


Abstract
Radiolabeled DNA and other oligomers are now under investigation for antisense targeting of a variety of messenger RNA (mRNA). Multidrug resistance (MDR) is detectable as P-glycoprotein (Pgp) expression in most cells and is often elevated in tumor cells, especially those exposed to chemotherapeutic drugs. Radiolabeled antisense DNA has not previously been considered for the targeting of mdr1 mRNA, the product of the mdr1 gene controlling Pgp expression of MDR. Methods: A 20-mer uniform phosphorothioate DNA, described elsewhere as targeting the AUG start codon of mdr1 mRNA, was used naked along with the sense phosphorothioate DNA control. The 3 cell lines were KB-G2, an epidermal carcinoma cell line that had been transfected to overexpress mdr1 mRNA (i.e., Pgp++) compared with its parent (Pgp+) KB-31, and TCO-1, a thyroid carcinoma cell line also reported to be Pgp++. The relative expression of mdr1 mRNA in these 3 cell lines was confirmed elsewhere by reverse transcriptase polymerase chain reaction. As a marker of Pgp expression, the uptake of 99mTc-sestamibi was measured in the 3 cell lines after 20 h of incubation with different concentrations of both antisense and sense DNA. Both DNAs were radiolabeled with 99mTc via mercaptoacetyltriglycine, and cellular uptake was measured after 24 h of incubation. Results: In the case of the sense DNA, the ratio of sestamibi uptake in cells incubated with the DNA to those not exposed to the DNA was unaffected regardless of cell line and regardless of DNA concentration. In contrast, this ratio was significantly higher in both the KB-G2 and TCO-1 cells when incubated with antisense DNA at concentrations greater than about 25 nmol/L (i.e., 150 ng/mL). Only in the KB-31 cells was the sestamibi accumulation unaffected by incubation with the antisense DNA. Thus, the antisense DNA was interfering with Pgp expression to a measurable extent in both Pgp++ cells, but not the Pgp+ cells. This behavior is almost certainly due to antisense targeting of mdr1 mRNA by the antisense DNA since the sense control DNA had no effect. A significant increased accumulation of 99mTc-antisense versus 99mTc-sense DNA was observed in all 3 cell lines. In all cases, this difference was greatest at the lowest DNA concentrations and decreased with increasing concentration as expected for specific binding. In the KB-G2 cells, cellular accumulation of 99mTc-antisense DNA was strikingly high at the lowest concentration at 54%, compared with 22% for 99mTc-sense DNA. These accumulations therefore probably reflect the higher mRNA target concentration in the MDR++ cells than in the MDR+ cells and the higher specific binding of 99mTc-antisense DNA than nonspecific binding of 99mTc-sense DNA. Conclusion: Further evidence was obtained suggesting that an antisense mechanism is responsible for the accumulation of 99mTc-oligomers in cells in culture. Finally, whereas evidence of in vitro targeting is not necessarily evidence of in vivo targeting, our results do suggest that radiolabeled antisense DNA against the mdr1 mRNA may potentially be useful for antisense imaging of MDR in cancer.
Antisense targeting is currently under investigation with radiolabeled DNA and its analogs against a variety of messenger RNA (mRNA) targets, usually in cultured cancer cells (1–5). One potential target may be the mRNA coding for P-glycoprotein (Pgp), the product of the mdr1 gene contributing to multidrug resistance (MDR). Most cancer cells are thought to develop MDR through the overexpression of Pgp, especially after exposure to chemotherapeutic drugs. Although Pgp is elevated in breast and a variety of other cancers (6), the protein is also found in relatively high levels in certain normal tissues such as the adrenals, kidneys, and epithelial cells of the gastrointestinal tract (7). Because the promise of antisense targeting is that any overexpressed mRNA can be targeted with antisense oligomers, a focus on Pgp for antisense targeting may be justified, especially since a noninvasive hot-spot imaging method to assess the MDR status of tumor could be clinically useful.
