


Abstract
Radiolabeled cyclic peptides containing the amino acid sequence arginine-glycine-aspartate (RGD peptides) have successfully been used to image the expression of the αvβ3 integrin in malignant tumors. However, the αvβ3 integrin also plays an important role in angiogenesis induced by chronic inflammatory processes. Therefore, the aim of this study was to evaluate whether radiolabeled RGD peptides may also be used to assess αvβ3 expression in inflammatory diseases. We studied a hapten-induced delayed-type hypersensitivity reaction (DTHR) as a model for inflammatory processes, since DTHRs are involved in many human autoimmune disorders. Methods: The abdominal skin of mice was sensitized by application of 2,4,6-trinitrochlorobenzene (TNCB). One week later, a DTHR was elicited by challenging the right ear with TNCB. Application of TNCB was then repeated every 48 h to induce chronic skin inflammation. Small-animal PET and autoradiography with the αvβ3 ligands 18F-galacto-RGD and 125I-gluco-RGD were performed at various times after TNCB application. The time course of tracer uptake by the treated ears was compared with histologic skin changes. Results: The first challenge with TNCB caused, within 12 h, an acute inflammatory response with dense dermal infiltrates of polymorphonuclear leukocytes and lymphocytes. However, autoradiography revealed no significant increase in 125I-gluco-RGD uptake at that time (mean uptake ratio for treated ear to untreated ear, 1.02 ± 0.1 [SD]). Further challenges with TNCB resulted in chronic skin inflammation with markedly increased small-vessel density in the ear tissue. This was paralleled by a continuous increase in uptake of 125I-gluco-RGD. After 13 challenges, the uptake ratio had increased to 2.30 ± 0.27 (P < 0.005 compared with baseline). Enhanced uptake of radiolabeled RGD peptides in chronic inflammation was also demonstrated noninvasively by PET with 18F-galacto-RGD. Pretreatment of the mice with nonradiolabeled cyclic peptide c(RGDfV) almost completely blocked uptake of 18F-galacto-RGD by the challenged ear, thus confirming the specificity of tracer uptake. Conclusion: Radiolabeled RGD peptides allow a noninvasive assessment of αvβ3 expression in inflammatory processes. PET with 18F-galacto-RGD might become a powerful tool to distinguish between the acute and chronic phases of T cell–mediated immune responses and may represent a new biomarker for disease activity in autoimmune disorders.
Integrins, a large family of cell adhesion molecules, are the main mediators of the binding and response of cells to the extracellular matrix. Each integrin is composed of 2 noncovalently associated transmembrane glycoprotein subunits called the α- and the β-chains. So far, 18 α- and 8 β-chains have been identified, which form at least 24 receptors with different substrate specificities (1). One of the best characterized of these receptors is the αvβ3 integrin, which plays an important role for endothelial cell migration and survival during angiogenesis. In contrast, αvβ3 is found only in minimal amounts in resting endothelial cells. Tumor cells of various origins, particularly melanomas, can also express αvβ3. In normal tissues, expression of αvβ3 is highly restricted, and significant amounts of this integrin have been observed only in osteoclasts (2). This limited expression by normal tissues makes αvβ3 a particularly attractive target for imaging probes.
The binding mechanisms of αvβ3 integrin to its ligands (e.g., vitronectin, fibronectin, fibrinogen, and osteopontin) have been studied extensively in recent years (3). It is now well established that αvβ3 binds to matrix proteins with an exposed RGD (arginine-glycine-aspartate) sequence (4). This binding motif is also used by several other integrin subtypes (e.g., αvβ5 and αIIbβ3) (3). However, the binding affinity of different integrins to proteins containing an RGD motif is critically influenced by the 3-dimensional arrangement of the 3 amino acids in this sequence as well as by the flanking amino acid residues (3). Intensive structure/activity investigations have resulted in the development of cyclic pentapeptides that specifically bind to αvβ3 but not to other related RGD-binding integrins (5). Radiolabeling of these peptides with radioiodine, 111In, 99mTc, and 18F, has allowed the development of various imaging agents that specifically localize to tumors expressing αvβ3 (6).
αvβ3 is also highly expressed in endothelial cells during wound healing (7), in restenosis after angioplasty (8), in the synovia of patients with rheumatoid arthritis (9), and in psoriatic plaques (10). In an experimental model of rheumatoid arthritis, treatment with the cyclic RGD peptide c(RGDfV) significantly inhibited disease progression (11). Furthermore, αvβ3 antagonists have been shown to inhibit bone resorption by osteoclasts. Antibodies against αvβ3 or cyclic RGD peptides are, therefore, currently being evaluated as a new therapeutic approach for rheumatoid arthritis (12). Thus, radiolabeled RGD peptides may be used to characterize not only malignant tumors but also inflammatory diseases.
To evaluate uptake of radiolabeled RGD peptides by inflammatory processes, we studied a murine model for the cutaneous delayed-type hypersensitivity reaction (DTHR). We selected this model because DTHRs are involved in several common autoimmune disorders in humans, such as rheumatoid arthritis or chronic inflammatory bowel disease. In these diseases, DTHRs are directed against peptide antigens present within the tissues. However, cutaneous DTHR may also be induced by haptens applied to the skin. In our experiments, we used the hapten 2,4,6-trinitrochlorobenzene (TNCB) to induce contact hypersensitivity reactions (CHSRs), which have been well characterized in previous studies (13,14). In mice previously sensitized to TNCB, the first challenge with TNCB at one ear causes acute skin inflammation (acute CHSR), and the other, nontreated, ear is used as a control. Repeated challenges with TNCB result in chronic skin inflammation (chronic CHSR). Thus, this model allowed us to study the expression of αvβ3 in acute and chronic inflammation. As imaging probes, we used the glycosylated RGD peptides 125I-gluco-RGD and 18F-galacto-RGD. These peptides show almost the same affinity and specificity for the αvβ3 integrin. They also demonstrate a similar biodistribution in tumor-bearing mice (15,16). PET with 18F-galacto-RGD has recently been shown to allow imaging of αvβ3 expression in patients with various malignant tumors (17). In the present study, 125I-gluco-RGD was used for autoradiographic studies and 18F-galacto-RGD for PET.
