


Abstract
Preclinical studies suggest that serotonin 1A receptors (5-HT1A) play a role in temporal lobe epilepsy (TLE). Previous PET studies reported decreased 5-HT1A binding ipsilateral to epileptic foci but did not correct for the partial-volume effect (PVE) due to structural atrophy. Methods: We used PET with 18F-trans-4-fluoro-N-2-[4-(2-methoxyphenyl)piperazin-1-yl]ethyl-N-(2-pyridyl)cyclohexanecarboxamide (18F-FCWAY), a 5-HT1A receptor antagonist, to study 22 patients with TLE and 10 control subjects. In patients, 18F-FDG scans also were performed. An automated MR-based partial-volume correction (PVC) algorithm was applied. Psychiatric symptoms were assessed with the Beck Depression Inventory Scale. Results: Before PVC, significant (uncorrected P < 0.05) reductions of 18F-FCWAY binding potential (BP) were detected in both mesial and lateral temporal structures, mainly ipsilateral to the seizure focus, in the insula, and in the raphe. Group differences were maximal in ipsilateral mesial temporal regions (corrected P < 0.05). After PVC, differences in mesial, but not lateral, temporal structures and in the insula remained highly significant (corrected P < 0.05). Significant (uncorrected P < 0.05) BP reductions were also detected in TLE patients with normal MR images (n = 6), in mesial temporal structures. After PVC, asymmetries in BP remained significantly greater than for glucose metabolism in hippocampus and parahippocampus. There was a significant inverse relation between the Beck Depression score and the ipsilateral hippocampal BP, both before and after PVC. Conclusion: Our study shows that in TLE patients, reductions of 5-HT1A receptor binding in mesial temporal structures and insula are still significant after PVC. In contrast, partial-volume effects may be an important contributor to 5-HT1A receptor-binding reductions in lateral temporal lobe. Reduction of 5-HT1A receptors in the ipsilateral hippocampus may contribute to depressive symptoms in TLE patients.
Experimental studies support a role for serotonin (5-hydroxytryptamine, 5-HT) neurotransmission in epilepsy. 5-HT1A receptor activation shows antiseizure effects in partial seizure models, blocked by the highly selective 5-HT1A antagonist WAY100635 (1,2).
In an initial PET study with 18F-trans-4-fluoro-N-2-[4-(2-methoxyphenyl)piperazin-1-yl-ethyl]-N-(2-pyridyl)cyclohexanecarboxamide (18F-FCWAY), a WAY100,635 analog (3,4), we reported decreased temporal 5-HT1A binding ipsilateral to seizure foci in patients with temporal lobe epilepsy (TLE) (5). Subsequent studies using 5-HT1A receptor antagonists carbonyl-11C-WAY-100,635 (11C-WAY) and 4,2-(methoxyphenyl)-1-[2-(N-2-pyridinyl)-p-fluorobenzamido]ethylpiperazine (18F-MPPF) reported reduced 5-HT1A receptor binding in insula and anterior cingulate as well as temporal lobe (6–8).
However, many patients in these studies had mesial temporal sclerosis (MTS) and hippocampal atrophy on MR images. Moreover, patients with intractable TLE frequently have extrahippocampal atrophy (9). Owing to the partial-volume effect (PVE), brain atrophy could have affected PET findings. Thus, physiologic significance was uncertain.
To properly address this issue, MR-based partial-volume correction (PVC) is necessary (10–12). PVC algorithms were used in TLE studies with 11C-flumazenil (13), and limited to mesial temporal regions, 18F-FDG (14), and the muscarinic antagonist 4-123I-iododexetimide (15), but not 5-HT1A ligands.
We implemented (16) an MR-based PVC algorithm that, for each frame, corrects gray matter activity for spill-out of gray as well as spill-in of white matter activity into gray matter pixels (11,12). This algorithm, appropriate for clinical investigations automatically estimated white matter activity and did not increase data variability.
In the present study we evaluated 18F-FCWAY binding before and after PVC in temporal as well as extratemporal regions in a larger patient group (5). Patients also underwent PET with 18F-FDG; PVC was applied to parametric images of the cerebral metabolic rate for glucose (CMRglu) as well as 18F-FCWAY binding.