Three Pgp-expressing cell lines, KB-31, KB-G2, and TCO-1, were used in this investigation. KB-G2 was transfected from its parent human oral epidermoid carcinoma cell line, KB-31, and has been reported to overexpress the target mdr1 mRNA (8). TCO-1, an anaplastic thyroid carcinoma cell line established from a chemotherapy-resistant cancer, was also reported by reverse transcriptase polymerase chain reaction (RT-PCR) to express mdr1 mRNA (9). KB-31 was used as a limited expression control cell line (10) but with caution, since MDR may easily and inadvertently be acquired. A recent measurement of relative levels of Pgp expression by RT-PCR of these 3 cell lines provided estimates for copies of the targeted mdr1 mRNA per microgram of total RNA of 103.56 (KB-31), 106.78 (KB-G2), and 105.53 (TCO-1) (Kumi Nagao, written communication, March 2003). The DNAs used were 20-mer uniformly modified phosphorothioates. The base sequence of the antisense DNA was selected to be against the mdr1 mRNA and had been shown elsewhere to inhibit Pgp in MDR 3T3 cells when incubated at a 100 nmol/L concentration (11). It was used along with its sense control DNA. The DNAs were obtained with a primary amine via a linker on the 5′ end for conjugation with S-acetyl N-hydroxysuccinimide-mercaptoacetyltriglycine (S-acetyl NHS-MAG3) for radiolabeling with 99mTc (1). The DNA was also obtained native (i.e., without linker and amine/amide/MAG3) for use in the 99mTc-sestamibi studies to measure the relative MDR status of the cell lines.
The first objective of this investigation was to confirm that the cell lines showed differing levels of Pgp expression by using 99mTc-sestamibi accumulation. This radiopharmaceutical has been shown to be a substrate of Pgp (12). The second objective was to measure accumulations of the 99mTc-labeled antisense and sense control DNAs in the cell lines at different DNA concentrations in the medium. We hoped to demonstrate a statistically significant antisense differential (i.e., cellular accumulations of antisense DNA minus cellular accumulation of sense DNA) at low concentrations and to demonstrate that this differential can be reduced to insignificance by increasing the DNA concentrations to saturating levels. Achievement of both objectives would provide further evidence of an antisense effect in tissue culture and might suggest that the mdr1 mRNA may be a useful target for antisense imaging of MDR+ cancer.
MATERIALS AND METHODS
The DNAs were obtained both with a primary amine on the 5′ end via a 6-carbon alkyl linker (Operon Technologies) and without the amine or linker (Qiagen K.K.). All DNAs were 20-mer uniform phosphorothioates purified by high-performance liquid chromatography by the supplier. The base sequence of the antisense DNA was 5′-CCA-TCC-CGA-CCT-CGC-GCT-CC, whereas that of the sense control was 5′-GGA-GCG-CGA-GGT-CGG-GAT-GG (11). S-acetyl NHS-MAG3 was synthesized in house (13), and its structure was confirmed by elemental analysis, proton nuclear magnetic resonance, and mass spectroscopy. Bio-Gel P4 medium for separation was purchased (Bio-Rad Laboratories Inc.). The 99mTc-pertechnetate was eluted from a 99Mo–99mTc generator (Daiichi Radioisotopes). The 3 cell lines were a generous gift from Isamu Sugawara (Research Institute of Tuberculosis). Standard chemicals were obtained from various suppliers and used without purification.
DNA Conjugation and Radiolabeling
Both DNAs were conjugated with S-acetyl NHS-MAG3 via the 5′ derivatized amine as previously described (14). A solution of 1 mg of DNA in 200 μL of a 0.2 mol/L concentration of N-(2-hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) buffer, pH 8.0, was added to a vial containing 1.7–2.0 mg of S-acetyl NHS-MAG3. The vial was immediately stirred with a vortex mixer and incubated for 1 h at room temperature. The DNA was then purified on a 0.7 × 20 cm P4 column with either a 0.25 mol/L concentration of NH4OAc buffer, pH 5.2, or a 0.05 mol/L concentration of phosphate-buffered saline (PBS), pH 7.2, as eluent. The peak fractions were pooled, and the DNA concentration was quantitated by ultraviolet absorbance at 265 nm.
DNA was radiolabeled by introducing about 37 MBq (20–40 μL) of 99mTc-pertechnetate generator eluate into a combined solution consisting of 50 μL (about 10 μg) of MAG3-conjugated DNA in either the NH4OAc buffer or PBS, 25 μL of Na2tartrate·2H2O, 50 μg/μL, in a pH 9.2 buffer, and 5 μL of fresh SnCl2·2H2O, 4 μg/μL, in a 10 mmol/L concentration and HCl. The final pH was about 7.8. After stirring with a vortex mixer and then heating for 20 min in boiling water, the labeling was confirmed by C18 SepPak (Waters), in which the first elution with PBS removes radiolabeled pertechnetate and tartrate and the second elution with 40% acetonitrile removes radiolabeled DNA and the SepPak-retains radiolabeled colloids. In most cases, the radiochemical purity exceeded 90% without purification. Each preparation was terminally filtered (0.22 μm) to ensure sterility.
