His-tagged VEGF121, the sample was dialyzed against 20 mmol/L Tris-HCl (pH 8.0) and 100 mmol/L NaCl and digested with recombinant enterokinase at room temperature to cleave the His6 tag from VEGF121. The enterokinase was removed by agarose-linked soybean trypsin inhibitor. The sample was then dialyzed against 20 mmol/L Tris-HCl (pH 8.0) and VEGF121 was purified by NaCl gradient elution after binding to Q Sepharose Fast Flow resin (GE Healthcare). VEGF121 was concentrated using Centricon 10 concentrators (Amicon, Inc.) and stored in sterile phosphate-buffered saline (PBS) at −20°C.
DOTA Conjugation and Radiolabeling
The detailed procedure for DOTA conjugation has been reported earlier (24,25). DOTA-VEGF121 was purified using a PD-10 column and concentrated by Centricon filter (Ultracel YM-10; Millipore). The final concentration of DOTA-VEGF121 was determined on the basis of ultraviolet absorbance at 280 nm using unconjugated VEGF121 of known concentrations as the standard. 64CuCl2 (74 MBq) was diluted in 300 μL of 0.1 mol/L sodium acetate buffer (NaOAc, pH 6.5) and added to 20 μg of DOTA-VEGF121. The reaction mixture was incubated for 1 h at 40°C with constant shaking. 64Cu-DOTA-VEGF121 was purified using a PD-10 column with PBS as the mobile phase. The radioactive fractions containing 64Cu-DOTA-VEGF121 were collected and passed through a 0.2-μm low-protein-binding syringe filter (Nalge Nunc International) for further in vitro and in vivo experiments. 64Cu-DOTA-VEGF121 was also incubated with complete mouse serum at 37°C for up to 24 h to evaluate the stability of this radiopharmaceutical.
The average number of DOTA chelators per VEGF121 was determined using a previously reported procedure with slight modifications (26). Briefly, 5 μg of DOTA-VEGF121 in 100 μL 0.1N NaOAc buffer were added to a defined amount of carrier-added 64CuCl2 solution. The number of DOTA molecules per VEGF121 was calculated using the following equation: The results are expressed as mean ± SD (n = 3).
The results are expressed as mean ± SD (n = 3).
Cell Lines and Animal Model
The U87MG human glioblastoma cell line was obtained from the American Type Culture Collection and cultured under standard conditions (27,28). Porcine aortic endothelial cells that express human KDR (PAE/KDR) were cultured in Ham's F-12 medium containing 10% fetal calf serum (Sigma). Animal procedures were performed according to a protocol approved by the Stanford University Institutional Animal Care and Use Committee. The U87MG tumor model was generated by subcutaneous injection of 5 × 106 cells in 50 μL PBS into the front leg of the mice. The mice were subjected to microPET studies when the tumor volume reached about 60 mm3 (small tumor, 1–2 wk after inoculation) or 1,200 mm3 (large tumor, 4 wk after inoculation).
Cell-Binding Assay and Functional Assay
The detailed procedure for the cell-binding assay has been reported earlier (29,30). Receptor-binding affinity of VEGF121 and DOTA-VEGF121 was analyzed by PAE/KDR cell-binding assay using 125I-VEGF165 as the radioligand. To determine the functional activity of DOTA-VEGF121, PAE/KDR cells were stimulated by serial concentrations of VEGF121 or DOTA-VEGF121 for 5 min, and the cell lysates were immunoblotted by antiphophorylated VEGFR2 antibody (Abcam Inc.).
MicroPET Studies
PET of tumor-bearing mice was performed on a microPET R4 rodent model scanner (Siemens Medical Solutions USA, Inc.) as described earlier (31,32). Each mouse was injected with about 5–10 MBq of 64Cu-DOTA-VEGF121 (∼2–4 μg of VEGF121) via tail vein. For each microPET scan, 3-dimensional region of interests (ROIs) were drawn over the tumor, liver, kidneys, and muscle on decay-corrected whole-body coronal images. The average radioactivity concentration within a tumor or an organ was obtained from mean pixel values within the ROI volume, which were converted to counts per milliliter per minute by using a conversion factor. Assuming a tissue density of 1 g/mL, the counts per milliliter per minute were converted to counts per gram per minute and then divided by the injected dose (ID) to obtain an imaging ROI-derived %ID/g. Mice bearing small U87MG tumors were also imaged using 64Cu-DOTA-VEGF121 injected 30 min after injection of 100 μg of VEGF121 (n = 3). Nine mice were used for this study: 3 small tumor-bearing mice, 3 large tumor-bearing mice, and 3 mice for blocking and biodistribution studies.