MATERIALS AND METHODS
Subjects
We studied 22 patients (18 men; mean age ± SD, 37 ± 11 y) referred to the Clinical Epilepsy Section, National Institute of Neurologic Disorders and Stroke (NINDS), National Institutes of Health (NIH), for evaluation of medically refractory TLE (5). Ictal video-electroencephalography (EEG) confirmed each patient’s epileptogenic zone localization. Five had subdural recording after PET for language mapping. Epilepsy onset age was 15 ± 10 y (range, 1–34 y); disease duration was 23 ± 13 y (range, 5–46 y). Patients with structural lesions (except MTS), progressive neurologic disorders, or taking any medications aside from antiepileptic drugs (AEDs), were excluded. Eleven were included in our earlier study (5). None experienced seizures for at least 2 d, or a secondarily generalized tonic clonic seizure for at least 1 mo, before PET studies. Depressive symptoms were assessed with the Beck Depression Inventory (BDI) (17). The study was approved by the NINDS Investigational Review Board and Radiation Safety Committee. We studied 10 healthy volunteers (7 men; mean age ± SD, 35 ± 9 y), screened with general physical examination and routine laboratory tests.
MR Procedure
All subjects underwent MRI using a 1.5-T Horizon (GE Healthcare). In patients, T1-weighted spoiled gradient images (repetition time [TR], 27 ms; minimum echo time [TE]; flip angle, 20°) were acquired in coronal orientation. In healthy subjects, 3-dimensional (3D) IRprepped (variable TR; minimum TE; and inversion time, 300 ms) were acquired in the transverse plane. Coronal and transverse MR images had the same spatial resolution (voxel size, 0.94 × 0.94 × 1.5 mm; 256 × 256 × 124 slices). Transverse slices were resliced in coronal planes for analyses. Standard coronal T2 and FLAIR (Fluid Attenuated Inversion Recovery) images were obtained. All MR images were read by experienced neuroradiologists who were unaware of PET results.
Radiochemistry
18F-FCWAY was synthesized as described (3); specific activity at injection was >18.5 TBq/mmol (>500 Ci/mmol).
PET Procedure
We used an Advance Tomograph (GE Healthcare; full width at half maximum [FWHM], 6–7 mm), scanning 35 simultaneous slices with 4.25-mm slice separation. During scanning, in a quiet, dim room with eyes closed and ears unoccluded, a thermoplastic facemask held the subject’s head in place (5). Continuous EEG monitoring was performed to ensure interictal studies. A transmission scan was performed for attenuation correction. Subsequently, a 333 ± 74 MBq (9 ± 2 mCi) 18F-FCWAY bolus was injected over 60 s and dynamic frames (1–5 min) were acquired in 3D mode for 120 min.
18F-FCWAY Input Function
About 30 radial arterial samples were taken to quantify 18F-FCWAY concentration and selected samples used to measure the 18F-fluorocyclohexanecarboxylic acid metabolite (18F-FC) (18). Unbound 18F-FCWAY fraction plasma protein was measured with ultracentrifugation (5).
Data Analysis
For each subject, 18F-FCWAY radioactivity frames were registered to MR images using a mutual information cost function (16). Tissue activity frames were corrected for brain acid metabolite 18F-FC uptake, vascular radioactivity, and 18F-fluoride metabolite spillover (18). Pixel data were fitted to a 2-tissue compartment model with 3 parameters using a metabolite-corrected input function to provide parametric images of V (4). A BP measurement was derived using cerebellum as a region devoid of 5-HT1A receptors (19) and calculated as:
 Eq. 1 where VROI and VCEREB are distribution volumes in the target region of interest (ROI) and cerebellum, respectively, and f1 is the 18F-FCWAY plasma free fraction. Asymmetry indices (AIs) were computed as 200·(I –C)/(I + C), where I and C represent PET values in regions ipsilateral and contralateral to the seizure focus, respectively. In control subjects, the right side was arbitrarily called ipsilateral.
Eq. 1 where VROI and VCEREB are distribution volumes in the target region of interest (ROI) and cerebellum, respectively, and f1 is the 18F-FCWAY plasma free fraction. Asymmetry indices (AIs) were computed as 200·(I –C)/(I + C), where I and C represent PET values in regions ipsilateral and contralateral to the seizure focus, respectively. In control subjects, the right side was arbitrarily called ipsilateral.