MATERIALS AND METHODS
Animals
All animal experiments were performed in accordance with the guidelines of the Technische Universität München for the use of living animals in scientific studies and the German Law for the Protection of Animals. The C57BL/6 mice (Charles River Laboratories) were between 8 and 12 wk old.
In Vivo Experiments
Mice were sensitized by application of 80 μL of 5% TNCB (dissolved in a 4:1 mixture of acetone and olive oil, w/w) to the shaved abdomen (size, ∼2 × 2 cm). One week later, the mice were challenged with 20 μL of 1% TNCB (dissolved in a 1:9 mixture of acetone and olive oil, w/w) on both sides of the right ear, to elicit acute TNCB-specific CHSR. In the 5% TNCB solution acetone is an irritant, whereas it is used solely as a solvent in the 1% TNCB solution. To induce chronic cutaneous DTHR, mice (n = 3–6) were recurrently challenged at the ear every 2 d for a total of up to 13 challenges. Elicitation of CHSR at an ear allows easy assessment of the degree of inflammation through measurement of ear thickness with a micrometer (Oditest; Kroeplin) before the first TNCB challenge and 12 h after every subsequent ear challenge. TNCB was purchased from Fluka (Switzerland), 18F was produced with an 11-MeV RDS cyclotron (CTI) at the Technische Universität München, and 125I was purchased from Amersham-Buchler.
Synthesis of Gluco- and Galacto-RGD
125I-gluco-RGD and 18F-galacto-RGD were synthesized as described previously (15,18). Briefly, the linear peptides aspartate-tyrosine-lysine-arginine-glycine and aspartate-phenylalanine-lysine-arginine-glycine were assembled on solid support using standard 9-fluorenylmethoxycarbonyl (Fmoc) protocols and cyclized under high dilution conditions. Thereafter, they were conjugated with the benzylated glucose-based sugar amino acid Ac-SAA1(Bn3)-OH (15) and Fmoc-protected galactose-based sugar amino acid Fmoc-SAA2-OH (15), respectively. 125I-gluco-RGD was labeled with no-carrier-added 125I-NaI by the IODO-GEN method (Pierce). The radiochemical purity as assessed by high-performance liquid chromatography was >95%, and the specific activity was >10 TBq/mmol. 18F-galacto-RGD was labeled using no-carrier-added 4-nitrophenyl-2-18F-fluoropropionate. The radiochemical purity of the final product was >98% as assessed by high-performance liquid chromatography, and the specific activity was >40 TBq/mmol (15,18).
Autoradiography
For digital phosphor storage autoradiography, 370 kBq of 125I-gluco-RGD were intravenously injected into the tail vein and the animals were sacrificed 3 h afterward. During the uptake period, the mice were not kept anesthetized. Immediately after dissection, the ears were embedded in optimal cutting temperature compound (TissueTek; Fisher Scientific) and frozen in liquid nitrogen. Autoradiography was performed with the phosphor imager 445SI (Amersham Bioscience/Molecular Dynamics) from 20-μm fresh-frozen sections of the left, nonchallenged, and right, challenged, ears. Autoradiographic studies were performed on groups of 3–6 mice 12 h (time of animal scarification) after the first, second, fourth, eighth, tenth, and thirteenth challenges. Further autoradiograms were obtained for a group of 3 mice 24 h (time of animal scarification) after the first challenge to observe potential dynamic changes in tracer uptake at different times. The autoradiography data were analyzed with ImageQuant software (version 5.1; Molecular Dynamics). For each mouse rectangular regions of interest of the same size were drawn around 3 sections of the left and right ears. For further analysis, the mean value of the 3 autoradiograms per ear was used to calculate tracer uptake.
Histology and Immunohistochemistry
Histologic staining with hematoxylin and eosin was performed on 5-μm cryosections. In addition, vascular endothelial cells were stained with a platelet endothelial cell adhesion molecule-1 (PECAM-1, cluster designation 31 [CD31]) rat-antimouse antibody (BD Pharmingen) to allow visualization of vascularization. As a control antibody, rat-Igb2ακ (BD Pharmingen) was used. Staining was performed according to the instructions of the manufacturer.
High-Resolution Animal PET
PET was performed using the Munich Avalanche Diode PET (MADPET) (19), a prototype small-animal positron tomograph. The scanner consists of 2 sectors comprising 3 detector modules each, which rotate around the animal for acquisition of complete projections in 1 transaxial slice (30 angular steps). List-mode data are reconstructed using statistical, iterative methods including the spatially dependent line-spread function. Reconstructed image resolution is about 2.5 mm (full width at half maximum) in a transaxial field of view of about 5 cm. A 9.25-MBq aliquot of 18F-galacto-RGD was injected into the tail vein 2 h before the 20-min PET scan. The animals were kept under anesthesia (intraperitoneal injection of 80 mg of Ketanest [Parke-Davis] and 10 mg of xylazine per kilogram of body weight) during scanning. MADPET has an axial field of view of only 3.7 mm; thus, the mice were scanned supine, with the neck gently flexed to the thorax so that only the ears were in the field of view. To estimate relative tracer uptake by the left and right ears, we manually drew regions of interest around both ears and calculated the ratio between mean counts for the right and left ears.
