


Abstract
It remains a matter of controversy as to whether cerebral perfusion declines with healthy aging. In vivo imaging with PET permits quantitative evaluation of brain physiology; however, previous PET studies have inconsistently reported aging reductions in cerebral blood flow (CBF), oxygen metabolism, and glucose metabolism. In part, this may be because of a lack of correction for the dilution effect of age-related cerebral volume loss on PET measurements. Methods: CBF PET scans were obtained using [15O]H2O in 27 healthy individuals (age range, 19–76 y) and corrected for partial-volume effects from cerebral atrophy using an MR-based algorithm. Results: There was a significant difference (P = 0.01) in mean cortical CBF between young/midlife (age range, 19–46 y; mean ± SD, 56 ± 10 mL/100 mL/min) and elderly (age range, 60–76 y; mean ± SD, 49 ± 2.6 mL/100 mL/min) subgroups before correcting for partial-volume effects. However, this group difference resolved after partial-volume correction (young/midlife: mean ± SD, 62 ± 10 mL/100 mL/min; elderly: mean ± SD, 61 ± 4.8 mL/100 mL/min; P = 0.66). When all subjects were considered, a mild but significant inverse correlation between age and cortical CBF measurements was present in the uncorrected but not the corrected data. Conclusion: This study suggests that CBF may not decline with age in healthy individuals and that failure to correct for the dilution effect of age-related cerebral atrophy may confound interpretation of previous PET studies that have shown aging reductions in physiologic measurements.
Functional imaging techniques, such as PET have permitted in vivo quantitative evaluation of brain metabolic measurements. However, whether physiologic measures, such as resting cerebral blood flow (CBF), are altered in normal aging is still not answered definitively. Using the 133Xe inhalation method, which shows particularly poor spatial resolution relative to other methods, several investigators have demonstrated a significant reduction in mean brain CBF throughout the adult life span (1–4). PET studies of aging have variably shown reductions in CBF and oxygen metabolism in healthy persons. Leenders et al. (5) used the [15O]-labeled gas inhalation technique to demonstrate a decline of 0.5%/y in PET measures of CBF, cerebral blood volume (CBV), and cerebral metabolic rate of oxygen (CMRO2) in cortical brain regions. A similar linear reduction in mean cortical CBF was also noted by Pantano et al. (6) using PET. Other researchers have observed aging declines in CMRO2, with a milder influence of age on CBF and oxygen extraction (7–9). Still other investigators have failed to find reduced CBF values in normal aging with PET (10).
Despite significant improvements in scanner resolution over the past decade, PET studies remain substantially poorer in spatial resolution relative to anatomic imaging techniques, such as magnetic resonance (MR) imaging or CT. Because of limitations of spatial resolution, cerebral volume loss resulting from the normal aging process will cause PET measurements to be underestimated in older persons due to partial-volume averaging of brain with expanded sulcal spaces. Failure to account for this partial-volume effect can confound the interpretation of PET studies of aging (11).
Although the potential impact of partial-volume effects in aging studies using CBF and metabolism has been acknowledged (5,6,8,11–14), few investigators have attempted to correct PET data for this important source of error (15,16). In this study, we used PET imaging to examine the effect of age on regional CBF in healthy individuals. This was assessed before and after correcting the PET measurements for the effects of partial-volume averaging using an MR-based method.
MATERIALS AND METHODS
Study Subjects
The study population consisted of 27 healthy individuals recruited from the local community. The data were collected from several ongoing PET research studies at the University of Pittsburgh PET facility. Subjects ranged in age from 19 to 76 y, and there was no significant difference in age between men and women (mean age ± SD, 46.2 ± 20 y for men; 40.3 ± 20.9 y for women; 2-tailed t test, P = 0.49). These subjects were also arbitrarily subdivided into 2 groups: a young/midlife group (< 50 y; n = 18; 5 men, 13 women; mean age ± SD, 28.5 ± 6.6 y) and an elderly group (> 50 y; n = 9; 4 men, 5 women; mean age ± SD, 69.8 ± 5.4 y). Subjects were screened for past or present evidence of psychiatric or neurologic disorders and for a history of psychiatric disease in a first-degree relative. There was no significant difference in years of education (P = 0.87) between the young/midlife (mean ± SD, 16.6 ± 1.9 y) and the elderly subjects (mean ± SD, 16.4 ± 3.3 y). Elderly subjects were carefully screened for evidence of dementing illness; Mini-Mental State Examination (MMSI) (17) scores were 29.2 ± 1.1 and Mattis Dementia Rating Scale (18) values were 138.5 ± 3.0 (Mattis was not performed in 1 subject who had a MMSE score of 30/30). All subjects had no history of substance abuse, head trauma, seizures, diabetes, or other significant medical illness. Individuals using psychotropic or other medications with known central effects, including β blockers, were not studied. None of the elderly female subjects was on estrogen replacement therapy. In women of reproductive age, serum β human gonadotropin pregnancy testing was performed within 48 h before the PET study. After complete description of the study to the subjects, written informed consent was obtained before study entry in compliance with University of Pittsburgh Institutional Review Board and Radioactive Drug Research Committee procedures.