Measurement of CMRglu
Patients underwent 18F-FDG PET for CMRglu measurement within 2 d of 18F-FCWAY scans, using a noninvasive approach (5). Generated CMRglu parametric images were registered to MR images. Dosimetric restrictions precluded 18F-FDG scans in control subjects.
PVC Algorithm
The algorithm (20) provides voxel-by-voxel binary masks of gray matter, white matter, and cerebrospinal fluid (CSF). Gray matter pixels are corrected for spill-out of gray matter activity as well as for spill-in of activity from white matter. CSF activity is set to zero. Brain white matter activity is assumed to be uniform (11). Visual inspection revealed poor segmentation in basal ganglia and thalamus, where a large fraction of gray pixels were misidentified as white matter, due probably to close white-gray intermixing (16). Since 5-HT1A receptor concentration in striatum and thalamus is negligible, and a 10% error in the estimation of white matter value would introduce only an ∼1% error in gray matter PVC data (11,12,16), potential bias of 18F-FCWAY binding in cortical gray regions is expected to be negligible. Subsequently, the binary masks are smoothed to the PET resolution by convolving the binary mask with a 6-mm FWHM 3D gaussian kernel (based on the reconstructed image resolution); these masks are named as sGM, sWM, and sCSF. The corrected activity gray matter values can be calculated as follows:
 Eq. 2 where CPVC represents the corrected activity value in a gray matter pixel after PVC, CORIG is the original uncorrected pixel value, CWM is the estimated white matter activity (16), and sGM and sWM are the pixel values from the smoothed masks for gray and white matter, respectively.
Eq. 2 where CPVC represents the corrected activity value in a gray matter pixel after PVC, CORIG is the original uncorrected pixel value, CWM is the estimated white matter activity (16), and sGM and sWM are the pixel values from the smoothed masks for gray and white matter, respectively.
ROI Analysis
3D hippocampal ROIs were manually drawn on MR volumes using an anatomic atlas (21). Large regions ensured adequate gray matter sampling and increased signal-to-noise ratio (16). To minimize the risk of potential spillover of skull 18F-fluoride activity, cerebellar ROIs were initially drawn along the outer cortical edge and then uniformly shrunk by about 50%. Due to finer sampling of MR than PET data, ROIs were placed on 6 or 7 consecutive slices for amygdala, every second or third slice for hippocampus, fusiform, and parahippocampus gyrus, and every third or fourth slice for remaining cortical regions and cerebellum. A similar strategy for ROI sampling had been previously adopted in a PVC study by our group (16).
Raphe boundaries are best defined on PET transverse planes. Thus, PET images coregistered to (transverse and coronal) MR images were resampled to a common spatial resolution (0.94 × 0.94 × 0.94 mm) and subsequently resliced to transverse planes. Circular ROIs (about 6-mm diameter) were placed sequentially on the 13 coregistered PET slices that best visualized raphe activity. Each ROI was centered on the pixels with the highest activity. Small ROIs were chosen to minimize sensitivity to PVE.
ROI measurements were computed using only gray matter pixels, as defined by the gray matter segment (16). For cerebellum and raphe, where gray matter segment definition is not accurate, the mean ROI value was computed from unmasked PET images coregistered to MR volumes. Global gray matter values were calculated as cortical average values. PET measurements were not performed for thalamus and basal ganglia, where segmentation was poor. Statistical analysis was performed using t tests, and statistical significance set at P < 0.05.
RESULTS
Sixteen patients had increased hippocampal signal or mesial temporal atrophy on MR images, whereas 6 had normal MR images. Fifteen had right and 7 had left temporal ictal EEG foci. Fourteen patients had surgery; 10 are seizure-free after 1 y. The mean BDI score was 9 ± 8 (range, 0–31); 5 of 23 patients had a score of >12, the cutoff for depression (17).