Statistical Analysis
All quantitative results are presented as the mean value ± SD. A t test result with a 2-tailed P value of less than 0.05 was considered statistically significant.
RESULTS
Morphologic Changes During Acute and Chronic Inflammation
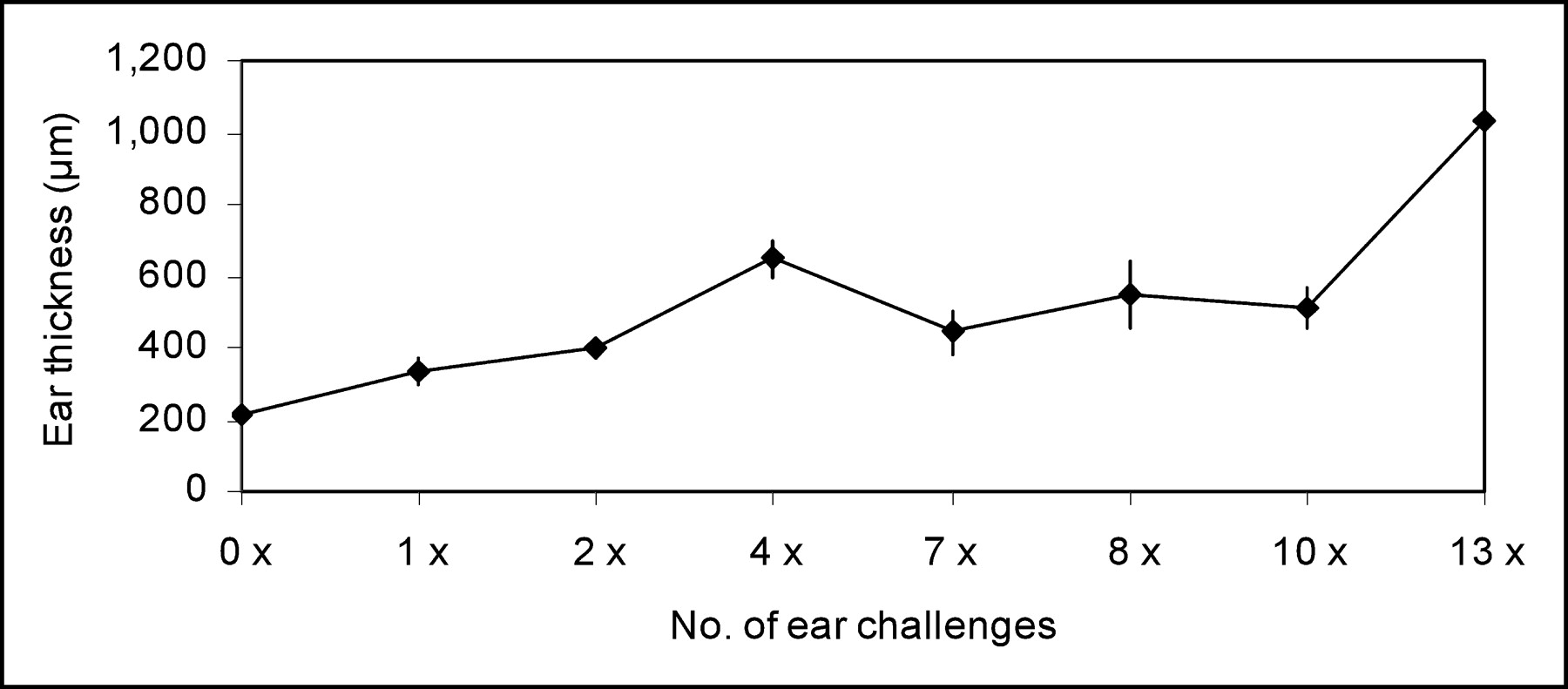
Twelve hours after the first elicitation of acute cutaneous DTHR, there was edematous ear swelling, resulting in an increase of ear thickness from 210 ± 9 μm to 331 ± 37 μm (P < 0.00001) at the site of TNCB challenge. Repeated challenges at the ear caused TNCB-specific chronic inflammation, which was associated with a stepwise increase in ear thickness (Fig. 1). The average thickness of the treated ears increased from 210 ± 9 μm (no challenge) to 1,037 ± 81 μm (12 h after the 13th challenge) (P < 0.003 compared with baseline). Almost no signs of inflammation and no increase in ear thickness were found in ears from naïve mice treated once with TNCB, confirming that the inflammatory reaction was a TNCB-specific immune response (Fig. 2A).

Time course of ear thickness 12 h after TNCB challenge. Error bars denote 1 SD.
