


Abstract
Cells can die by several pathways, such as accidental death, apoptosis, autophagy, pyroptosis, and oncosis. These are important in normal physiology and many disease states, such as cancer and cardiovascular disease. Specific biochemical changes occur in cells undergoing apoptosis that provide potential targets for molecular imaging agents. Several of these molecular steps have been evaluated to date, including phosphatidylserine exposure at the extracellular face of the plasma membrane, detected by proteins such as annexin V; caspase activation in the intracellular compartment, detected by labeled enzyme substrates or inhibitors; and mitochondrial membrane potential collapse, detected by reduced levels of phosphonium cations that normally accumulate in healthy mitochondria. Phase I clinical trials have been performed with 1 of these agents, annexin V. Future work will likely include development of new agents that detect targets not exploited by current agents, translational research on the significance of imaging the different forms of cell death, and further improvements in the techniques for labeling existing agents to improve sensitivity and reduce nonspecific background.
… death, The undiscovered country, from whose bourn No traveler returns…
Shakespeare, Hamlet, 3.1.78–80
Although death remains as mysterious to us today as it was to Hamlet long ago, cell death has become progressively less mysterious to scientists in recent years. In this review, I summarize recent progress toward molecular imaging agents that can detect cell death in vivo. In keeping with the format of this series, literature citations will be limited to selected reviews and representative examples of recent work; several excellent reviews are cited that summarize the earlier literature (1–4).
TERMINOLOGY AND CLASSIFICATION OF CELL DEATH
Cell death is central to normal physiology and numerous disease states (5–7). Although cell death is often characterized simply as apoptotic (programmed) or necrotic (accidental), many additional forms of programmed cell death (autophagy, oncosis, pyroptosis, etc.) exist—as many as 11 types according to one classification (8,9). These forms of cell death can be classified by the morphology of the dying cells, the biochemical pathways activated, the disposal mechanisms used, and the extracellular consequences of cell death (normally noninflammatory in apoptosis and autophagy and normally proinflammatory in pyroptosis and oncosis). Although this article focuses on apoptosis, much of the work reviewed is likely to be applicable to other forms of cell death. Considerable cross-talk exists between different forms of cell death, and 2 or more processes may often be activated concurrently (10). Biochemical mechanisms can be shared among different pathways; for example, pyroptosis involves activation of an inflammatory subfamily of caspases homologous structurally and functionally to the caspases involved in apoptosis (5). Similarly, both phosphatidylserine exposure and loss of the mitochondrial membrane potential are not limited to apoptosis (9); thus, both imaging targets will likely be useful for detecting multiple forms of cell death.