There were no significant differences in 18F-FCWAY unmetabolized or acid metabolite fraction between control subjects and patients. In controls, the 18F-FCWAY fraction was 0.10 ± 0.03, 0.034 ± 0.009, and 0.021 ± 0.007, whereas the 18F-FC fraction was 0.17 ± 0.04, 0.074 ± 0.020, and 0.027 ± 0.006 at 15, 60, and 120 min, respectively. The distribution volume in cerebellum (VCEREB) was significantly greater in patients than that in controls (0.93 ± 0.30 vs. 0.67 ± 0.32, respectively; P < 0.05). The 18F-FCWAY plasma free fraction (f1) was significantly higher in patients than that in controls (0.121 ± 0.033 vs. 0.066 ± 0.024, respectively; P < 0.001). Cerebellar V/f1 was significantly lower in patients than that in controls (7.9 ± 2.0 vs. 11.1 ± 5.3, respectively; P < 0.05). These results led us to use the form of the BP equation (Eq. 1) that does not divide by distribution volume in cerebellum and corrects for f1.
Figure 1 shows coronal T2-weighted (A, top) and T1-weighed (A, bottom) MR images of a TLE patient with a right mesial temporal focus and coregistered 18F-FCWAY V/f1 (B) and CMRglu (C) images before (top) and after (bottom) PVC. Images are at the level of the posterior hippocampus, anterior to the pons. The T1- and T2-weighed images show hippocampal atrophy and dilated right ventral temporal horn, typical of MTS. Before PVC, there is clear 18F-FCWAY BP ipsilateral mesial and lateral temporal reduction, but less CMRglu asymmetry. PVC clearly increased values in both images. However, there is no change in the pattern of asymmetric 18F-FCWAY binding reduction.

Coronal T2-weighted (A, top) and T1-weighed (A, bottom) MR images of patient with TLE with a right mesial temporal focus and coregistered 18F-FCWAY distribution volume (V/f1, B) and CMRglu (C) images before (top) and after (bottom) PVC. Coronal images are at level of posterior hippocampus, anterior to pons. V/f1 and CMRglu images are scaled to a maximum of 100 mL/mL and 10 mg/min/100 g, respectively.
In healthy subjects, BP values were highest in mesial temporal structures, intermediate in frontal, lateral temporal and parietal cortex, and lowest in occipital lobe (Table 1), consistent with human 5-HT1A receptor distribution (19). In patients, pre-PVC BP was significantly (P < 0.05) lower than that in control subjects in all mesial temporal regions, in ipsilateral temporal regions, and in contralateral middle temporal cortex. Reduced 18F-FCWAY binding was also detected bilaterally in insula and raphe. No significant (P < 0.05) group differences were detected in frontal, parietal, or occipital lobes. When patients with normal MR images (n = 6) were compared with controls, significant BP reductions were detected bilaterally in parahippocampus, hippocampus, amygdala, insula, and raphe. There was no difference in 18F-FCWAY BP between patients with and without MTS in any region.
BP Values (mL/mL) in Control Subjects and Patients with TLE Before and After PVC
After PVC, patient-control differences remained significant in most mesial temporal regions (Table 1); in ipsilateral fusiform gyrus, parahippocampus, and hippocampus and bilaterally in the insula, group differences had a statistical significance that survived correction for multiple ROI comparisons. However, group differences were no longer significant in any lateral temporal region. No significant group differences were introduced by PVC in the frontal, parietal, and occipital cortex.
Before PVC, 18F-FCWAY BP AIs (Table 2) were significantly (P < 0.05) greater in patients than those in control subjects in inferior temporal, fusiform gyrus, parahippocampus, hippocampus, amygdala, and insula. After PVC, statistical significance was lost only in the insula. Pre-PVC patient AIs were significantly greater for BP than for CMRglu in parahippocampus, hippocampus, and amygdala. After PVC, group differences remained significant in parahippocampus and hippocampus.
AIs of 18F-FCWAY BP and CMRglu Before and After PVC
Before PVC, global gray matter BP was significantly (P < 0.05) lower in patients than that in control subjects (38 ± 11 and 54 ± 14 mL/mL, respectively). After PVC, group BP differences (72 ± 21 vs. 91 ± 22 mL/mL, respectively) remained significant.
In patients, global gray matter CMRglu values were 6.9 ± 1.0 mg/min/100 g before and 11.5 ± 1.9 mg/min/100 g after PVC. The estimated CMRglu white matter value was 2.39 ± 0.47 mg/min/100 g. Asymmetries between regions ipsilateral and contralateral to the epileptic focus (Table 2) were consistent with previous reports (22,23).