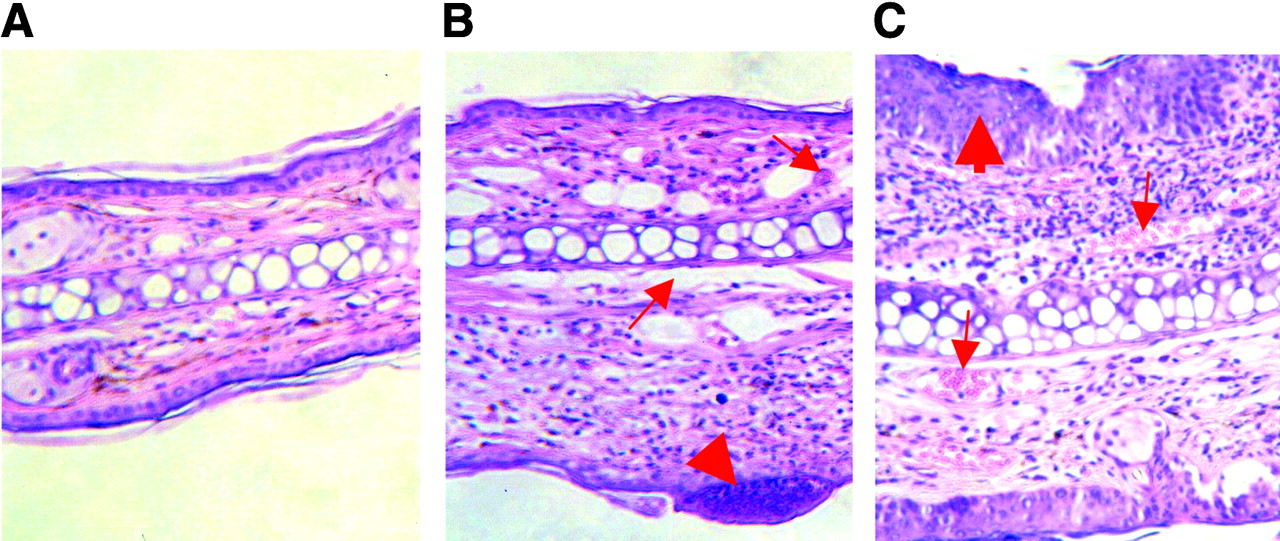
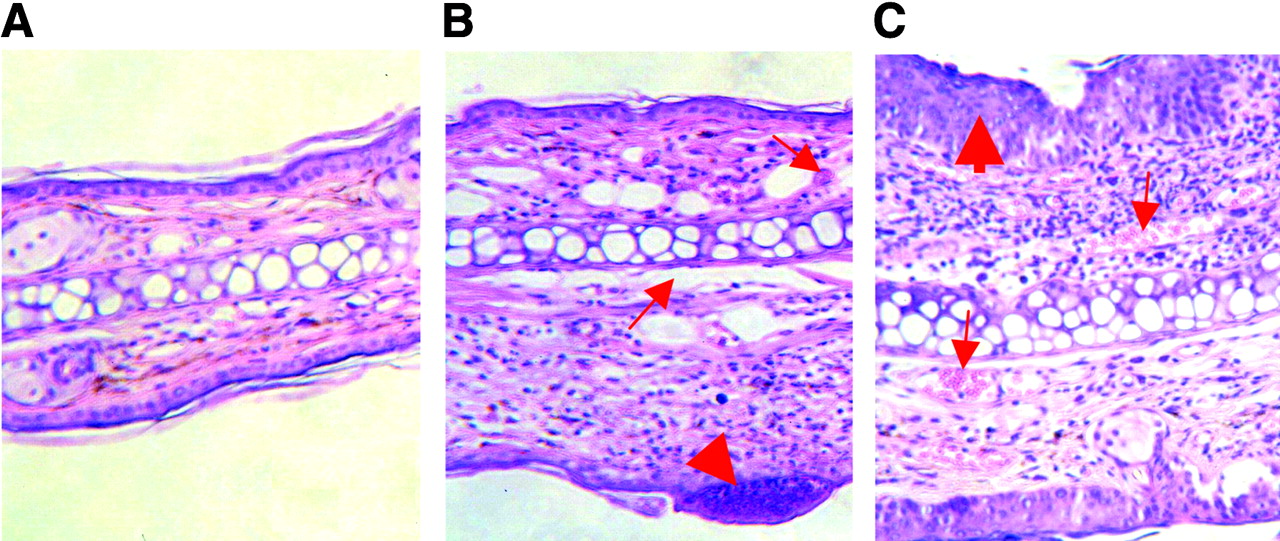
Hematoxylin- and eosin-stained ear sections of control mouse (A) and TNCB-sensitized mice (B and C) show morphologic changes 12 h after a single TNCB challenge (B) and after 10 challenges (C). B and C show, respectively, acute and chronic contact hypersensitivity reactions. Thick arrow in B indicates infiltrates by polymorphonuclear leukocytes and lymphocytes; thin arrows indicate dilated vessels. Thick arrow in C indicates epidermal hyperplasia and hyperkeratosis; thin arrows indicate blood vessels.