CELLULAR BIOCHEMICAL CHANGES DURING CELL DEATH AND MOLECULAR PROBES THAT DETECT THEM
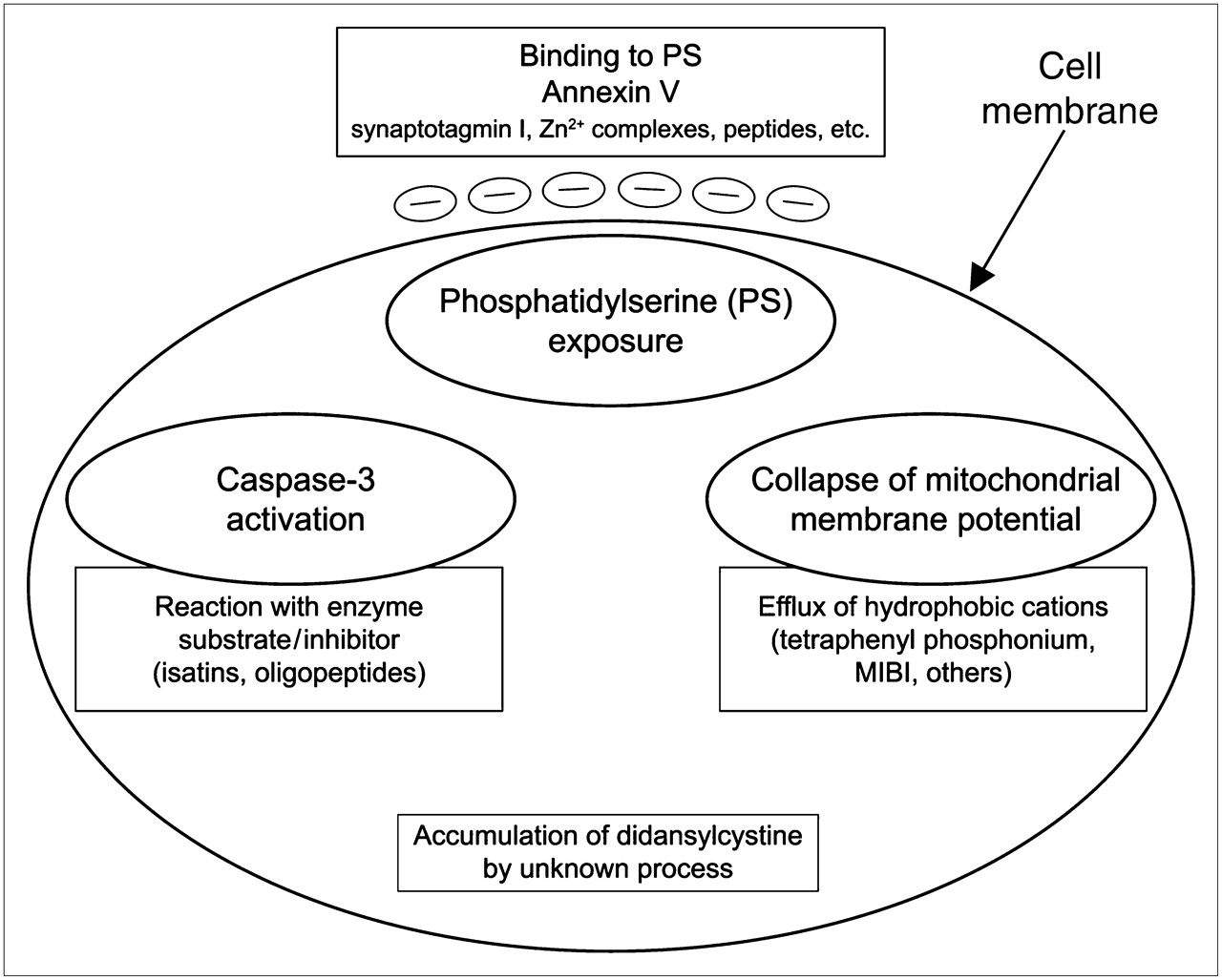
We can classify most apoptosis imaging agents now being investigated into 4 categories on the basis of the cellular processes they detect (Fig. 1).

Illustration of main categories of imaging agents developed to date and cellular processes that they detect. Contents of large ovals are targets inside or outside cell, and contents of large boxes are known agents that can be used to target imaging radionuclides or fluorophores. Phosphatidylserine exposure at extracellular face of cell membrane is represented by small ovals containing negative charge. PS = phosphatidylserine.
Plasma Membrane Phospholipid Asymmetry and Phosphatidylserine Exposure
In healthy cells, phospholipids are asymmetrically distributed, with the anionic phospholipid phosphatidylserine normally confined to the cytoplasmic face of the plasma membrane by an active transport mechanism. This asymmetric distribution of phosphatidylserine can be perturbed either reversibly or irreversibly in a variety of physiologic states, most notably during apoptosis, in which it serves as a primary signal for the phagocytic removal of apoptotic cells (11). Phosphatidylserine exposure also occurs in some other forms of cell death, including autophagy (9). In the final stage of cell death via most pathways, phosphatidylserine on the intracellular face of the plasma membrane also becomes accessible to phosphatidylserine-directed probes in a nonspecific fashion due to complete loss of membrane integrity.
Phosphatidylserine exposure has received the most attention as an imaging target in apoptosis for several reasons. This exposure is a near-universal event in apoptosis, it occurs within a few hours of the apoptotic stimulus, and it presents a very abundant target (millions of binding sites per cell) that is readily accessible on the extracellular face of the plasma membrane (2,4). Annexin V (also known as annexin A5), a natural human phosphatidylserine-binding protein, is by far the most widely used phosphatidylserine-directed agent. The use of annexin V reflects several advantages, including the very high affinity for apoptotic cells (low nanomolar to subnanomolar dissociation constant values), ready production by recombinant DNA technology, and lack of in vivo toxicity (1,2,4) of the protein. Several other protein probes against phosphatidylserine are under investigation. The C2A domain of synpaptotagmin I has a reasonably high affinity for phosphatidylserine and has been used in several recent studies (12). Another novel approach has been the development of small-molecule Zn2+ coordination complexes directed against cell-surface phosphatidylserine (13). Another report describes a 12-residue phosphatidylserine-binding peptide discovered by screening a phage library of random peptides (14). These alternative approaches are promising, but at this point they lack the base of experience in human and animal studies that is present with annexin V and its derivatives.