There was greater interregion variability for BP than for CMRglu before and after PVC. The mean percent coefficient of variation (%CV) for BP across ROIs (excluding raphe) was quite similar for control subjects and patients before and after PVC (controls: 28% ± 2% and 26% ± 2%, respectively; patients: 34% ± 3% and 33% ± 3%, respectively). Thus, PVC did not increase 18F-FCWAY data variability. In patients, mean regional %CV for CMRglu was significantly higher (P < 0.001) after PVC (20% ± 2% and 17% ± 1%, respectively).
18F-FCWAY BP and CMRglu increases after PVC varied locally, ranging in patients from about 37% to 113% for BP and 26% to 107% for CMRglu, respectively (Fig. 2). For BP, the percentage increase was lowest in amygdala, insula, and hippocampus and highest in occipital and parietal lobes. In patients, the percentage changes were significantly (P < 0.05) higher than those in healthy subjects bilaterally in hippocampus, frontal, parietal, and occipital lobes. The percentage increases in BP were significantly (P < 0.05 after correction for multiple ROI comparisons) greater than those in CMRglu for every region, except for occipital lobes. This finding is most likely related to the lower ratio of gray matter to white matter for 18F-FCWAY than that for 18F-FDG.

Relationship between percentage increase in 18F-FCWAY distribution volume (V/f1, mL/mL) and CMRglu (mg/min/100 g) in TLE patients. Each point represents mean ROI value. Regression equation was V = 21.5 + 0.90 CMRglu; r2 = 0.95; P < 0.001. Dashed line represents identity line.
No significant relationship was found between CMRglu or BP values and either age of seizure onset or duration of epilepsy. A significant relationship (Fig. 3) was found between the BDI and 18F-FCWAY BP but not CMRglu in ipsilateral hippocampus (y = 47.6 –1.24x; r2 = 0.35, P = 0.004), which persisted after PVC (y = 81.2 –1.97x; r2 = 0.26, P = 0.016). No significant relationships were found in other regions for both 18F-FCWAY BP and CMRglu, before or after PVC, and BDI.
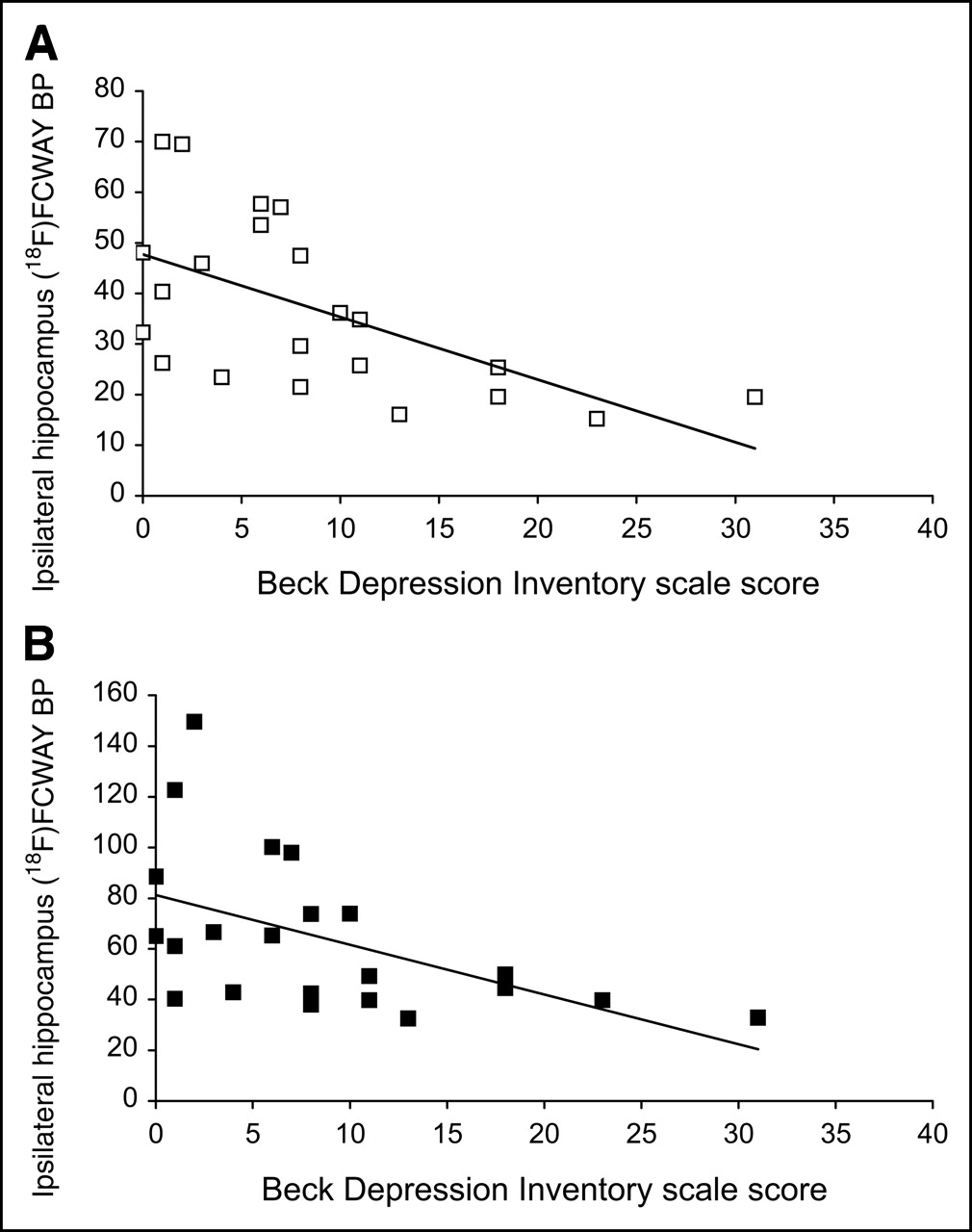
Relationship between 18F-FCWAY BP (mL/mL) in hippocampus ipsilateral to seizure focus and BDI scale score in TLE patients before (A) and after (B) PVC. Regression equations were y = 47.6 –1.24x; r2 = 0.35, P = 0.004 (A) and y = 81.2 –1.97x; r2 = 0.26, P = 0.016 (B).
DISCUSSION