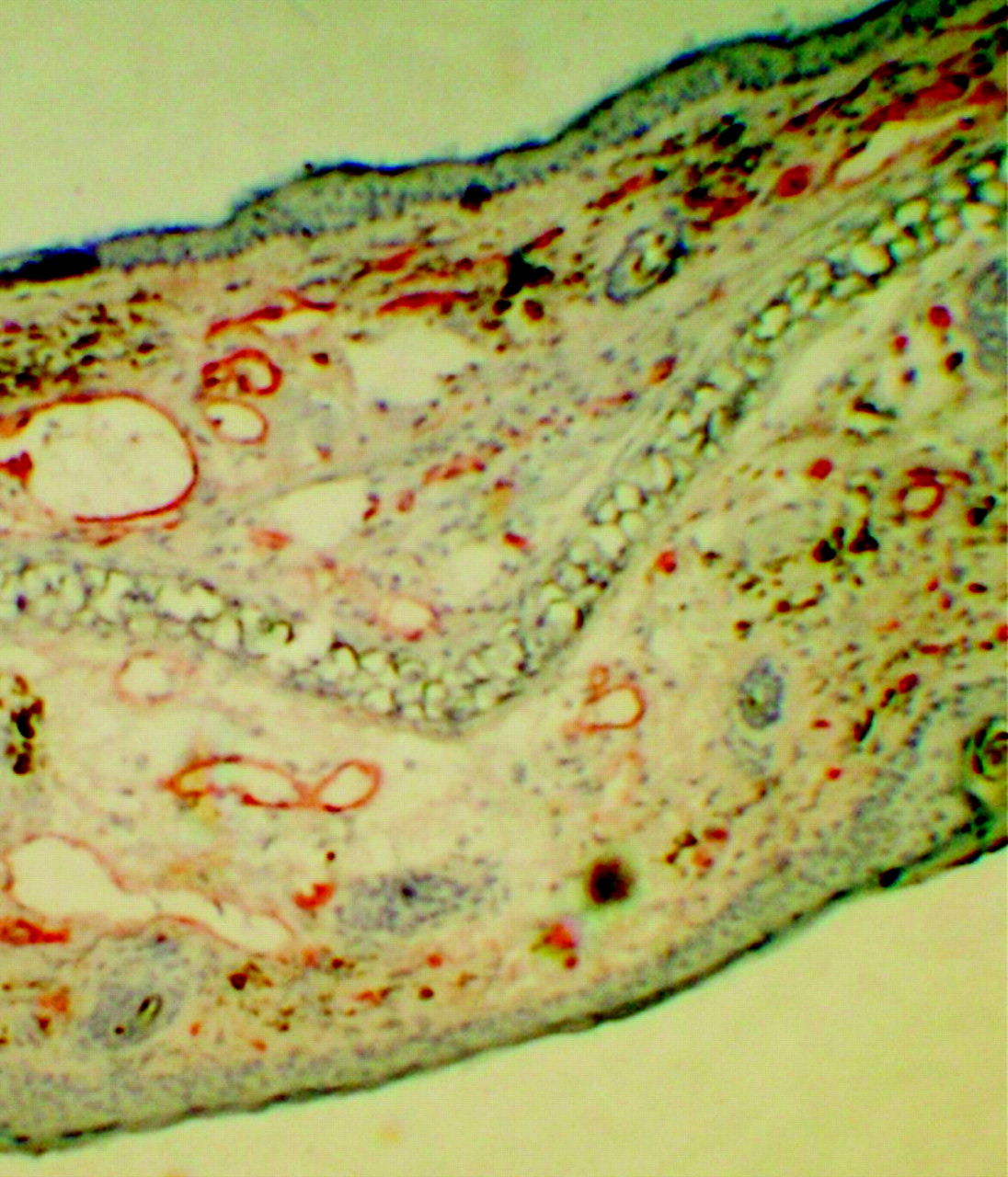
Histologic examination of hematoxylin- and eosin-stained ear tissue 12 h after the first TNCB challenge of sensitized mice confirmed the presence of typical changes found in acute inflammatory responses, such as a dense dermal infiltration of polymorphonuclear leukocytes and lymphocytes, as well as the formation of subepidermal abscesses (Fig. 2B). Furthermore, extensive dilation of blood vessels and strong interstitial edema were found. In contrast, chronic TNCB-specific inflammation was associated with acanthosis, hyperkeratosis, and an increase of the number of new blood vessels, whereas the interstitial edema was less pronounced (Fig. 2C). PECAM-1 (CD31) immunohistochemistry confirmed the extensive neovascularization after the 10th challenge (Fig. 3).

CD31 immunohistochemistry of ear section 12 h after 10th TNCB challenge demonstrates extensive neovascularization.
RGD Peptides as Specific Markers for Chronic Inflammation
Twelve hours after the TNCB challenge, autoradiography demonstrated only a minimally increased uptake of 125I-gluco-RGD in the challenged right ear, compared with the untreated left ear (Fig. 4). Quantitatively, the ratio of tracer uptake by the right and left ears was 1.03 ± 0.2 (P = not significant; Fig. 5). Tracer uptake also remained low at 24 h after the first TNCB challenge (uptake ratio for treated ear to untreated ear, 1.02 ± 0.01). Repeated challenges with TNCB caused chronic inflammation, resulting in a steady increase in 125I-gluco-RGD uptake (Fig. 5). After 13 challenges with TNCB, the ratio of tracer uptake of the treated ear to the untreated ear was 2.30 ± 0.27 (P < 0.005 compared with baseline; Fig. 5).

Typical autoradiograms of 20-μm sections from left (L) and right (R) mouse ears. (A) TNCB-sensitized mouse after first TNCB challenge (acute contact hypersensitivity). (B) TNCB-sensitized mouse after 10th challenge with TNCB.