PET Imaging and Data Acquisition
PET studies were conducted with a Siemens 951R/31 PET scanner (CTI PET Systems, Knoxville, TN), which acquires images in 31 parallel planes with an interplane separation of 3.4 mm. After a 10-min transmission scan using rotating rods of 68Ge/68Ga, 50 mCi [15O]H2O was injected as an intravenous bolus using an automatic injection system (19) and dynamic emission scanning performed in 2-dimensional imaging mode (septa extended) for 3 min collected in 20 frames (ten 3-s, three 10-s, four 15-s, and three 20-s frames). The participants kept their eyes closed; their ears were unplugged; and they were in a quiet, dimly lit room. Head movement was minimized by the use of a thermoplastic mask and head-holder system. For the [15O] H2O studies, arterial blood sampling was performed using a Siemens liquid activity monitoring system that withdraws blood at a rate of 6 mL/min and detects radioactive events with dual BGO scintillation crystals. The blood was monitored for 3.5 min. PET data were corrected for radioactive decay, attenuation by transmission, and scatter and reconstructed with a Hanning-window cut off at 80% of the Nyquist frequency.
MR Imaging
MR imaging, performed on a Signa 1.5 Tesla scanner (GE Medical Systems, Milwaukee, WI) using a standard head coil, was used to guide region-of-interest (ROI) selection and for partial-volume correction of the PET data. After a scout T1-weighted sagittal sequence, a T1-weighted axial series (TE = 18, TR = 400, NEX = 1, slice thickness = 3 mm/interleaved) and coronal volumetric spoiled gradient recalled (SPGR) sequence (TE = 5, TR = 25, flip angle = 40°, NEX = 1, field of view = 24 cm, image matrix = 256 × 192 pixels) were acquired. Fast-spin echo T2 and proton density weighted images were also acquired to exclude unexpected neuropathology. The SPGR MR images were used for partial-volume correction and ROI selection. Pixels that corresponded to scalp and calvarium were removed from the SPGR MR images (20) before registration with the PET images. The MR images and [15O] H2O PET data (summed over the interval 30–120 s) were co-registered according to the method of Woods et al. (21) as further validated in our laboratory (22).
Data Analysis
ROI Selection.
ROIs were hand drawn on the co-registered MR images and transferred to the dynamic PET data to generate time–activity curves.
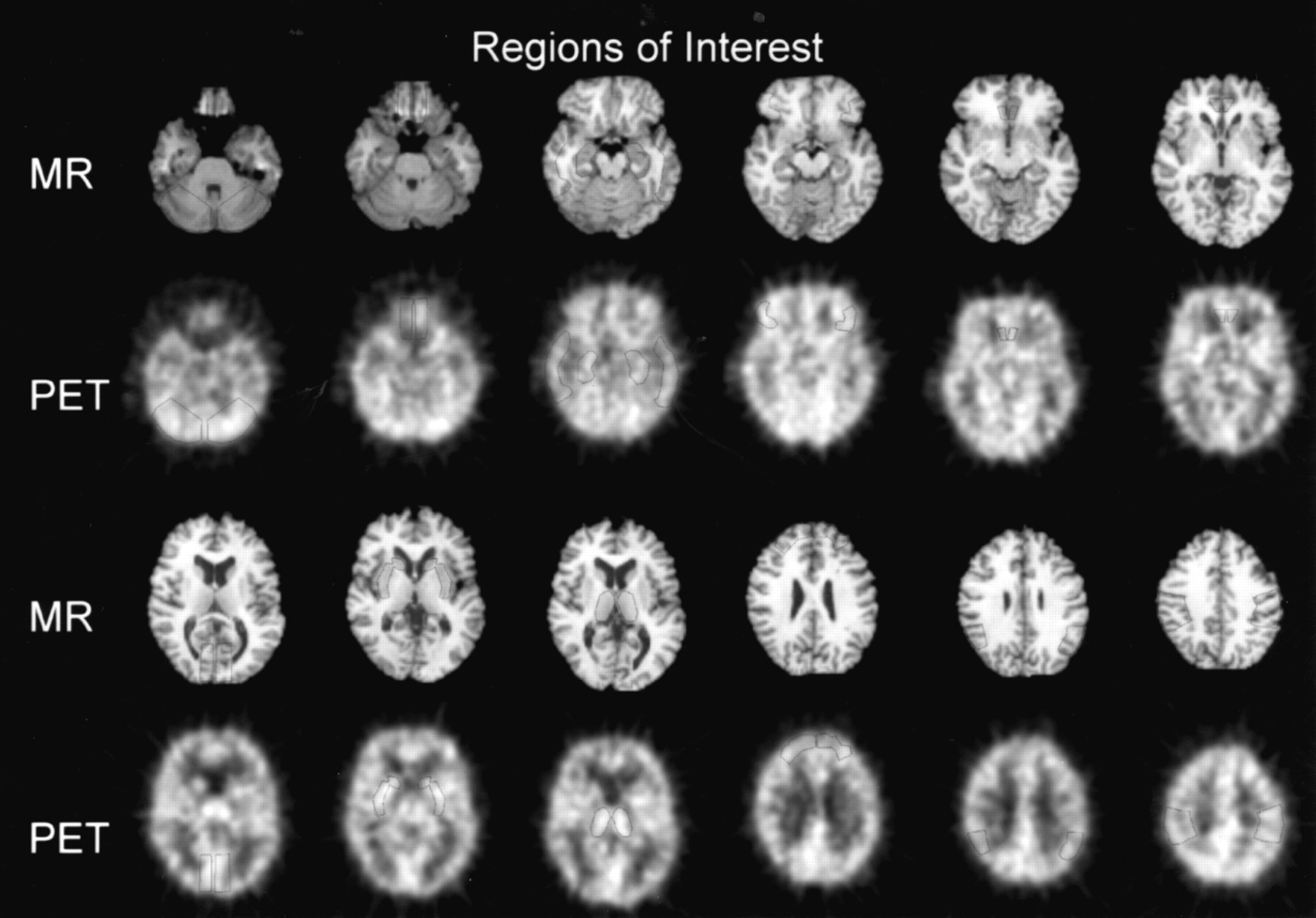
A total of 11 representative brain regions were sampled (Fig. 1). These included 8 cortical areas: anterior cingulate (drawn on 3 planes), sensory–motor cortex, parietal cortex, medial temporal cortex, lateral temporal cortex, medial orbito-frontal cortex, lateral orbito-frontal cortex, and occipital cortex. Subcortical ROIs included the thalamus and basal ganglia. The cerebellum was also sampled. For all brain regions, paired right and left ROIs were averaged to reduce noise.