Sestamibi Cellular Accumulation Studies
The cells were seeded at 5 × 105 cells per well in 6-well plates at 2 mL per well and incubated at 37°C in 5% carbon dioxide for 24 h for adherence and growth. The cells were then rinsed twice with PBS. For the sestamibi studies, either antisense or sense DNA without the primary amine attached was added in triplicate to each well at a concentration of up to 500 nmol/L in OptiMEM medium (Gibco BRL Products). Control wells did not receive DNA. The plates were incubated overnight, the supernatant was removed, and 2 mL of Dulbecco’s modified Eagle medium containing 2% fetal bovine serum (FBS) was added before the cells were incubated for a further 24 h. Thereafter, the medium was replaced with Dulbecco’s modified Eagle medium containing 10% fetal calf serum. To each well was added 10 μL of sestamibi (148 kBq 99mTc), and the well plates were incubated at 37°C overnight. The medium was then removed, and 100-μL aliquots of each were saved for counting. Each well was then rinsed twice with PBS, and 2 mL of Bio-Rad A solution (BioRad) was added to each well for cell lysis. An aliquot of the lysate was removed for counting in an automatic NaI (Tl) well counter. After counting, the protein concentration of the cell suspension was determined by adding BioRad B, and counts were corrected for cell number.
DNA Cellular Accumulation Studies
Cells were cultured and seeded at 0.5 × 105 cells per well in 24-well plates. The 99mTc-labeled antisense and sense DNAs were diluted in OptiMEM medium to the desired concentrations in the range 25–1,400 nmol/L. One-half milliliter was added to each well in quadruplet, and the cells were incubated for 20 h at 37°C in 5% carbon dioxide. The cells were harvested, and aliquots of the medium and cell lysate were collected. The percentage of radioactivity that was cell associated was measured against a standard of the added radioactivity. As a further control, wells without cells were treated identically to correct for radioactivity retained on the plate during rinsing and recovered in the lysis solution. This radioactivity was found to be negligible in all cases. After counting, the protein concentration of the cell suspension was determined as before. Cell-associated radioactivity, corrected for cell number, was then plotted against DNA concentration.
RESULTS
Sestamibi Cellular Accumulation Studies
Figure 1 presents the results of measuring sestamibi accumulation in KB-31 cells, KB-G2 cells, and TCO-1 cells. The results have been presented as a ratio of sestamibi accumulated in cells incubated with DNA over that accumulated in cells not receiving the DNA. Each cell line separately received unlabeled antisense and sense DNA at concentrations between 1 and 500 nmol/L. Statistical significance (P < 0.01) was established by the paired Student’s t test with 2-tailed distribution. The figure shows that no difference in the ratio was evident in any cell line incubated with sense DNA at any concentration. Therefore, the control DNA was unable to interfere with Pgp clearance of sestamibi. By contrast, at least in the range of 10–100 nmol/L, the antisense DNA was successful in reducing Pgp clearance of sestamibi from KB-G2 and TCO-1 cells but was unsuccessful in KB-31 cells. However, at concentrations below about 10–25 nmol/L DNA, incubation with the antisense DNA no longer increased sestamibi uptake. These are exactly the results that would be expected should the antisense DNA act on Pgp++ cells by an antisense mechanism to reduce sestamibi clearance. This decreased clearance would then not be expected if control DNA was used in place of the antisense DNA, nor would it be expected if the antisense DNA concentration were too low. Finally, a decrease in clearance might not be expected in a cell line such as KB-31 in which the Pgp expression was low to begin with.