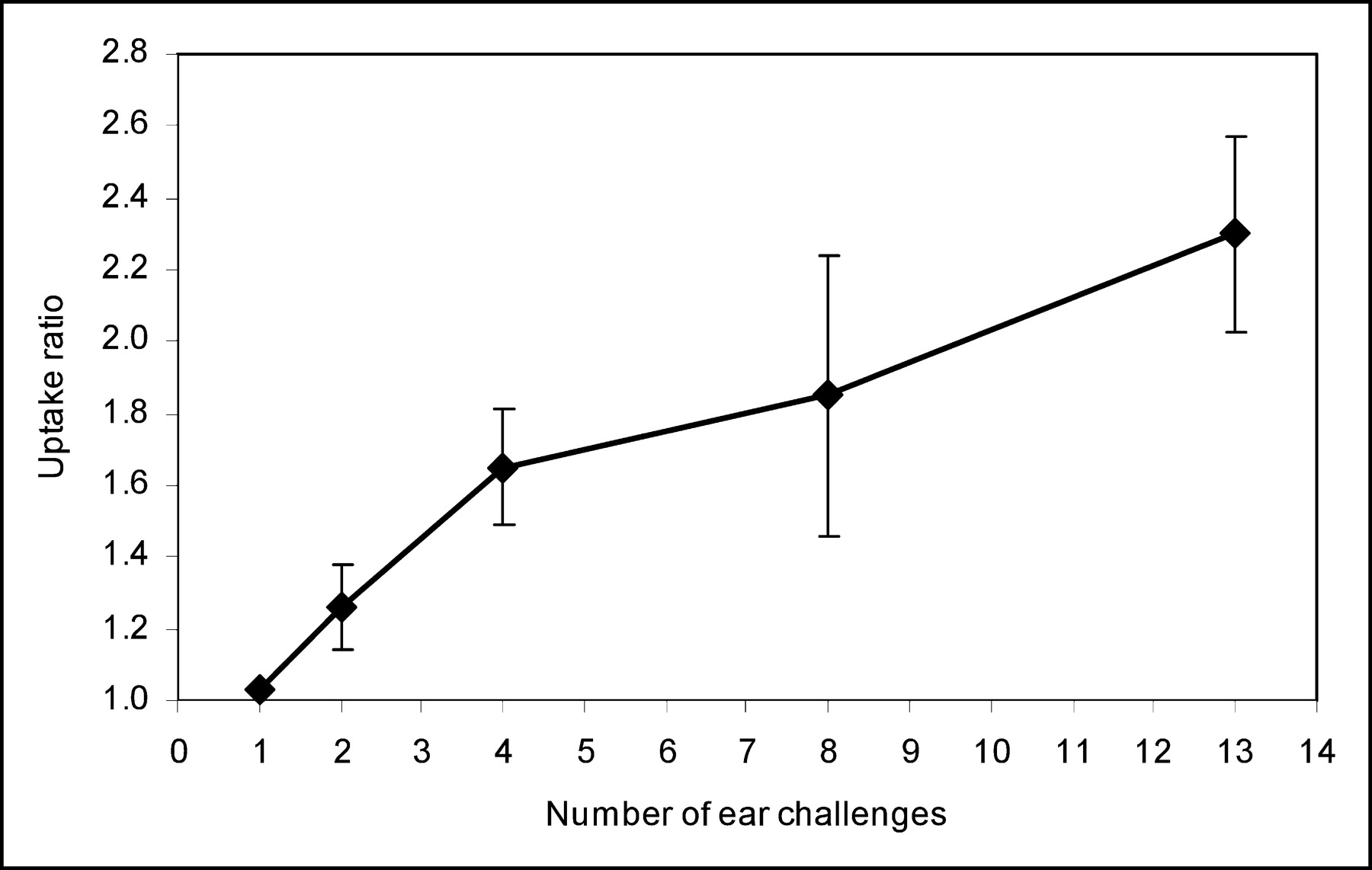
Time course of 125I-gluco-RGD uptake ratio of challenged right ear and nonchallenged left ear versus number of challenges. Error bars denote 1 SD.
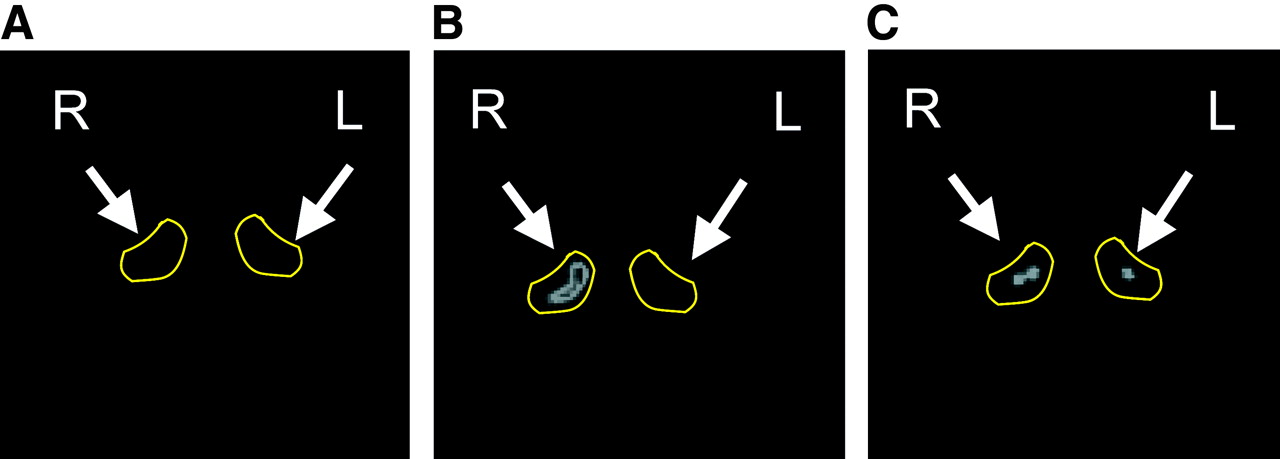
Increased tracer uptake of radiolabeled RGD peptide after repeated challenges with TNCB was also seen on PET imaging with 18F-galacto-RGD (Fig. 6). In untreated mice, the ears were not seen in the PET images. After 1 challenge, only minimal tracer uptake in the treated ear was visible (Fig. 6A). However, after 10 challenges there was marked focal uptake of 18F-galacto-RGD in the treated ear. The intensity of tracer uptake was about 4.7 times higher in the challenged right ear than in the untreated ear (Fig. 6B). Most important, when mice were intravenously injected with nonradiolabeled c(RGDfV), 18 mg/kg, 10 min before injection of 18F-galacto-RGD, tracer uptake by the challenged ear could be reduced to background levels (Fig. 6C), confirming the specificity of 18F-galacto-RGD uptake.