Selected summed [15O]H2O PET images and co-registered SPGR MR images illustrate location of sampled ROIs.
Tracer Kinetic Modeling.
The CBF data were analyzed using a 1-tissue compartment model that accommodates for vascular volume and blood input function dispersion and timing delays (23,24). This method provides kinetic parameter estimates of blood flow (K1) and distribution volume (K1/k2). K1 was assumed to represent blood flow and was the measure of interest.
Partial-Volume Correction.
An MR-based partial-volume correction method was applied to the regional K1 values. This method has been previously validated in simulation and phantom data and applied to human metabolic and receptor PET data (15,25–27). Briefly, the registered MR images were segmented into brain and cerebrospinal fluid (CSF) pixels by selection of an optimal threshold value obtained by fitting pixel signal intensities to Gaussian distributions. A binary brain tissue image dataset was then created by assigning brain voxels a value of unity and CSF voxels a value of zero. This binary brain map was convolved using a Gaussian smoothing kernel approximating that of the PET spatial resolution as measured with point sources in water (in-plane, full-width at half-maximum [FWHM], 10.3 mm; z-axis FWHM, 5.9 mm) (27). A set of regional tissue correction factors (range, 0 to 1.0) was generated for each subject by ROI sampling of the corresponding convolved brain tissue image. Partial-volume correction of the regional CBF measurements was performed by dividing the uncorrected K1 by its tissue correction factor to generate a corrected K1 value.
In 3 subjects, the volumetric SPGR sequence was not performed. In these cases, the T1-weighted axial images were used for partial-volume correction. To ensure that the image registration and segmentation achieved with this series were equivalent to that obtained with the SPGR images, we compared the results of partial-volume correction performed independently using the SPGR images and the T1-weighted images in 3 other subjects in whom both image sequences had been obtained.
Statistical Analysis
The PET data were analyzed in 2 ways: by examining between-group comparisons of young/midlife and elderly subgroups of subjects and by pooling all subject data to examine correlations between age and CBF. Two-tailed Student's t tests were applied to the comparison of uncorrected CBF values in the young/midlife and elderly groups. This comparison was similarly examined for the corrected PET data. Using the pooled data, Pearson correlations were performed between age and both uncorrected and corrected CBF measurements for each ROI individually and for the mean of the 8 cortical ROIs. The relationship between age and the magnitude of tissue correction values was also characterized using Pearson correlations. Statistical significance was set at P < 0.05.
RESULTS
The mean difference (±SD) in the regional tissue correction factors achieved with the SPGR and T1-weighted images in the 3 subjects in which this comparison was made was 1.53% ± 1.44%. On the basis of this finding, we concluded that use of the T1-weighted images for partial-volume correction was an acceptable alternative to the volumetric SPGR series in the 3 cases when the latter was not available.
Group Analysis: Young/Midlife Versus Elderly Groups
The data demonstrated a significant difference between the young/midlife and elderly groups in the mean uncorrected CBF for all 8 cortical regions sampled (mean [±SD], 56 ± 10 mL/100 mL/min vs. 49 ± 2.6 mL/100 mL/min, respectively; P = 0.01, 2-tailed t test) (Fig. 2A). The CBF values were comparable with those previously reported (23,28). The MR-derived tissue correction factors were significantly smaller (corresponding to a greater proportion of CSF within the region) for the elderly relative to the young/midlife subjects in all brain regions (with the exception of trend-level significance in the medial orbito-frontal cortex) (Table 1). After partial-volume correction, the intergroup difference in cortical CBF was no longer significant (young/midlife mean ± SD, 62 ±10 mL/100 mL/min; elderly mean ± SD, 61 ± 4.8 mL/100 mL/min; P = 0.66) (Fig. 2B). When all regions (both cortical and subcortical) were considered, the mean uncorrected CBF was reduced in the elderly subjects with borderline significance achieved (P = 0.04) (young/midlife mean ± SD, 58 ± 10; elderly, 52 ± 2.9 mL/100 mL/min). Similarly, this age effect lost significance (P = 0.72) after partial-volume correction of the PET data (young/midlife mean ± SD, 63± 11; elderly, 62 ± 4.4 mL/100 mL/min).