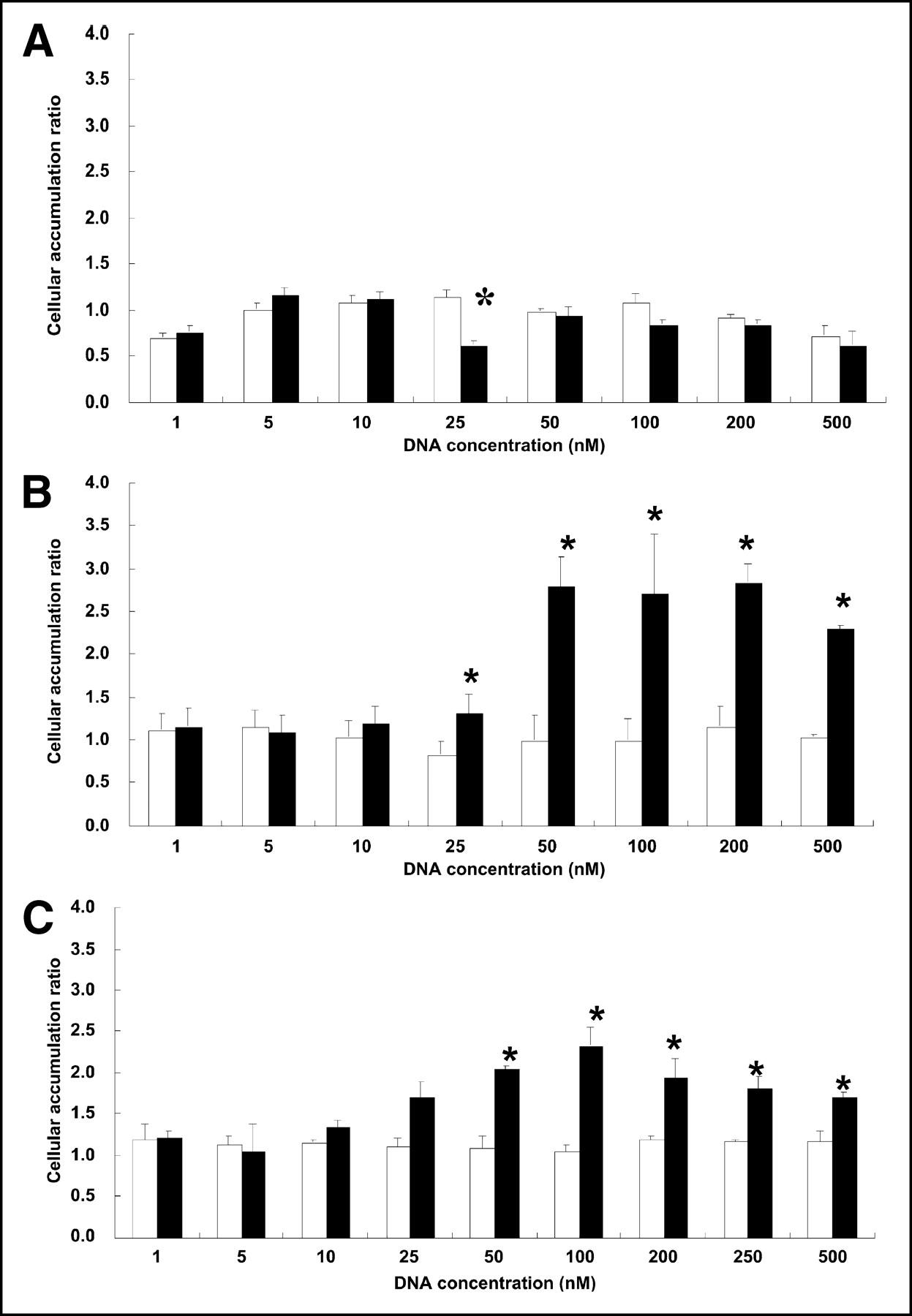
Accumulation of 99mTc-sestamibi in KB-31 cells (A), KB-G2 cells (B), and TCO-1 cells (C) of antisense DNA (black bars) and sense DNA (white bars) for concentrations of DNA between 1 and 500 nmol/L. Results are averaged over the 3 to 4 measurements at each concentration and are presented as ratio of sestamibi in cells incubated with DNA to sestamibi in cells not receiving DNA. Error bars represent 1 SD, and asterisks indicate that differences between ratios obtained with antisense DNA and ratios obtained with sense DNA is statistically significant at that concentration.
DNA Cellular Accumulation Studies
Figure 2 presents the results of measuring 99mTc-antisense and 99mTc-sense DNA accumulations at 20 h in KB-31 cells, KB-G2 cells, and TCO-1 cells incubated with the antisense and sense DNAs at concentrations between 25 and 300–1,400 nmol/L. The results show that the absolute accumulation of 99mTc, whether on antisense or sense DNA, is clearly highest in the KB-G2 cells, reaching a remarkable 54% for antisense DNA in KB-G2 cells at the lowest concentration. This high cellular accumulation is particularly surprising for DNA added naked (i.e., without carrier), as in this investigation.