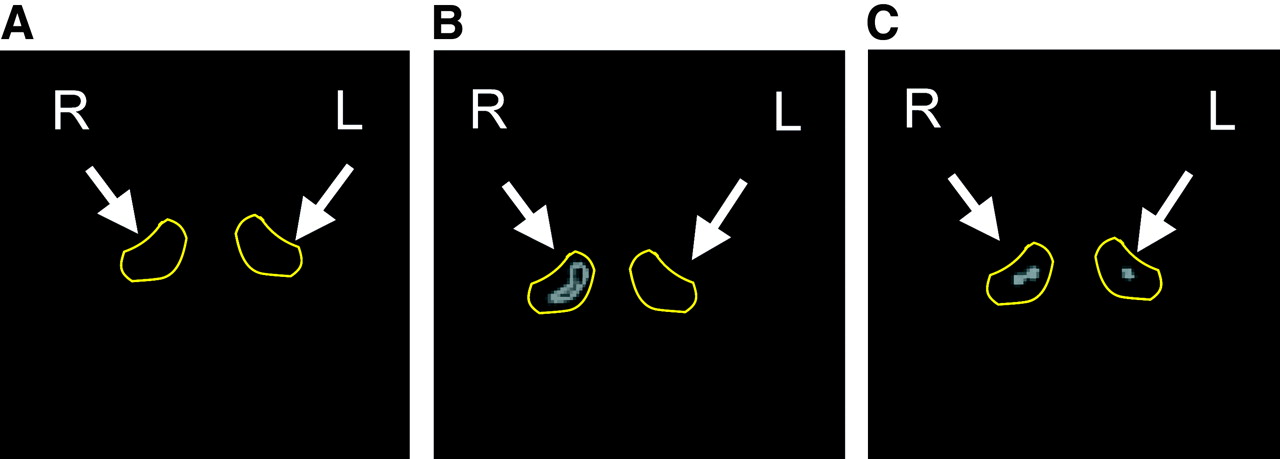
High-resolution PET images of challenged right ear and nonchallenged left ear with 18F-galacto-RGD. Images are of mouse after 1 challenge (A), after 10 challenges (B), and after 10 challenges and blocking of αvβ3-receptors with unlabeled c(RGDfV) peptide, 18 mg/kg (C).
DISCUSSION
We used a TNCB-induced CHSR as a model to study αvβ3 integrin expression during acute and chronic inflammatory processes. CHSR can be divided into a sensitization phase, which is the priming of specific interferon-γ–producing T helper 1 and cytotoxic T lymphocyte cells, and an elicitation phase, which is the induction of T cell–mediated inflammation. Sensitization requires application of a hapten, such as TNCB, in combination with an irritant, such as acetone, to activate epidermal Langerhans’ antigen-presenting cells. Activated hapten TNCB-carrying Langerhans’ cells then traffic through afferent lymphatic vessels of the dermis to the draining axillary and inguinal lymph nodes. Inside the paracortical lymph node area, Langerhans’ cells attract T cells and present hapten to the T cells, inducing TNCB-specific T cell activation and 1,000- to 100,000-fold T cell proliferation. These activated TNCB-specific T helper 1 and T lymphocyte cells can then migrate from the lymph nodes into the bloodstream.
Seven days after the initial sensitization, specific CHSR can be elicited by exposure of TNCB in a nonirritant oil solution on the right ear. This second TNCB exposure, the “challenge,” attracts and activates the TNCB-specific T helper 1 and T lymphocyte cells at the site of challenge. This process is associated with rapid proliferation of TNCB-specific T helper 1 and cytotoxic T lymphocyte cells and cytokine production, specifically of interleukin-2, interferon-γ, and tumor necrosis factor (13), inducing a cascade of events that involve stimulation of multiple other cell types, including polymorphonuclear leukocytes, endothelial cells, mast cells, and keratinocytes (20,21).
Repeated exposure to TNCB at the same site induces a chronic inflammatory response associated with downregulation of interferon-γ production and upregulation of interleukin-4 production (13,22). The chronic TNCB-specific inflammatory process is accompanied by dense cellular infiltration, marked neovascularization, and a continuous increase in ear thickness due to acanthosis, hyperkeratosis, and interstitial edema.