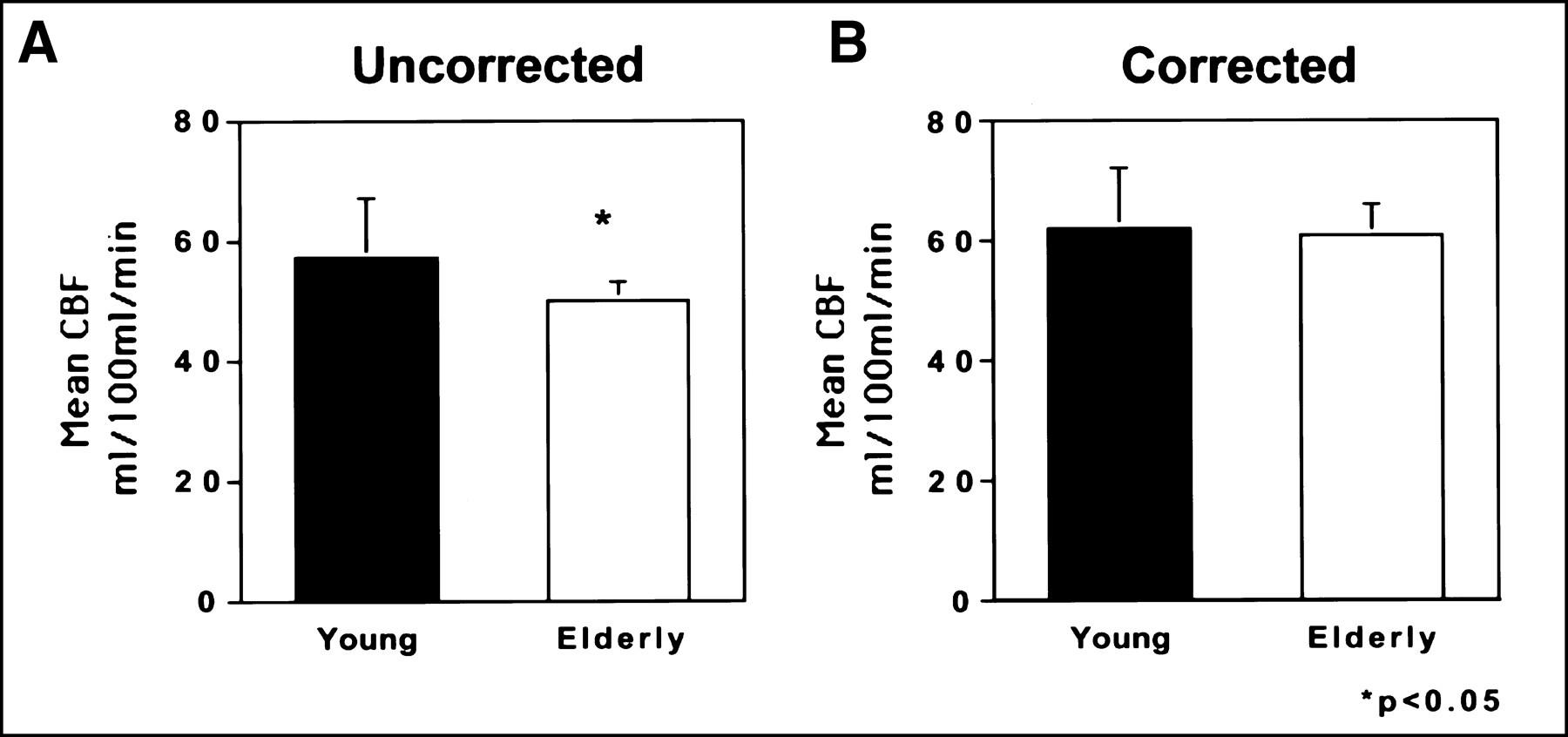
Bar graphs of mean cortical CBF (mL/100 mL/min) in elderly versus young/midlife subjects. These data demonstrate significant reduction in cortical CBF in elderly subjects for uncorrected data (A). However, after partial-volume correction of PET data, no difference in CBF is observed between groups (B).