Radiation Dosimetry Extrapolation to Humans
Estimated human dosimetry was calculated from microPET results on Sprague–Dawley female rats (Harlan) injected with about 37 MBq of 64Cu-DOTA-VEGF121 (n = 3), assuming that the biodistribution of the radiopharmaceutical in rats is the same as in adult humans. The rats were scanned at 2 bed positions to cover the whole body, and ROI analysis was performed on major organs. Time–activity curves were generated from the mean values obtained in rats for each organ of interest. We then calculated source organ residence times for the human model by integrating a monoexponential fit to the experimental biodistribution data for major organs (blood, liver, kidneys, and muscle) and the whole body. The source organ residence times obtained forthwith were used with a standard quantitation platform, Organ Level Internal Dose Assessment (OLINDA; Vanderbilt University) (33).
Western Blot
U87MG tumor tissue protein was extracted using T-PER tissue protein extraction buffer (Pierce Biotechnology, Inc.) and the concentration was determined using a MicroBCA Protein Assay kit (Pierce Biotechnology, Inc.). After sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) separation of 100 μg of total protein, it was transferred to a polyvinylidene fluoride membrane (Invitrogen Corp.) and incubated at room temperature with 5% nonfat milk blocking buffer. The blots were then incubated overnight at 4°C with rabbit anti-VEGFR2 primary antibody (Lab Vision Corp.) followed by incubation at room temperature for 1 h with horseradish peroxidase–conjugated antirabbit secondary antibody (GE Healthcare). Tubulin was used as the loading control, and the bands were detected using the electrochemoluminescent Western Blotting Detection system (GE Healthcare).
Immunofluorescence Staining
U87MG tumor and kidney frozen tissue slices (5-μm thickness) were fixed with cold acetone for 10 min and dried in air for 30 min. The slices were rinsed with PBS for 2 min and blocked with 10% donkey serum for 30 min at room temperature. The slices were then incubated with rat antimouse VEGFR2 antibody overnight at 4°C and visualized using Cy3-conjugated donkey antirat secondary antibody (1:200; Jackson ImmunoResearch Laboratories, Inc.). For VEGFR1 staining, the tissue slices were incubated with rabbit antimouse VEGFR1 antibody (1:50; Lab Vision Corp.) at room temperature for 1 h and visualized with Cy3-conjugated donkey antirabbit secondary antibody (1:200; Jackson ImmunoResearch Laboratories, Inc.). For CD31 staining, the slices were incubated with rat antimouse CD31 antibody (1:100; BD Biosciences) at room temperature for 1 h and visualized with Cy3-conjugated donkey antirat secondary antibody (1:200; Jackson ImmunoResearch Laboratories, Inc.).
After CD31 staining, 7 random views in both the center and the periphery of the tumor slices were selected for microvessel density (MVD) analysis using an observer-set threshold to distinguish vascular elements from surrounding tissue parenchyma. The vessel that contains branching points was counted as a single vessel. The number of vessels counted was divided by the field of view to yield the MVD, as vessels/mm2.
Statistical Analysis
Quantitative data were expressed as mean ± SD. Means were compared using 1-way ANOVA and the Student t test. P values < 0.05 were considered statistically significant.
RESULTS
Receptor-Binding Assay and Functional Activity of DOTA-VEGF121
The binding of VEGF121 and DOTA-VEGF121 to endothelial cells expressing VEGFR2 was assessed using 125I-VEGF165 as the radioligand. The 50% inhibitory concentration (IC50) values were 1.02 nmol/L and 1.66 nmol/L for VEGF121 and DOTA-VEGF121, respectively (Fig. 1A). The minimal difference in VEGFR2 binding affinity between VEGF121 and DOTA-VEGF121 suggested that the lysine residues or N-terminal amine groups used for DOTA conjugation may not be located at the VEGFR2 binding domain. DOTA conjugation of VEGF121 resulted in decreased functional activity (Fig. 1B). However, full functional activity is not required for imaging applications.