Angiogenesis is a typical component of chronic CHSR. Several studies have shown expression of vascular endothelial growth factor and its receptors during the elicitation phase of CHSR (23). Blood vessels in chronic CHSR are dilated and demonstrate increased permeability to macromolecules, typical features of angiogenesis (24). Knockout mice lacking the angiogenesis inhibitor thrombospondin show more intense and prolonged CHSR reactions (25). In our study, increased microvessel density in the skin of the repeatedly challenged ear was documented by immunohistochemistry. Involvement of the αvβ3 integrin was determined using radiolabeled glycosylated RGD peptides. It has already been shown that this class of tracer allows noninvasive determination of αvβ3 expression in murine tumor models (15,16) and in patients (17). Therefore, the accumulation of RGD peptides observed in our study most likely reflects αvβ3 expression on activated endothelial cells during angiogenesis. In mice, however, immunohistochemical staining of the αvβ3 integrin is not feasible because of the lack of specific antibodies directed against murine αvβ3. Because αvβ3 can also be expressed on activated macrophages, lymphocytes, and granulocytes (26,27), we cannot fully exclude that the uptake of gluco- or galacto-RGD in chronic CHSR was also caused by accumulation in these cell types. However, tracer uptake was significantly greater in the treated ear than in the untreated ear only after the third challenge with TNCB. In contrast, histologic evaluation had already demonstrated dense leukocyte infiltrates 12 h after the first challenge, suggesting that αvβ3 expression by leukocytes is not a major factor for RGD uptake in CHSR.
Inflammatory processes may cause nonspecific uptake of numerous radiopharmaceuticals. This effect is due to regional increases in blood flow, vascular permeability, and extracellular volume (28). Therefore, it is particularly important to assess the specificity of tracer uptake in inflammatory processes. Two findings of our study indicated that uptake of gluco- and galacto-RGD was not due to unspecific mechanisms but to specific binding to the αvβ3 integrin. First, we did not observe increased uptake of gluco- or galacto-RGD in the acute phase of CHSR, although this phase is characterized by a marked increase in vascular permeability and edema formation. This finding suggests that unbound peptides had been washed out of the inflamed tissue by the time the imaging was performed. Second, tracer uptake in the chronic phase of CHSR was nearly completely blocked by pretreatment with an αvβ3 antagonist, confirming the receptor-specific tracer accumulation.
Because of the thinness of the normal mouse ear, the tracer uptake ratio calculated from the PET studies was clearly influenced by partial-volume effects: Tracer uptake in the treated ear might be overestimated relative to the untreated ear, because the treated ear is up to 5 times thicker than the untreated ear. However, we observed a significant difference in tracer uptake between the treated and untreated ears not only in the PET studies but also in autoradiograms of cryosections of identical thickness. Therefore, the possibility that the increased tracer accumulation found in the chronic CHSR is simply due to partial-volume effects can be excluded. Hence, our study indicated that radiolabeled RGD peptides specifically accumulate in the chronic inflamed tissue because of receptor-specific binding and that this process can be monitored noninvasively by PET.
CONCLUSION
Our data indicate that PET imaging with 18F-galacto-RGD may be used to evaluate αvβ3 expression in inflammatory disorders and may represent a new biomarker for disease activity and for the monitoring of therapeutic interventions. In addition, this study showed, for what is to our knowledge the first time, that PET with radiolabeled RGD peptides might be a powerful tool to distinguish between acute and chronic T cell–mediated immune responses. Thus, further clinical studies of galacto-RGD for imaging of inflammatory processes in humans appear to be justified.
Acknowledgments
The authors thank Claudia Bodenstein, Wolfgang Linke (Nuklearmedizinische Klinik der Technischen Universität München), Dagmar Dick, Sebastian Harrasser, and Doris Jakob (Department of Dermatology, Ludwig-Maximilians-University, Munich) for excellent technical support. The work was supported by Wilhelm-Sander-Stiftung (97.041.2) and the Deutsche Krebshilfe (10-1917).
Footnotes
Received Mar. 19, 2004; revision accepted Sep. 13, 2004.
For correspondence or reprints contact: Bernd J. Pichler, PhD, Department of Radiology, University of Tübingen, Hoppe-Seyler-Strasse 3, 72076 Tübingen, Germany.
E-mail: Bernd.Pichler{at}med.uni-tuebingen.de
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- 68Ga-DOTA-E[c(RGDfK)]2 PET Imaging of SHARPIN-Regulated Integrin Activity in Mice
- PET/CT Imaging of Integrin {alpha}v{beta}3 Expression in Human Carotid Atherosclerosis
- Positron Emission Tomography/Computed Tomographic and Magnetic Resonance Imaging in a Murine Model of Progressive Atherosclerosis Using 64Cu-Labeled Glycoprotein VI-Fc
- 99mTc-3PRGD2 for Integrin Receptor Imaging of Lung Cancer: A Multicenter Study
- Impaired Mast Cell Activation in Gene-Targeted Mice Lacking the Serum- and Glucocorticoid-Inducible Kinase SGK1
- Evaluation of {alpha}v{beta}3 Integrin-Targeted Positron Emission Tomography Tracer 18F-Galacto-RGD for Imaging of Vascular Inflammation in Atherosclerotic Mice
- Molecular Imaging of Metastatic Potential
- microPET of Tumor Integrin {alpha}v{beta}3 Expression Using 18F-Labeled PEGylated Tetrameric RGD Peptide (18F-FPRGD4)
- Noninvasive Characterization of Myocardial Molecular Interventions by Integrated Positron Emission Tomography and Computed Tomography
- Positron Emission Tomography Using [18F]Galacto-RGD Identifies the Level of Integrin {alpha}v{beta}3 Expression in Man.
- PET-Based Human Dosimetry of 18F-Galacto-RGD, a New Radiotracer for Imaging {alpha}v{beta}3 Expression
- Visualization of a primary anti-tumor immune response by positron emission tomography
- Radiolabeled RGD Peptides Move Beyond Cancer: PET Imaging of Delayed-Type Hypersensitivity Reaction