Mean Regional Tissue Correction Factors and Pearson Correlation Coefficients for Comparison of Age Versus Tissue Correction Factor
Pooled Analysis: Age Correlations
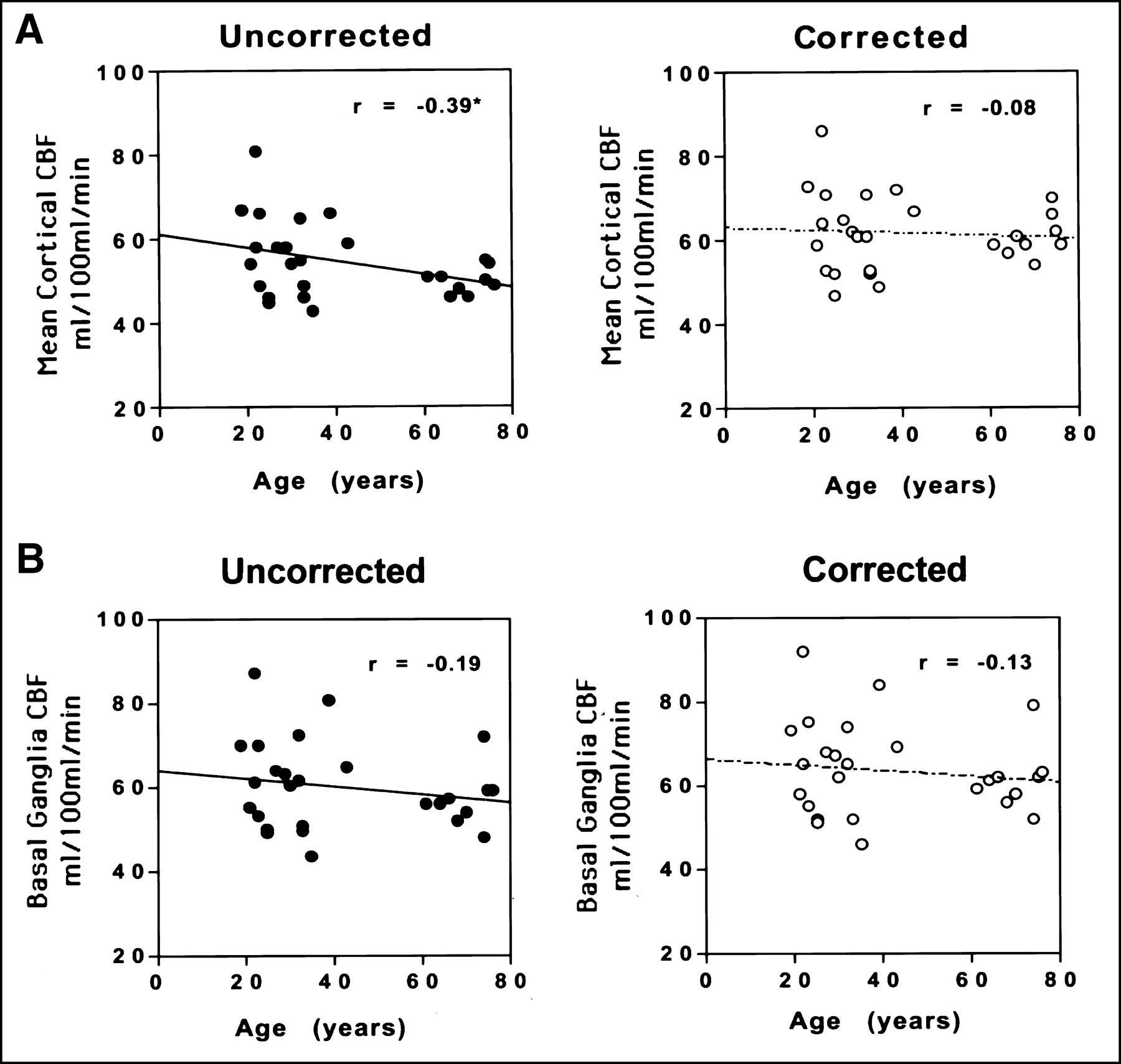
When the 27 subjects were pooled, there was a small but statistically significant inverse correlation between age and mean uncorrected cortical CBF (Pearson r = −0.39; P = 0.04) (Table 2; Figure 3A). Similarly, cortical tissue correction factors showed a strong correlation with age (Pearson r = −0.84; P = 0.0001) (Table 1; Figure 4). No significant correlation with age was observed, however, when the PET data were corrected for partial-volume effects (r = −0.075; P = 0.71). Individual regional correlations between age and CBF for the pooled data are also shown in Table 2. Several regions showed a similar pattern of a statistically significant relationship between age and CBF values before but not after partial-volume correction (lateral temporal, lateral orbito-frontal cortex; trend significance: anterior cingulate, parietal cortex). In 1 cortical region, the medial orbito-frontal cortex, a significant age–CBF inverse correlation existed in both the uncorrected and corrected data. In the basal ganglia, thalamus, and cerebellum, no significant relationship between age and CBF was present in either the uncorrected or corrected data (Fig. 3B).