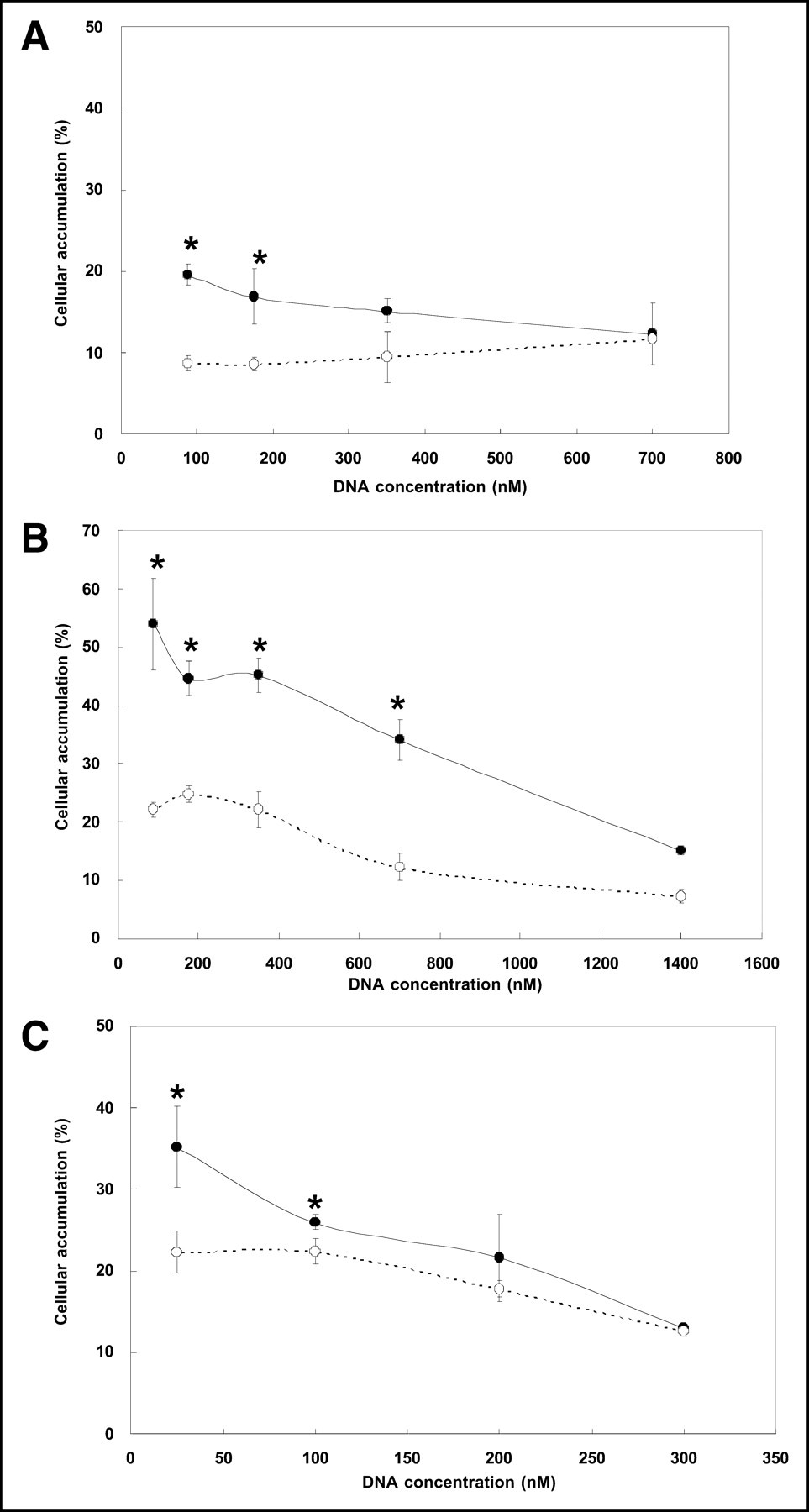
Accumulation of radiolabeled antisense DNA (•) and sense DNA (○) in KB-31 cells (A), KB-G2 cells (B), and TCO-1 cells (C). DNA was added at concentrations between 25 nmol/L (i.e., 150 ng/mL) and higher. Results are average of 4 measurements at each concentration. Error bars represent 1 SD, and asterisks indicate that differences in accumulation at the same concentrations are statistically significant.