Cell-binding assay and functional assay. (A) Cell-binding assay of VEGF121 and DOTA-VEGF121 using PAE/KDR cells. IC50 values are 1.02 and 1.66 nmol/L for VEGF121 and DOTA-VEGF121, respectively. (B) Functional assay of VEGF121 and DOTA-VEGF121. Tubulin was used as loading control.
Radiolabeling of DOTA-VEGF121
64Cu labeling, including the final purification, of DOTA-VEGF121 took 90 ± 10 min (n = 5), and the radiolabeling yield was 87.4% ± 3.2% (based on 37 MBq of 64Cu per 10 μg of DOTA-VEGF121; n = 5). The specific activity of 64Cu-DOTA-VEGF121 was 3.2 ± 0.1 GBq/mg, with a radiochemical purity of >98%. The number of DOTA molecules per VEGF121 molecule was determined to be 4.3 ± 0.2 (n = 3). No significant metabolite peak was observed on radio–high-performance liquid chromatography when 64Cu-DOTA-VEGF121 was incubated with mouse serum at 37°C for up to 24 h, indicating that the radiopharmaceutical is stable in mouse serum.
MicroPET of 64Cu-DOTA-VEGF121 in U87MG Tumor-Bearing Mice
Mice bearing small (tumor volume: 64.9 ± 24.6 mm3, n = 3; high VEGFR expression) and large (tumor volume: 1,164.3 ± 179.6 mm3, n = 3; low VEGFR expression) U87MG tumors were subjected to microPET scans at various time points after injection of 64Cu-DOTA-VEGF121. The coronal slices that contain the tumor are shown in Figure 2A. The uptake of 64Cu-DOTA-VEGF121 into small tumors was rapid and high, reaching 14.9 ± 0.7, 16.3 ± 0.7, 16.3 ± 0.6, and 15.1 ± 0.8 %ID/g at 2, 4, 16, and 23 h after injection, respectively. As early as 1 h after injection, the tumor was clearly visible (data not shown). The accumulation of 64Cu-DOTA-VEGF121 in the large tumor was at a very low level at all time points examined, with the uptake at the peripheral region (∼3–4 %ID/g) slightly higher than at the necrotic center of the tumor (∼1–2 %ID/g). The 64Cu-DOTA-VEGF121 uptake was significantly different between the small tumors and the large tumors at all time points examined (P < 0.01, Fig. 3A).

MicroPET of 64Cu-DOTA-VEGF121 in U87MG tumor-bearing mice. (A) Serial microPET scans of large and small U87MG tumor-bearing mice injected intravenously with 5–10 MBq of 64Cu-DOTA-VEGF121 (∼2–4 μg of VEGF121). Mice injected with 64Cu-DOTA-VEGF121 30 min after injection of 100 μg VEGF121 are also shown (denoted as “Small tumor + block”). (B) Two-dimensional whole-body projection of the 3 mice shown in A at 16 h after injection of 64Cu-DOTA-VEGF121. Tumors are indicated by arrows.