Our study shows that reduced 5-HT1A 18F-FCWAY BP in mesial and insular temporal structures ipsilateral and, to a lesser extent, contralateral to temporal lobe epileptic foci is not an artifact related to PVE. Mesial temporal reductions remained significant after PVC (and correction for multiple comparisons). PVE contribution was marginal in these regions compared with the decrease in receptor binding. In contrast, reduced 18F-FCWAY BP in lateral temporal lobe did not remain significant after PVC. No group differences were detected in frontal, parietal, and occipital lobe before or after PVC.
The finding of significant contralateral BP reductions is not surprising, since radiologic and histologic evidence for bilateral hippocampal pathology, and hypometabolism on PET 18F-FDG studies, can be present in 30%–50% of patients with intractable TLE (24).
Previous 5-HT1A PET studies reported reduced receptor concentrations in some lateral temporal and frontal areas (6–8). However, in patients with TLE, atrophy is not limited to mesial structures but involves lateral temporal and frontoparietal cortex as well (25). In our study, these regions showed no significant differences from control in 18F-FCWAY BP after PVC, indicating that cortical atrophy was an important contributor to reduced uncorrected values. Thus, PVE, in addition to propagation of ictal activity, might be an additional explanation for reduced 5-HT1A binding reported in these regions (6).
For both 18F-FCWAY BP and CMRglu, there was little effect of PVC on the AIs (Table 2). This finding is likely related to the fact that bilateral atrophy minimizes the magnitude of PVE. Uncorrected BP AIs were significantly larger than corresponding CMRglu values in hippocampus, parahippocampus, and amygdala; after PVC, statistical significance was lost only in amygdala.
PVC Algorithm
MR-based algorithms for PVC have been used successfully in PET studies of healthy aging and Alzheimer’s disease (10,11,16,26,27). In a previous study with 11C-arachidonic acid, no increase in variability after PVC was found (16), whereas a significant increase in variability was detected for CMRglu in the current study. For PET data with large intersubject variability, such as FCWAY BP and arachidonic acid incorporation rate (16), the additional noise of PVC is undetectable. For CMRglu, with lower intersubject variability, a moderate increase in variability owing to PVC is detectable.