Some recent developments with annexin V illustrate the potential for improvements in this imaging agent and more general trends in protein-based molecular imaging agents (Fig. 2). A short N-terminal peptide was introduced into the protein sequence via recombinant DNA methods; this then allowed both direct chelation of 99mTc (15) and site-specific derivatization with an 18F maleimide compound (16), multiple fluorophores (17), or the metal chelator 1,4,7,10-tetraazacyclododecane-N,N′,N″,N′″-tetraacetic acid (Jonathan F. Tait, D. Scott Wilbur, Ming-Kuan Chyan, Donald K. Hamlin, Christina Smith, and Donald F. Gibson, unpublished data, 2008). This approach potentially allows multiple imaging modalities to be based on the same molecular platform and was also shown to double the in vivo uptake on apoptotic tissues via improvements in binding affinity for apoptotic cells relative to randomly modified forms of annexin V (18). Recent structure–function studies on annexin V have also clarified the mechanism of binding, providing detailed guidance on how the molecule can be modified for imaging without compromising bioactivity (17).
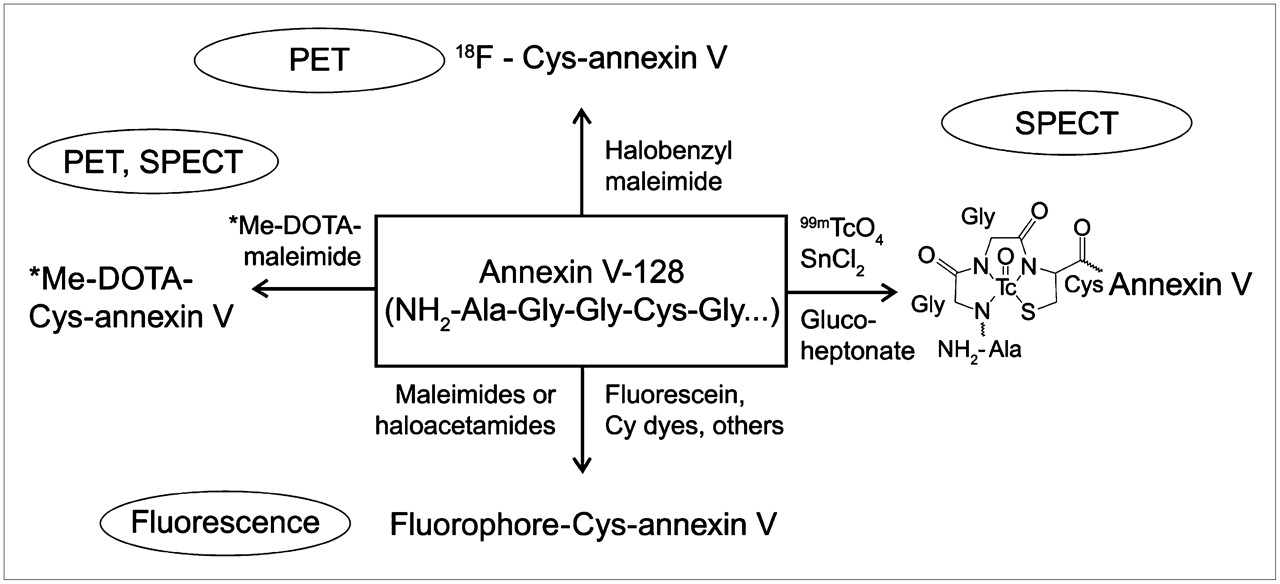
Example of contemporary protein-based apoptosis imaging platform. Recombinant phosphatidylserine-binding protein called annexin V-128 has 6-residue N-terminal peptide built into its structure that allows site-specific labeling with 99mTc via direct chelation and attachment of various positron-emitting isotopes and fluorophores via reaction with unique N-terminal cysteine residue. This allows imaging via variety of modalities. Me represents metal ion, for example, 111In, 64Cu, 117mSn, or other radionuclides, useful for imaging.