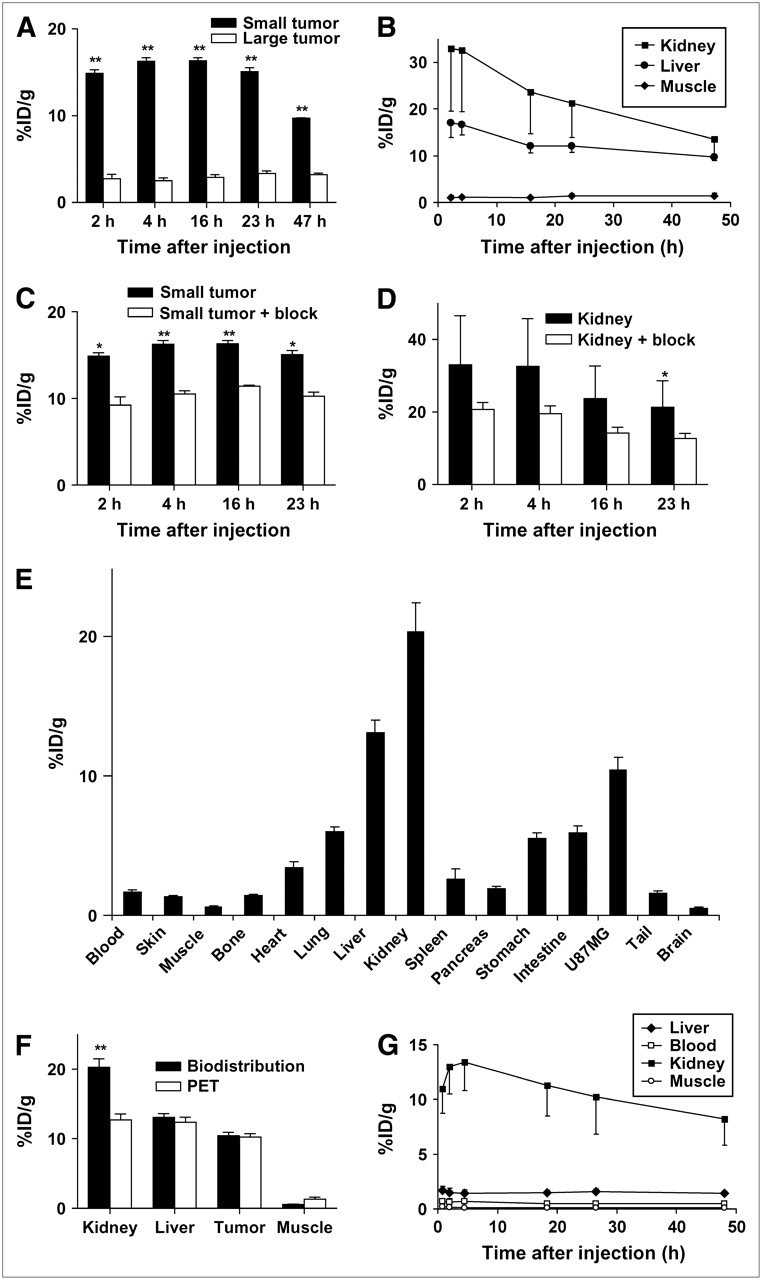
MicroPET and biodistribution results. (A) Comparison of 64Cu-DOTA-VEGF121 uptake in small and large U87MG tumors (3 mice per group). (B) Time–activity curves of 64Cu-DOTA-VEGF121 uptake in the kidney, liver, and muscle (n = 6). (C) Comparison of 64Cu-DOTA-VEGF121 uptake in small U87MG tumors with those injected previously with 100 μg of VEGF121 (3 mice per group). (D) Comparison of 64Cu-DOTA-VEGF121 uptake in kidneys (n = 6) with those injected previously with 100 μg of VEGF121 (n = 3). (E) Biodistribution at 23 h after injection of 64Cu-DOTA-VEGF121 in mice injected previously with 100 μg of VEGF121 (n = 3). (F) Comparison of quantification results obtained from biodistribution and microPET studies (n = 3). (G) Time–activity curves of liver, blood, kidney, and muscle in Sprague–Dawley rats injected with 64Cu-DOTA-VEGF121 (n = 3). *P < 0.05; **P < 0.01.
64Cu-DOTA-VEGF121 exhibited high uptake in the kidneys and liver at early time points (33.0 ± 13.5 and 17.1 ± 3.2 %ID/g at 2 h after injection, respectively; n = 6, 3 small tumor-bearing mice and 3 large tumor-bearing mice). The uptake in most other organs was at a very low level. The uptake of 64Cu-DOTA-VEGF121 in both the kidneys and the liver dropped steadily over time (Fig. 3B). For the kidneys, the uptake was 33.0 ± 13.5, 21.3 ± 7.3, and 13.5 ± 3.5 %ID/g at 2, 23, and 47 h after injection, respectively. Although the kidney uptake was high, there was no appreciable activity accumulation in the urinary bladder, suggesting a very slow renal clearance rate of 64Cu-DOTA-VEGF121.