Choice of Binding Index
Uncorrected VCEREB was significantly greater in patients than that in control subjects. Cerebellum is assumed to be devoid of 5-HT1A receptors (19) and, thus, to provide a reliable estimate of free plus nonspecific binding (5–8). Since VCEREB is proportional to f1/f2, where f2 represents 18F-FCWAY brain free fraction (28), this finding might be related to a combination of changes in f1 and f2. Subtraction of higher VCEREB for regional distribution volumes (VROI, Eq. 1) might, in principle, have overestimated group differences, most likely in low binding regions—that is, occipital and parietal lobes. However, no such differences were found before or after PVC. This issue might be more relevant if a reference tissue model were adopted (6,7) and a different BP derivation used. In this case, the BP (28), is equal to:
 Eq. 3 Thus, the denominator, not just the numerator (Eq. 1), of the ratio would be affected. When this model is used, the absence of significant group differences in cerebellum binding is assumed but cannot be verified (29,30).
Eq. 3 Thus, the denominator, not just the numerator (Eq. 1), of the ratio would be affected. When this model is used, the absence of significant group differences in cerebellum binding is assumed but cannot be verified (29,30).
The higher value of 18F-FCWAY free fraction (f1) in patients than that in control subjects (0.121 ± 0.033 vs. 0.066 ± 0.024, respectively; P < 0.001) is most likely due to AED displacement of 18F-FCWAY from protein-binding sites (31). Almost all patients were taking at least one highly protein-bound AED, such as carbamazepine, phenytoin, or valproic acid, which could have led to this effect. Correction for f1 eliminates the bias in distribution volume measurements that would be introduced by this difference.
Increases of 18F-FCWAY BP and CMRglu after PVC were lowest in amygdala, insula, and hippocampus and highest in occipital and parietal lobes. This finding appears to contrast with the difference between hippocampal and occipital involvement in TLE. However, regional differences in PVC effects are influenced by local geometry. Cortical gray matter pixels in thin gyri surrounded in 3 dimensions by several white matter pixels will have small sGM and high sWM values (Eq. 2). The opposite will occur in regions, such as hippocampus and amygdala, that are relatively homogeneous gray matter (16). This explains the strong linear relationship in Figure 2. The offset from the line of identity is due to a larger white matter spill-in for 18F-FDG than for 18F-FCWAY, since white matter BP is very small.
Biologic Interpretation
The BP used in this study (Eq. 1) is equivalent to Bmax/Kd, where Bmax (nmol/L/mL) is the maximum receptor concentration available for ligand binding, and Kd (nmol/L) is the ligand-receptor dissociation constant at equilibrium (28). Reduced BP may be due to decreased receptor or increased 5-HT concentration (decrease in Bmax), decreased ligand receptor affinity (increased Kd), or a combination. Estimation of Bmax and Kd would require at least 2 PET scans with different 18F-FCWAY specific activities. This is impractical in clinical studies. Moreover, 18F-FCWAY BP is not likely to be flow sensitive (32).
Previous studies in TLE patients reported greater decreases in 11C-flumazenil binding than neuronal loss in mesial temporal regions ipsilateral to seizure foci (13). Autoradiography performed with 3H-flumazenil on surgical specimens showed reduced Bmax and increased Kd (13). Preliminary results from an autoradiographic study on temporal lobectomy specimens from a subgroup of our patients indicated a decrease in Bmax, with no changes in Kd in the hippocampus, whereas neuronal counts were diminished to a lesser extent (33).
Since some AEDs may increase 5-HT concentration (5), direct competition of the ligand with an increased 5-HT concentration cannot be excluded. However, for the serotonergic system, there is little evidence of sensitivity of high- affinity tracers, such as 18F-FCWAY, to changes in 5-HT concentration (34). Moreover, a preliminary analysis performed on PET data did not show significant AED effects on mesial and lateral temporal or raphe 18F-FCWAY BP values (35).