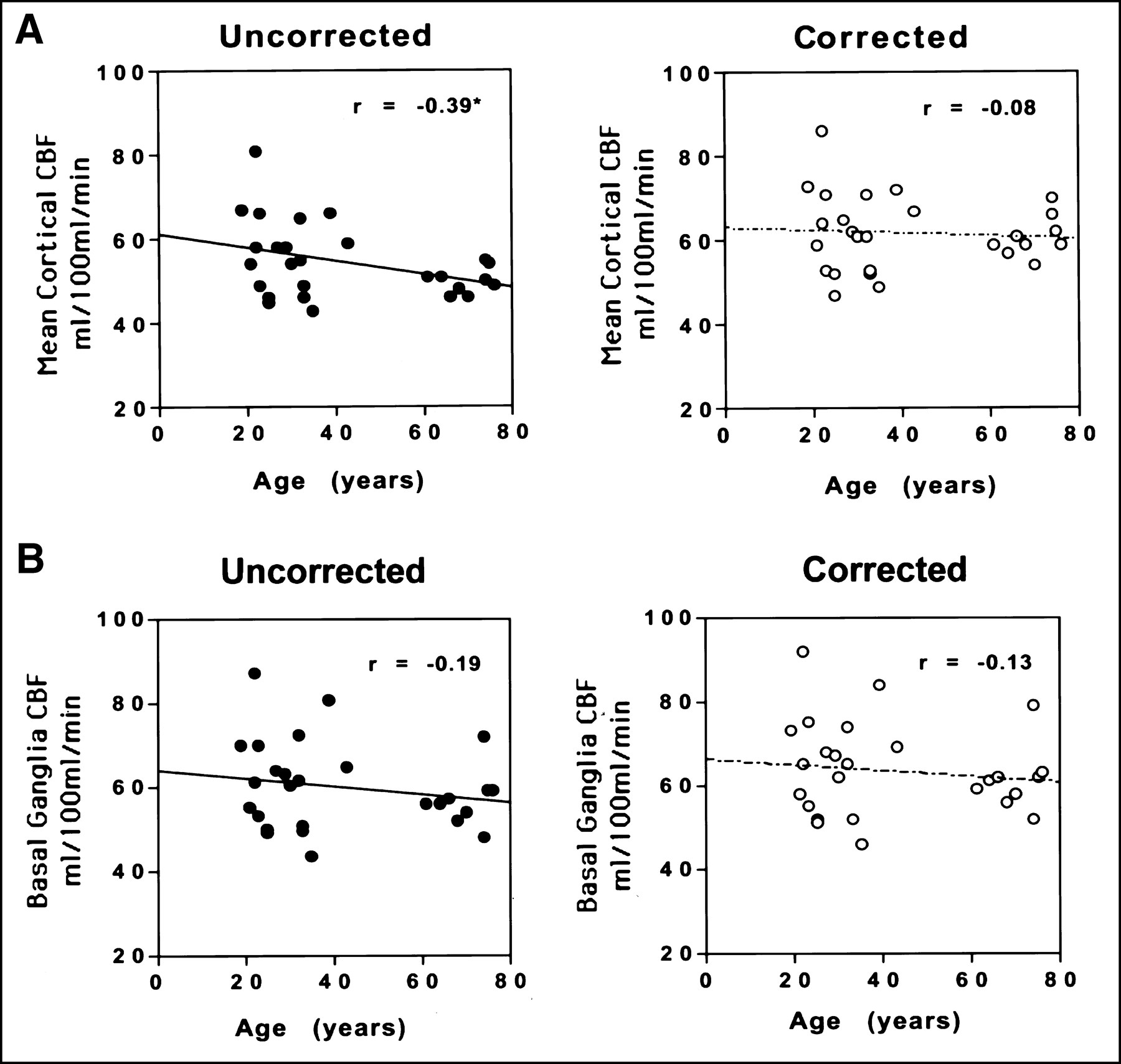
Scatter plots of age versus CBF in cortex (mean of 8 cortical ROIs) (A) and basal ganglia (B). Note the significant correlation before but not after partial-volume correction in cortical regions. In basal ganglia, however, partial-volume correction has little effect on nonsignificant correlation between age and CBF.
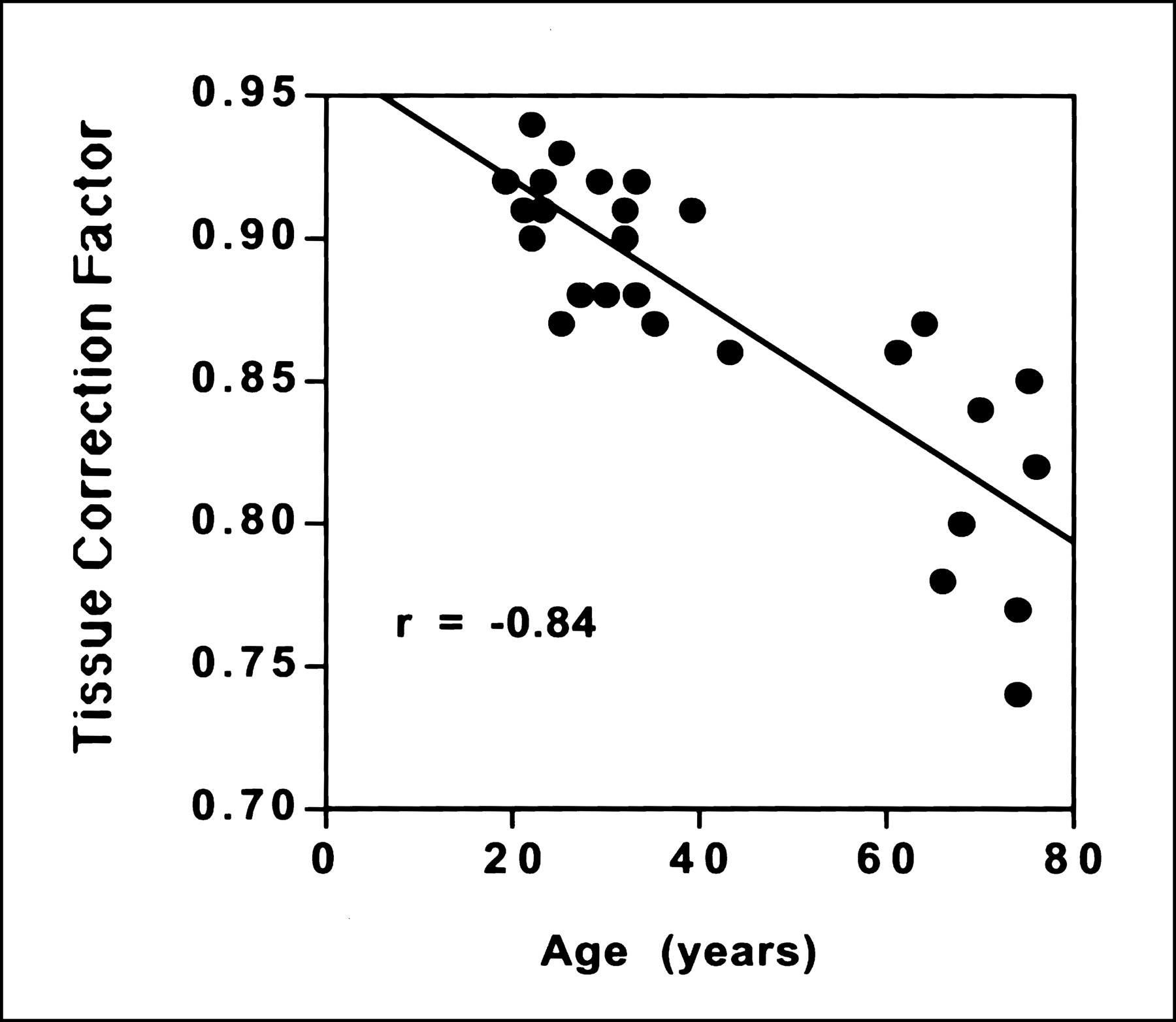
Scatter plot demonstrating inverse correlation between average cortical tissue correction factor and subject age. Tissue correction factor is a regional measure of the relative influence of CSF and brain tissue in PET image, and, therefore, lower numbers imply greater atrophy.
Regional Pearson Correlation Coefficients for Age Versus CBF Values Before and After Partial-Volume Correction
DISCUSSION
The results of this study demonstrated an apparent reduction in mean cortical CBF with healthy aging, which lost significance after correcting the PET data for partial-volume effects that were from cerebral volume differences among the subjects. The relationship between CBF and age in the uncorrected data was consistent with that reported in other PET studies of aging in which cerebral atrophy was not considered (5,13). We previously predicted, using PET simulations generated from brain MR images acquired in healthy individuals (age range, 18–79 y), that a similar effect of aging on CBF could be produced (27). In these simulations, although brain tracer concentration was held constant, partial-volume effects caused an apparent substantial inverse correlation between mean gray-matter CBF and age. After MR-based partial-volume correction was applied to the simulated PET data, the apparent age–CBF relationship no longer existed, which agrees with the findings of this study.
Previous studies have shown the largest age effects on CBF in frontal and perisylvian temporal brain areas (6,29), which is consistent with the uncorrected PET results in the current study. These same brain regions have also been shown to exhibit preferential volume loss with normal aging (30), which further supports the need to correct PET data for structural brain changes when comparing young and elderly subject groups. Although all cortical regions showed the same pattern of a reduction in the magnitude of the inverse correlation between age and CBF, this correlation was not statistically significant in the uncorrected data for several individual ROIs. This may be, in part, because of limitations of sample size. Expected differences among individuals in the pattern of healthy brain aging likely account for the larger variability in tissue correction