Blocking and Biodistribution Studies
To test the VEGFR specificity of 64Cu-DOTA-VEGF121 in vivo, blocking experiments were performed, in which 100 μg VEGF121 were injected into small U87MG tumor-bearing mice (tumor volume, 61.3 ± 10.6 mm3, n = 3) 30 min before administration of 64Cu-DOTA-VEGF121. As can be seen from both the coronal slices (Fig. 2A) and the 2-dimensional whole-body projection at 16 h after injection (Fig. 2B), the small U87MG tumor uptake is clearly lower compared with that of those mice without VEGF121 blocking. Although 100 μg of VEGF121 (∼4 mg/kg) was unable to completely block the tumor uptake of 64Cu-DOTA-VEGF121, the activity accumulation in the tumor (9.2 ± 1.7, 10.5 ± 0.6, 11.4 ± 0.2, and 10.3 ± 0.8 %ID/g at 2, 4, 16, and 23 h after injection, respectively) was significantly lower at all time points examined compared with that of the control mice (P < 0.05; Fig. 3C). The kidney uptake of 64Cu-DOTA-VEGF121 was also lower in the mice injected with VEGF121. However, statistical significance was achieved only at 23 h after injection (Fig. 3D), due to the large variance in kidney uptake between individual animals and the insufficient blocking dose of VEGF121. The variance in kidney uptake of 64Cu-DOTA-VEGF121 between each individual animal was not fully understood. Successful partial blocking of the small U87MG tumor and the kidney uptake with a limited dose of VEGF121 demonstrated the VEGFR specificity of 64Cu-DOTA-VEGF121 in vivo.
After the microPET scans at 23 h after injection, the mice were immediately sacrificed and biodistribution studies were performed (Fig. 3E). When comparing the quantification results obtained from biodistribution studies and PET scans, there was no significant difference between the liver, tumor, and muscle (P > 0.05; Fig. 3F), suggesting that quantification of noninvasive microPET scans is a true reflection of the biodistribution of 64Cu-DOTA-VEGF121 in these organs. The difference in kidney uptake between the 2 studies was likely due to the heterogeneity of 64Cu-DOTA-VEGF121 uptake in the kidney and the difficulty in ROI analysis because of its irregular shape.
Immunofluorescence Staining and Western Blot
After decay of most of the radioactivity in the microPET studies, the mice were sacrificed and the frozen tumor slices were stained for CD31, VEGFR1, and VEGFR2. As can be seen in Figure 4, VEGFR1 expression was low in both small and large U87MG tumors. VEGFR1 expression is known to be expressed on endothelial cells of preglomerular vessels, glomeruli, and postglomerular vessels in the kidneys, which is most likely responsible for the observed high kidney uptake of 64Cu-DOTA-VEGF121. Small U87MG tumors had high VEGFR2 expression, whereas large tumors demonstrated very low VEGFR2 expression, resulting in high 64Cu-DOTA-VEGF121 uptake in small U87MG tumors but very low uptake in large tumors. There was also a measurable level of VEGFR2 in the kidneys, primarily on the small vessels but not on the large, mature vessels. CD31 staining indicated much higher vascular density in the small U87MG tumor than in the large tumor. The vessels in the small tumor were mostly of regular shape, whereas the vessels in the large U87MG tumor had much larger diameter and the shapes were more irregular. Costaining of VEGFR2 and CD31 was unsuccessful. However, visual examination of CD31 and VEGFR2 staining of different slices in the same tumor suggests that VEGFR2 and CD31 are colocalized on the newly developed tumor vessels. MVD analysis revealed that the small U87MG tumor has a significantly higher vessel density (96 ± 19 vessels/mm2) than the large tumor (20 ± 9 vessels/mm2; P < 0.01, Fig. 5A). Western blot also showed a higher VEGFR2 protein level in the small U87MG tumor than in the large tumor (Fig. 5B). Because of the low expression level of VEGFR1 in both the small and the large U87MG tumors, a western blot of VEGFR1 was not obtained. The good correlation of ex vivo results with in vivo microPET indicates that noninvasive microPET using 64Cu-DOTA-VEGF121 can reflect the VEGFR expression level in vivo.

Immunofluorescence staining of VEGFR1, VEGFR2, and CD31 for kidney, small U87MG tumor, and large U87MG tumor. For VEGFR1 staining, frozen tissue slices (5-μm thick) were stained with a rabbit antimouse VEGFR1 primary antibody and a Cy3-conjugated donkey antirabbit secondary antibody. For VEGFR2 staining, tissue slices were stained with a rat antimouse VEGFR2 primary antibody and a Cy3-conjugated donkey antirat secondary antibody. For CD31 staining, slices were stained with a rat antimouse CD31 primary antibody and a Cy3-conjugated donkey antirat secondary antibody.