That an antisense mechanism may be responsible in part for the accumulation of antisense DNA is evident in that in all 3 cell lines, the antisense differential (i.e., antisense minus sense) was greatest at the lowest DNA concentrations and decreased with increasing concentration as expected for specific binding. Although statistical insignificance was reached at about 300 nmol/L for TCO-1 cells and at about 700 nmol/L for KB-31, a concentration of about 1,400 nmol/L was required to eliminate statistical significance in the KB-G2 cells.
Calculations based on the cellular radioactivity added to a well combined with the known number of cells and the specific accumulation (i.e., antisense DNA accumulation minus sense DNA accumulation) show that about 107 antisense DNA accumulated specifically in each cell line over the 20-h incubation period. Our laboratories have frequently observed specific accumulations of similar magnitude in cell culture for antisense DNAs against different mRNA targets and in different cell lines (5). These values are much higher than expected on the basis on estimates of steady-state mRNA concentrations and require that the concentration of antisense DNA increase over the incubation period from nanomolar in the medium to millimolar in the cellular cytoplasm—a phenomenon requiring some mechanism of selective retention (2).
DISCUSSION
Despite an increasing number of recent reports describing targeting of cells in culture with radiolabeled oligomers by what appears to be an antisense mechanism (5), the weight of evidence is still often considered insufficient to constitute proof of antisense targeting. The case for antisense targeting will be strengthened by additional observation of increased accumulations of radioactivity when added bound to an antisense oligomer compared with a radiolabeled control oligomer differing only in base sequence. Therefore, one objective of this investigation was to continue these studies by consideration of a novel mRNA target.
Pgp is a transmembrane protein and the product of mdr1 mRNA translation, which, in turn, is the product of mdr1 gene expression. Pgp participates in one mechanism of MDR and has been shown responsible for cellular clearance of sestamibi (12). Therefore, a second objective of this investigation was to explore whether a radiolabeled antisense DNA against the mdr1 mRNA might accumulate in cells in rough proportion to the extent of Pgp expression. If so, it may be possible to develop an approach for hot-spot nuclear medicine imaging of increased Pgp expression, in contrast to sestamibi imaging, in which upregulation of Pgp would be expected to decrease accumulation of sestamibi.
The control cell line of this investigation was KB-31 as a “normal” expresser of Pgp (i.e., Pgp+). Our KB-31 cell line and other human oral epidermoid carcinoma cell lines also designated KB-31 have been shown to express the mdr1 mRNA at low levels and therefore may be expected to express Pgp at low levels as well (10). The study cell lines were KB-G2, a transfectant of KB-31 expected to overexpress Pgp (i.e., Pgp++), and TCO-1, also expected to be Pgp++. The result of RT-PCR performed elsewhere and described here tends to confirm these relative expression levels.
Use of sestamibi accumulation as a measure of Pgp expression made it possible to show a difference among the 3 cell lines in the degree to which the antisense DNA interfered with Pgp expression (Fig. 1). Incubation with antisense DNA had no obvious influence on sestamibi accumulation in KB-31 cells, probably because they are low in Pgp expression. That was not the case for KB-G2 and TCO-1 cells, in which incubation with antisense DNA at sufficient concentration significantly increased sestamibi accumulation. These results suggest an antisense mechanism in that a reduction in sestamibi clearance would not be expected should a control sense DNA be used in place of the antisense DNA, or should the concentration of the antisense DNA be too low, or should a particular cell line not express Pgp at sufficient levels to show an effect.
These results may be used to estimate the number of mRNA targets per tumor cell by assuming that the influence of antisense DNA on sestamibi accumulation disappears on target mRNA saturation. Because 1 g of tumor occupies a volume of about 1 mL (i.e., density of 1) then 109 cells per gram of tumor translate into 10−12 L per cell. From the results presented in Figure 1 and assuming an intracellular concentration equal to the medium concentration, saturation at about 1,000 nmol/L is the equivalent of about 107 DNA per cell, or a value roughly equal to that calculated below from the results of incubation with radiolabeled DNA.