factors that was observed in the elderly subject group. It is important that the intersubject variability in regional CBF values was comparable with previously published values (28) and did not appear to be influenced by applying partial-volume correction. It is interesting, however, that the significant inverse correlation between age and blood flow in the medial orbito-frontal cortex remained significant after partial-volume correction. This finding suggests a potential true decline in regional CBF with age in this brain region, which was also relatively less affected by atrophy (as evidenced by only a trend-level difference in tissue correction factors between the 2 age groups; Table 1). Although further studies are needed to confirm these results, these data are consistent with the observations of alterations in frontal lobe executive function and mood in healthy aging (31). It is not surprising that no relationship between CBF and age was seen for the subcortical nuclei and cerebellum. Aging effects on CBF have been reported less frequently in these brain areas, which are also relatively less susceptible to the diluting influence of age-related atrophy on regional PET measurements (5,32).
Several investigators have attempted to address the potential confound of age-related cerebral atrophy on PET measurements in studies of CBF and metabolism. Marchal et al. (7) observed no consistent relationship between mean cortical CMRO2 measures and subjective ratings of atrophy determined from CT images, and they concluded that atrophy did not explain aging reductions in metabolic measurements. Furthermore, De Leon et al. (14) observed that regional values of cerebral metabolic rate for glucose (CMRglu) showed disproportionately weaker correlation with age than did linear CT measurements of atrophy. In addition, Fazekas et al. (33), in a subjective comparison of FDG PET and CT, observed an inconsistent pattern between atrophy and hypometabolic lesions in elderly subjects. It should be noted, however, that CT measurements of cerebral atrophy are considerably less reliable than those determined from MR imaging (34). Using MR-derived measures of atrophy, Tanna et al. (11) suggested that cerebral volume loss associated with healthy aging could account for more than 15% of the variance in PET metabolic measurements. Furthermore, Yoshii et al. (35) noted a significant correlation (r = 0.24) between MR estimates of brain atrophy and whole-brain CMRglu. These investigators suggested that when brain atrophy and brain volume were considered, there was little evidence that CMRglu truly declined with healthy aging.