MVD analysis and western blot. (A) MVD analysis of small and large U87MG tumor. **P < 0.01. (B) Western blot of VEGFR2 in small and large U87MG tumor. Tubulin was used as loading control.
Radiation Dosimetry
Human absorbed doses to normal organs from 64Cu-DOTA-VEGF121 were estimated from microPET quantification data in female Sprague–Dawley rats, assuming that the biodistribution and pharmacokinetics of 64Cu-DOTA-VEGF121 in rats and adult human are the same, and the results are presented in Table 1. The 64Cu-DOTA-VEGF121 uptake of different organs in rats was similar to that of mice (Fig. 3G). Except for the blood, liver, and kidneys, all other organs exhibited very low level of 64Cu-DOTA-VEGF121 uptake at all time points examined. The radiopharmaceutical was excreted primarily via the hepatic pathway. No appreciable activity was observed in the urinary bladder at all time points examined, suggesting that 64Cu-DOTA-VEGF121 also has a very slow renal clearance in rats. The highest radiation-absorbed dose is to the kidneys (1.05 ± 0.27 mGy/MBq). Except for the kidneys and the liver (0.12 ± 0.02 mGy/MBq), all other organs have a very low level of radiation-absorbed doses. The whole-body absorbed dose was found to be 0.05 ± 0.01 mGy/MBq administered. Although the dose-limiting organ is the kidneys, the relatively low radiation dose will not likely cause any adverse effect, as 64Cu-DOTA-VEGF121 will be used only for imaging application, where limited radioactivity (∼200–400 MBq) will be injected in patients.
Estimated Radiation-Absorbed Doses to Adult Human After Intravenous Injection of 64Cu-DOTA-VEGF121 Based on microPET Imaging Data Obtained in Female Sprague–Dawley Rats (n = 3)
DISCUSSION
This study demonstrated that 64Cu-labeled VEGF121 exhibits strong and specific VEGFR-binding affinity both in vitro and in vivo. Food and Drug Administration approval of anti-VEGF antibody bevacizumab and the fact that many other antibody and small molecule inhibitors against VEGFR2 are currently in advanced clinical trials confirm the validity and importance of VEGF/VEGFR2 signaling in anticancer therapy (10–12). PET using radiolabeled VEGF121 is critical in early and sensitive lesion detection, in patient selection for clinical trials based on in vivo VEGFR expression quantification, in better treatment monitoring and dose optimization based on noninvasive detection of early response to VEGF- or VEGFR-targeted therapy, and in elucidating the mechanisms of treatment efficacy underlying VEGF/VEGFR signaling.
It is well accepted that tumor angiogenesis occurs when the tumor reaches a certain size (usually 1–2 mm in diameter), as new blood vessel formation is needed to supply oxygen and nutrients to cancer cells and to remove waste (34). VEGF-A and its receptors are the best-characterized signaling pathway in developmental angiogenesis as well as tumor angiogenesis (2). VEGFR2 appears to be the most important receptor in VEGF-induced mitogenesis, angiogenesis, and permeability increase, whereas the role of VEGFR1 in endothelial cell function is less clear (1). During the exponential growth stage, VEGFR expression is highly upregulated on the newly developed tumor vasculature. Being the naturally existing VEGFR ligand, VEGF121 offers several advantages over the synthetic small-molecule VEGFR ligands or anti-VEGFR antibodies. It has much higher binding affinity to VEGFR (nanomolar range) than reported peptidic VEGFR inhibitors (submicromolar to micromolar range) (35,36). Comparing with antibody-based radiopharmaceuticals, VEGF121 clears much faster from the blood pool and the nontargeting organs because of its smaller size (25 kDa for the dimeric form of VEGF121). In this proof-of-principle study, we chose two different sizes of the same tumor type, hypothesizing that the small tumors have higher VEGFR expression than the large tumors.