Figure 2 presents the accumulation in the 3 cell lines of radiolabeled antisense and sense DNA added at concentrations between 25 nmol/L (i.e., 150 ng/mL) and 1,400 nmol/L (i.e., 8.2 μg/mL) in the case of KB-G2 cells. The figure shows that an antisense differential (i.e., significantly higher accumulation of antisense DNA than of sense control DNA) was observed in all 3 cell lines when the DNA concentration was sufficiently low. Therefore, the mdr1 mRNA target in both KB cell types may be added to the growing list of mRNA targets and cell types for which an antisense differential has been observed in culture (1–5). That the effect is specific, as expected for antisense targeting, is shown by the decreasing antisense differential with increasing DNA concentration. For each cell line, the DNA concentration has been increased to the point at which the accumulation no longer shows a significant antisense differential. This behavior is therefore consistent with a specific accumulation in which the antisense DNA concentration has been increased beyond the point of target mRNA saturation, such that accumulations at higher concentrations are nonspecific. The concentration at which this occurred differed among cell types but was highest in KB-G2 cells, as was expected because this cell line was shown by RT-PCR to express the highest concentration of the target mRNA. This behavior therefore provides further evidence of an antisense mechanism of accumulation.
The high, 50%, accumulation for the antisense DNA in KB-G2 cells was a surprise. Accumulations on this order are not generally seen even when DNA is added with carriers such as cationic peptide or liposome (4). Because of the high accumulation, the number of antisense DNA that specifically accumulated (i.e., antisense DNA minus sense DNA) is calculated at about 107 per cell after 20 h of incubation. This value is higher by about one order of magnitude than that reported previously for different targets (2) and requires that the antisense DNA be concentrated from nanomolar concentrations in the medium to millimolar concentrations in the cell.
The differences in antisense DNA accumulation between the control KB-31 cells and the study cells may be less than expected on the basis of the steady-state mdr1 mRNA levels measured elsewhere by RT-PCR and reported here. One possible explanation is that the antisense DNA accumulation is only partly determined by steady-state target mRNA levels and may be influenced by mRNA production rates as well (2). The less-than-expected difference in cell accumulation between KB-31 and the study cells may also not necessarily suggest poor imaging since in vivo imaging with antisense oligomers will probably include carriers to increase cellular accumulation.
CONCLUSION
These results provide further evidence of the accumulation of 99mTc-oligomers in cells in culture by an antisense mechanism. Not only was an antisense differential observed in all 3 cell lines shown to express the target mRNA, but the differential was also shown to be dependent on DNA concentration. Finally, whereas successful in vitro targeting does not necessarily translate into successful in vivo targeting, these results do suggest that Pgp expression may be a useful target for antisense imaging in cancer using radiolabeled DNA directed against the mdr1 mRNA.
Acknowledgments
The authors thank Dr. Isamu Sugawara (Research Institute of Tuberculosis) for providing the cell lines for this investigation, Dr. Kumi Nagao (Center for Molecular Biology and Cytogenetics, SRL, Inc.) for providing RT-PCR results of relative mdr1 mRNA cellular concentration, and Dr. Ghozheng Liu (University of Massachusetts Medical School) for helping with the radiolabeling of DNAs. This research was supported in part by the Office of Science (BER), U.S. Department of Energy, Grants DE-FG02-99ER62781 and DE-FG02-03ER63602, and a Grant-in-Aid for Scientific Research (13670965) from the Japanese Ministry of Education, Science, Sports and Culture.
Footnotes
Received Apr. 9, 2004; revision accepted Oct. 26, 2004.
For correspondence or reprints contact: Kayoko Nakamura, Department of Radiology, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo, 160-8582 Japan.
E-mail: kumako{at}sc.itc.keio.ac.jp
REFERENCES
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Auger Radiation-Induced, Antisense-Mediated Cytotoxicity of Tumor Cells Using a 3-Component Streptavidin-Delivery Nanoparticle with 111In
- Modulation of drug resistance by artificial transcription factors
- Noninvasive Imaging of Human Telomerase Reverse Transcriptase (hTERT) Messenger RNA with 99mTc-Radiolabeled Antisense Probes in Malignant Tumors
- Cell Culture and Xenograft-Bearing Animal Studies of Radiolabeled Antisense DNA Carrier Nanoparticles with Streptavidin as a Linker
- Initial Mechanistic Studies of Antisense Targeting in Cells