There is, however, evidence that aging affects the autoregulatory capacity of the cerebrovascular system to respond to vasodilatory challenge (36). Therefore, we might hypothesize that the brains of healthy older persons may exhibit unaltered baseline perfusion, but the diminished ability of small arterioles to compensate for changes in systemic perfusion pressure may increase susceptibility of the brain to ischemic damage. Consistent with a loss of flow reserve is the observation of an aging reduction in the functional MR-imaging response to visual stimulation (37) and cognitive tasks (38). Marchal et al. (7), however, found a preserved cerebrovascular reserve capacity, as evidenced from an unchanged CBF:CBV ratio in healthy aging.
Some of the variability in findings among studies of the effects of aging on cerebrovascular physiology may result from subject-selection considerations and the lack of a unified definition of “healthy” or “successful” aging. Indeed, CBF and other metabolic parameters may be most prone to deterioration with age in individuals with elevated blood pressure or other cerebrovascular risk factors (1). Martin et al. (13) used statistical parametric mapping to demonstrate an age-related decrease in relative regional CBF in limbic and association areas and suggested that regionally specific functional losses may be linked to aging changes in cognition. However, in very healthy elderly individuals, alterations in cognition have been less consistently reported (39). The nature of the interplay among the variables of age, cerebrovascular risk measures, and cognition, and their relationship with CBF, may be addressed in future, larger studies.
Although gender differences in cerebral perfusion have been previously reported (2,40), the predominance of women in this study and a limited total sample size made it difficult to examine the data for gender effects. However, the similarity of the age distributions of men and women in this study would tend to limit any potential bias introduced by gender. Furthermore, there is considerable evidence for differential patterns of regional cerebral volume loss with aging in men and women (41,42), which may have influenced prior PET and SPECT reports of gender effects on CBF or metabolism. Partial-volume correction performed on the PET data in this study would have eliminated the influence of sex differences in brain morphology on the CBF measurements.
CONCLUSION
In neurodegenerative disorders, it has been demonstrated that deficits in PET measurements of energy metabolism are not fully accounted for by regional cerebral atrophy (15,33). However, the findings of this study suggest that, in the absence of significant medical problems, resting cortical perfusion may be unaltered by healthy aging. Furthermore, this study illustrates that failure to correct PET data for partial-volume effects when comparing subjects across a large age range may be misleading.
Acknowledgments
The authors wish to thank Marsha Dachille, Donna Milko, James Rusckiewicz, Denise Ratica, David Manthei, and Daniel Holt for their assistance in conducting the PET studies. This work was supported by Public Health Service grants MH52247 (Mental Health Clinical Research Center for Late-Life Mood Disorders), AG05133 (Alzheimer's Disease Research Center), MH49936, MH57078, MH01621, MH01210, and MH00295; and the Radiological Society of North America Research and Education Fund.
Footnotes
Received Jul. 21, 1999; revision accepted Oct. 27, 1999.
For correspondence or reprints contact: Carolyn Cidis Meltzer, MD, University of Pittsburgh Medical Center, PET Facility, B-938, 200 Lothrop St., Pittsburgh, PA 15213-2582.
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Microvascular pulsatility of the ageing brain and confounding effects of anaesthesia
- Interspecific scaling of blood flow rates and arterial sizes in mammals
- Accuracy of Parenchymal Cerebral Blood Flow Measurements Using Pseudocontinuous Arterial Spin-Labeling in Healthy Volunteers
- Perfusion of Ischemic Brain in Young and Aged Animals: A Laser Speckle Flowmetry Study
- Age-Related Metabolic Profiles in Cognitively Healthy Elders: Results from a Voxel-Based [18F]Fluorodeoxyglucose-Positron-Emission Tomography Study with Partial Volume Effects Correction
- Initial Experience in Using Continuous Arterial Spin-Labeled MR Imaging for Early Detection of Alzheimer Disease
- Comparison of 18F-FDG and PiB PET in Cognitive Impairment
- MRI-Based Correction for Partial-Volume Effect Improves Detectability of Intractable Epileptogenic Foci on 123I-Iomazenil Brain SPECT Images
- Brain Imaging in Patients With Diabetes: A systematic review.
- Effects of antidepressant treatment with rTMS and fluoxetine on brain perfusion in PD
- Lack of Effect of Pravastatin on Cerebral Blood Flow or Parenchymal Volume Loss in Elderly at Risk for Vascular Disease
- 5-HT1A Receptors Are Reduced in Temporal Lobe Epilepsy After Partial-Volume Correction
- Brain Incorporation of 11C-Arachidonic Acid, Blood Volume, and Blood Flow in Healthy Aging: A Study With Partial-Volume Correction
- White matter lesions impair frontal lobe function regardless of their location
- Application of MRI-Based Partial-Volume Correction to the Analysis of PET Images of {micro}-Opioid Receptors Using Statistical Parametric Mapping
- Correction for Partial-Volume Effects on Brain Perfusion SPECT in Healthy Men
- Importance of Partial-Volume Correction in Brain PET Studies