Indeed, in the small U87MG tumor, where the diameter is about 4–6 mm, the tumor is at an exponential growth stage. Both immunofluorescence staining (VEGFR1, VEGFR2, and CD31; Fig. 4) and western blot (Fig. 5B) demonstrated that there is a high level of VEGFR2 expression in the tumor. For the large tumor, where the tumor diameter is about 10–15 mm, the tumor vessels are primarily mature (much larger diameter compared with the vessels in the small tumor, and the vessel density is also much lower), and both VEGFR1 and VEGFR2 expression are quite low, as confirmed by ex vivo immunofluorescence staining and western blot. The ex vivo results supported the in vivo PET findings, where the small tumors have much higher 64Cu-DOTA-VEGF121 uptake (∼15 %ID/g) than that in the large tumors (∼2–3 %ID/g). PET of other less vascularized tumors (e.g., MDA-MB-435 breast cancer, where the vessel density is quite low when the tumor reaches a certain size) also exhibited very low 64Cu-DOTA-VEGF121 uptake (data not shown). Tiny (<2 mm in diameter) and medium-size (8–10 mm in diameter) U87MG tumors had a very low (<3 %ID/g) and an intermediate level (5–8 %ID/g) of 64Cu-DOTA-VEGF121 uptake, respectively (data not shown). These results suggest that the time window of high VEGFR expression is quite narrow. In the clinical setting, the right timing is critical for VEGFR-targeted cancer therapy. PET using 64Cu-DOTA-VEGF121 can play a very important role in determining whether, and when, to start the VEGFR-targeted cancer therapy, as it can provide a straightforward and convenient way to monitor VEGFR expression level in vivo.
In addition to immunofluorescence staining and western blot analysis, FACS sorting of tumor cells may be another method to quantify VEGFR2 expression. Tumor cells can be harvested and stained for CD31 or VEGFR1/VEGFR2. Florescence-activated cell sorter analysis can then be performed to evaluate the VEGFR1/VEGFR2 expression level on endothelial cells. Other microvessel markers such as CD105 may also be used to correlate VEGFR expression and MVD in future studies (37). VEGFR2-specific ligand can be developed in the future. It may offer a certain advantage over VEGF121-based radiopharmaceuticals, such as lower renal uptake (VEGF121 binds both VEGFR1 and VEGFR2 and kidney has high VEGFR1 expression). A certain level of VEGFR2 expression in the kidney was also observed in this study (Fig. 4). Comparison of the CD31 and VEGFR2 staining of the kidney slices showed that the VEGFR2 expression is localized primarily to the glomerulus but not on the big vessels (e.g., intralobular vein or the efferent arteriole), which is expected as VEGFR2 is expressed primarily on microvessels. Peptidic VEGFR antagonists that can be labeled with 18F (more readily available than 64Cu) may also be tested, and it can allow for high throughput, as usually 1–2 h after injection is sufficient for a peptide-based radiopharmaceutical to clear from the nontargeted organs and give high-contrast PET images.
Endogenous VEGF isoforms may also compete with 64Cu-DOTA-VEGF121 in VEGFR binding. However, the intact VEGFR-binding potency of DOTA-VEGF121 and the fact that the endogenous VEGF concentration is far from saturating the VEGFR resulted in prominent 64Cu-DOTA-VEGF121 uptake in the high VEGFR-expressing small U87MG tumors. Although the functional activity of DOTA-VEGF121 is significantly lower than that of VEGF121, this is not a concern for imaging applications. Serial microPET of 64Cu-DOTA-VEGF121 indicated that 64Cu is the optimal isotope for VEGF121-based radiopharmaceuticals, as the half-life of 64Cu (12.7 h) is well suited for the time frame needed to follow the radiopharmaceutical uptake and clearance (48 h in this study, slightly less than 4 half-lives of 64Cu). Although not evaluated, blood flow (related to microvessel density) may also contribute to a certain extent to the higher uptake of radioactivity in the small U87MG tumors than in the large tumors.
CONCLUSION
We report herein, to our knowledge, the first example of 64Cu-labeled VEGF121 for PET of tumor VEGFR expression. MicroPET revealed rapid and high 64Cu-DOTA-VEGF121 uptake in small U87MG tumors but very low uptake in large tumors, corresponding with the VEGFR2 expression level in vivo. VEGFR specificity in vivo was confirmed by blocking experiments and ex vivo studies. The success of VEGFR-specific tumor imaging using 64Cu-DOTA-VEGF121 may be translated into the clinic to characterize the pharmacokinetics, tumor-targeting efficacy, dose optimization, and dose interval of VEGF- or VEGFR-targeted cancer therapeutics. It may also significantly aid in patient stratification and treatment monitoring of VEGFR-targeted cancer therapy.