Our study, along with the preliminary autoradiographic results (33), suggests that receptor loss may be part of the initial phase of neuronal dysfunction in TLE, followed by hypometabolism and eventual structural atrophy. Since patients with new-onset epilepsy may have a low incidence of hypometabolism (36), 18F-FCWAY PET might be particularly useful for early detection of functional abnormalities in TLE patients. Early 5-HT1A receptor loss might lower the seizure threshold (1,37). Results with both 11C-flumazenil and 18F-FCWAY suggest that receptor-binding studies may be of clinical value for patients with uncontrolled epilepsy being considered for surgery, particularly when other imaging studies are equivocal (5,13). Studies with ligands such as 5-HT2A receptors or the serotonin transporter, not yet performed in epilepsy patients, might be of interest as well.
The physiologic events underlying 5-HT1A binding reduction in raphe remain elusive. In contrast to cortex, 5-HT1A raphe receptors are somatodendritic. After rapid increases in extracellular 5-HT, these receptors inhibit serotonergic neuronal firing rate and, thus, 5-HT release in cortical areas (38). Desensitization of 5-HT1A autoreceptors occurs after 3–4 wk of antidepressant therapy, and a similar mechanism cannot be excluded in TLE patients (38).
Epilepsy and Depression
Depression is found frequently in TLE patients. In a previous study, a significant inverse correlation between the Montgomery Åsberg Depression Rating scale and ipsilateral anterior cingulate 5-HT1A binding was reported (8). We did not detect such a relationship, possibly owing to the lack of specific frontal lobe subregion sampling (39). However, we found a significant inverse relationship, both before and after PVC, between BDI and 18F-FCWAY BP in hippocampus ipsilateral to the seizure focus (Fig. 3). Interestingly, no significant relationship was detected between BDI and BP or CMRglu in any other region. Previous PET studies in patients with depression reported reduced 5-HT1A receptor binding in regions relevant to epilepsy, such as raphe nuclei and the amygdala-hippocampus complex (40). Additional studies will be necessary to assess the relation of 5-HT1A receptors to depression in TLE.
CONCLUSION
Our study shows the critical importance of PVC for PET receptor studies of TLE patients refractory to medical therapy. Reduction of 5-HT1A BP remained significant in mesial, but not lateral, temporal regions of TLE patients after MR-based PVC. The decrease in 5-HT1A binding exceeded both CMRglu hypometabolism and hippocampal atrophy and could be detected in mesial temporal regions in patients with normal MR scans.
Acknowledgments
The authors thank the entire staff of the NIH PET Department for their technical assistance. The authors thank Nicholas Patronas, MD, and John Butman, MD, for reading MR scans.
Footnotes
Received Dec. 17, 2004; revision accepted Mar. 30, 2005.
For correspondence contact: William H. Theodore, MD, Bldg. 10, Room 5N-250, 9000 Rockville Pike, MSC 1408, National Institutes of Health, Bethesda, MD 20892-1408.
E-mail: theodorw{at}ninds.nih.gov.
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Temporal lobe epilepsy and affective disorders: the role of the subgenual anterior cingulate cortex
- The 5-HT1A receptor and 5-HT transporter in temporal lobe epilepsy
- PET of Serotonin 1A Receptors and Cerebral Glucose Metabolism for Temporal Lobectomy
- Using Cerebral White Matter for Estimation of Nondisplaceable Binding of 5-HT1A Receptors in Temporal Lobe Epilepsy
- Decreased GABA-A binding on FMZ-PET in succinic semialdehyde dehydrogenase deficiency
- Hippocampal volume and depression: insights from epilepsy surgery
- MRI-Based Correction for Partial-Volume Effect Improves Detectability of Intractable Epileptogenic Foci on 123I-Iomazenil Brain SPECT Images
- Striatal D2 Receptor Availability After Shunting in Idiopathic Normal Pressure Hydrocephalus
- Disulfiram Inhibits Defluorination of 18F-FCWAY, Reduces Bone Radioactivity, and Enhances Visualization of Radioligand Binding to Serotonin 5-HT1A Receptors in Human Brain
- Prognosis of children with partial epilepsy: MRI and serial 18FDG-PET
- Hippocampal 1H-MRSI correlates with severity of depression symptoms in temporal lobe epilepsy
- Synthesis and Biologic Evaluation of a Novel Serotonin 5-HT1A Receptor Radioligand, 18F-Labeled Mefway, in Rodents and Imaging by PET in a Nonhuman Primate