Caspase Activation
Caspases are attractive targets because of their central role in the execution of cell death (19). Both the intrinsic (mitochondrial) and extrinsic (death receptor) pathways of apoptosis eventually activate several effector caspases (19). Several groups have recently described molecules that target caspase-3, one of the key effector caspases (20–22). Although work on molecules that target caspase-3 is promising, there are limited animal data so far and no human data, and it is not yet known how sensitive these agents will be for actual imaging in humans.
Mitochondrial Membrane Potential Collapse
One interesting category of compounds that has received relatively little attention is lipophilic cations that accumulate in healthy mitochondria because of the strongly negative mitochondrial transmembrane potential produced during normal oxidative metabolism. Collapse of the mitochondrial electrochemical potential is one of the central events in apoptosis (7) and a potentially abundant target because cells contain numerous mitochondria. Radiolabeled probes based on phosphonium cations show the predicted decrease in cellular uptake in vitro as mitochondrial potential is decreased, and in vivo these probes show highest uptake in the heart and kidneys (23,24). This approach is different because apoptosis would be registered as a decrease in uptake, rather than an increase in uptake as with other categories of probes discussed here. Thus, these probes might work best in organs such as the heart and kidneys, with a high density of mitochondria, but might work less well in organs with lower levels of metabolic activity. Interpretation of results may also be complicated by cellular efflux mediated by the multidrug-resistance proteins, which occurs to variable degrees depending on the specific structure of the compound involved (24).
Uncategorized
Two interesting compounds, with the trade name Apo-Sense, have recently been described (25,26): N,N′-didansylcystine and a closely related dansylated compound code named NST-732 ([5-dimethylamino]-1-naphthalenesulfonyl-α-ethyl-fluoroalanine). Both accumulate modestly (about 2-fold) in apoptotic cells. Aloya et al. speculate that these compounds are excluded from normal cells but enter apoptotic cells when membrane phospholipids are scrambled early in apoptosis (25). However, this mechanism is unproven, and at this point the use of these probes is more empiric than mechanistically based. The doses of these agents are also 100- to 1,000-fold higher than doses for other classes of agents, raising concern about potential toxicity in dose ranges more typically associated with therapeutic drugs rather than radiopharmaceutical tracers.
CLINICAL APPLICATIONS AND FUTURE PROSPECTS
Research over the last decade has produced an abundant selection of potential probes, both proteins and small organic molecules, which are in various stages of development and evaluation. Many areas of clinical medicine could potentially benefit from effective imaging of cell death in vivo. For example, imaging of apoptotic response could provide a much faster way to predict effectiveness of cancer chemotherapy than currently used morphologic measurements (1). In cardiovascular medicine, imaging of apoptosis could be highly useful in managing myocardial infarction, unstable atherosclerotic plaques, and cardiac allograft rejection (3). However, enthusiasm for these applications must be tempered by the inherent delays and difficulties of developing a new class of imaging agents for clinical use and the recognition that no agent has yet progressed to Federal Drug Administration approval.
What can we expect in the future? First, numerous biochemical features of apoptosis remain unexploited, providing great scope for developing new classes of imaging agents, which may turn out to be inherently superior to existing classes of agents. Second, translational research is needed to evaluate the variety of cell death pathways and their potential significance for imaging in the diagnosis or monitoring of disease. Third, for protein-based agents, site-specific labeling is a preferable approach to older methods of random labeling and will likely improve the quality of imaging for many of the protein-based agents in use or being developed. Also, the biodistribution and metabolism of labeled proteins is often very different from that of small molecules, requiring novel approaches to reducing the impact of nonspecific uptake or metabolism on the quality of images. For some probes, it will be important to gain a better mechanistic understanding of how they work. Clearly, much needs to be done before the bourn of cell death is no longer shrouded in mystery.
Acknowledgments
I thank Dr. Scott Wilbur for helpful comments on the manuscript. This work was supported by U.S. Public Health Service grant CA-102348.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication July 7, 2008.
- Accepted for publication August 7, 2008.




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- 18F-ML-10, a PET Tracer for Apoptosis: First Human Study
- Apoptosis Imaging: Beyond Annexin V
- Small-Molecule Biomarkers for Clinical PET Imaging of Apoptosis
- Time Course of Paclitaxel-Induced Apoptosis in an Experimental Model of Virus-Induced Breast Cancer
- Detection and Quantification of the Evolution Dynamics of Apoptosis Using the PET Voltage Sensor 18F-Fluorobenzyl Triphenyl Phosphonium