Acknowledgments
We thank Dr. Zi-bo Li, Andrew R. Hsu, and Lina He for their excellent technical support and the cyclotron team at the University of Wisconsin, Madison, for 64Cu production. We also thank Dr. Philip E. Thorpe and Dr. Sophia Ran at UT Southwestern Medical Center, Dallas, for providing the antimouse VEGFR2 primary antibody. This work was funded by National Institute of Biomedical Imaging and Bioengineering grant R21 EB001785, National Cancer Institute (NCI) grant R21 CA102123, NCI In Vivo Cellular Molecular Imaging Center grant P50 CA114747, NCI Small Animal Imaging Resource Program grant R24 CA93862, NCI Centers of Cancer Nanotechnology Excellence grant 1U54CA119367-01, Department of Defense (DOD) Breast Cancer Research Program IDEA award W81XWH-04-1-0697, DOD Ovarian Cancer Research Program award OC050120, DOD Prostate Cancer Research Program New Investigator award DAMD17-03-1-0143, the Benedict Cassen Postdoctoral Fellowship from the Education and Research Foundation for the Society of Nuclear Medicine, and the Clayton Foundation for Research.
Footnotes
-
↵* Contributed equally to this work.
References
- Received for publication July 13, 2006.
- Accepted for publication September 11, 2006.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Radiolabeling Molecular Biomarkers of Invasive Pituitary Neuroendocrine Tumors: A Systematic Review
- PET Imaging of Receptor Tyrosine Kinases in Cancer
- 64Cu-Labeled Repebody Molecules for Imaging of Epidermal Growth Factor Receptor-Expressing Tumors
- PET Imaging of Tissue Factor in Pancreatic Cancer Using 64Cu-Labeled Active Site-Inhibited Factor VII
- PET Imaging of VEGFR-2 Expression in Lung Cancer with 64Cu-Labeled Ramucirumab
- A Tyrosine Kinase Inhibitor-Based High-Affinity PET Radiopharmaceutical Targets Vascular Endothelial Growth Factor Receptor
- Immuno-PET of Tissue Factor in Pancreatic Cancer
- Specific biomarkers of receptors, pathways of inhibition and targeted therapies: pre-clinical developments
- Targeted Systemic Radiotherapy with scVEGF/177Lu Leads to Sustained Disruption of the Tumor Vasculature and Intratumoral Apoptosis
- Approaches to Multimodality Imaging of Angiogenesis
- Molecular Imaging: 18F-FDG PET and a Whole Lot More
- Serial Noninvasive Targeted Imaging of Peripheral Angiogenesis: Validation and Application of a Semiautomated Quantitative Approach
- Positron Emission Tomography Imaging of Poststroke Angiogenesis
- Multimodality Imaging of IL-18-Binding Protein-Fc Therapy of Experimental Lung Metastasis
- Multimodality Molecular Imaging of Tumor Angiogenesis
- Integrin-targeted imaging and therapy with RGD4C-TNF fusion protein
- Advances in Anatomic, Functional, and Molecular Imaging of Angiogenesis
- Imaging of VEGF Receptor in a Rat Myocardial Infarction Model Using PET
- Monitoring of the Biological Response to Murine Hindlimb Ischemia With 64Cu-Labeled Vascular Endothelial Growth Factor-121 Positron Emission Tomography
- Immuno-PET: A Navigator in Monoclonal Antibody Development and Applications
- Dual-Function Probe for PET and Near-Infrared Fluorescence Imaging of Tumor Vasculature
- In Vivo VEGF Imaging with Radiolabeled Bevacizumab in a Human Ovarian Tumor Xenograft
- Small-Animal PET of Tumor Angiogenesis Using a 76Br-Labeled Human Recombinant Antibody Fragment to the ED-B Domain of Fibronectin
- Multimodality Molecular Imaging of Glioblastoma Growth Inhibition with Vasculature-Targeting Fusion Toxin VEGF121/rGel