


Abstract
Neutrophil granulocytes play a key role in the pathogenesis of a wide variety of pulmonary diseases. In many such conditions, the injury observed reflects the activation status rather than the total number of inflammatory cells present. The metabolic activity of neutrophils can now be assessed noninvasively using PET to measure the regional uptake of 18F-FDG after intravenous injection. Methods: To understand the mechanism responsible for the increased 18F-FDG signal, we have measured the uptake of tritiated deoxyglucose (DG) in neutrophils isolated from human peripheral blood and sought to determine which aspects of neutrophil function correlate with an increase in DG uptake. Results: Our results show that formyl-methionyl-leucyl-phenylalanine (fMLP)-stimulated respiratory burst activity and 3H-DG uptake are temporally dissociated, that neutrophil-priming agents such as tumor necrosis factor-α (TNFα) cause an identical increase in 3H-DG uptake compared with fMLP without affecting respiratory burst activity, and that fMLP stimulation of TNFα-primed cells causes major upregulation of superoxide anion generation (O2−) yet no incremental increase in 3H-DG uptake. Furthermore, direct activation of reduced nicotinamide adenine dinucleotide phosphate oxidase activity with the ester phorbol 12-myristate 13-acetate resulted in a concentration-dependent loss of cell-associated 3H-DG, and preincubation of neutrophils with the phosphoinositide 3-kinase inhibitor wortmannin, which abolished both agonist-stimulated superoxide anion generation and degranulation, had no effect on TNFα- or fMLP-stimulated 3H-DG uptake. In contrast, the fMLP-stimulated change in neutrophil shape was not influenced by priming or wortmannin, and of the functional responses examined, this appeared to correlate most closely with 3H-DG uptake. Conclusion: DG uptake occurs in both primed and activated neutrophils. It does not correlate with respiratory burst or secretory activity but may reflect the polarization and migrational status of these cells. These data have important implications for the analysis of 18F-FDG signals in vivo.
External imaging of radiolabeled markers of inflammatory processes has great potential for providing noninvasive and repeatable methods of monitoring inflammatory cell behavior in vivo. The use of neutrophils radiolabeled ex vivo with 111In and monitored by γ-scintigraphy has given invaluable information about the migration and emigration of these in response to inflammatory stimuli (1–5) but is limited to monitoring cell trafficking and gives no indication of their postmigratory metabolic activity.
Neutrophils become highly activated in response to a variety of inflammatory stimuli, and the consequent increase in their energy requirements results in enhanced glucose uptake and metabolism. This allows the use of radiolabeled glucose analogues to monitor the metabolic activity of inflammatory cells by external imaging. In previous studies in which acute lung inflammation has been induced by localized instillation of Streptococcus pneumoniae into the right upper lobe of rabbit lungs, we have shown that PET imaging after injection of 18F-FDG provides a remarkably specific index of neutrophil metabolic activity in vivo (6). The increased signal occurred at a time when neutrophil migration had effectively ceased and hence was thought to reflect the activity of the cells within the inflammatory site. Autoradiography using tritiated deoxyglucose (3H-DG) confirmed that the signals obtained in this model were derived almost exclusively from the neutrophils (6), and identical data have now been obtained for other animal models after instillation of both fibrogenic and nonfibrogenic agents (7). We have also shown a dissociation between neutrophil migration and neutrophil activation in human lung disease (8) and have used this technique to show increased 18F-FDG uptake in the lungs of patients known to be at risk of adult respiratory distress syndrome after head trauma, at a time when there are no respiratory symptoms or radiologic abnormalities (9).
The neutrophil is recognized to be a key effector cell of the immune system and plays an essential role in host defense. To serve this function, it has evolved a highly efficient but limited repertoire of functional responses (including chemotaxis, phagocytosis, degranulation, and the generation of toxic reactive oxygen intermediates) that allow it to target and respond to a bacterial insult in the host (10). Moreover, neutrophil priming by agents such as tumor necrosis factor-α (TNFα) is recognized to cause a dramatic increase in many of the responses of neutrophils to activating agents, and this process has been shown to be critical for neutrophil-mediated tissue injury. These events are highly coordinated and, in an ideal situation, permit rapid, transient, and targeted accumulation of neutrophils at an inflamed site and efficient bacterial uptake and clearance. The subsequent removal of granulocytes from the inflamed site is thought to reflect the capacity of these cells to undergo constitutive apoptosis, which results in their prompt recognition and phagocytic removal by inflammatory macrophages (11). In view of the clinical and experimental data highlighting the utility of 18F-FDG to monitor neutrophil activation in vivo and the known spectrum of functional responses observable in these cells, we have sought to investigate which aspects of neutrophil function are responsible for 18F-FDG uptake.
MATERIALS AND METHODS
Materials
Percoll, dextran, and 3H-DG were obtained from Amersham Pharmacia Biotech U.K. Ltd. (Buckinghamshire, U.K.). Cytochrome C, superoxide dismutase, phorbol 12-myristate 13-acetate (PMA), formyl-methionyl-leucyl-phenylalanine (fMLP), phosphate-buffered saline (PBS, with or without CaCl2 and MgCl2), Triton X-100, and iodoacetic acid were purchased from Sigma Chemical Co. (Poole, Dorset, U.K.). Human recombinant TNFα was obtained from R&D Systems (Abingdon, U.K.), wortmannin was purchased from Calbiochem (San Diego, CA), and Soluene-350 and liquid scintillation counting cocktail (Hionic-Fluor) were purchased from Canberra Packard (Pangbourne, Berkshire, U.K.).
Preparation of Human Neutrophils
Human neutrophils were prepared from the peripheral blood of healthy volunteers using dextran sedimentation and discontinuous plasma-Percoll gradients as previously described (12). Sterile, lipopolysaccharide-free reagents and plasticware were used throughout. This method routinely yielded unprimed and fully viable neutrophils that were >95% pure, with <0.1% mononuclear cell contamination.
Superoxide Anion (O2−) Release
Purified neutrophils (106 cells in 90 μL PBS containing CaCl2 and MgCl2) were equilibrated at 37°C in a gently shaking water bath for 5 min. Cells were primed for 30 min with 200 U/mL TNFα and then stimulated for 10 min with fMLP (100 nmol/L) or vehicle (PBS) in the presence of prewarmed cytochrome C (final concentration, 1.2 mg/mL in PBS). One of each set of quadruplicate determinations included superoxide dismutase (375 U) to confirm the specificity of the cytochrome C reduction. Reactions were stopped by placing the cells on ice, and the supernatants were collected after centrifugation. The reduction of cytochrome C was determined by measuring the peak absorbance between 535 and 565 nm using a scanning spectrophotometer. O2 generation (expressed as nmoles O2 generated per 106 neutrophils) was calculated from the difference between values obtained in the presence and absence of superoxide dismutase. Lucigenin-dependent chemiluminescence was used to measure the kinetics of O2− release. Lucigenin (0.25 mmol/L) was added to each well and allowed to equilibrate at 37°C before the addition of TNFα primed or unprimed cells (106 cells per well). Buffer or fMLP (100 nmol/L) was added, and the chemiluminescence was recorded continuously at 9-s intervals for 4 min. Data were recorded online (Cellular Chemiluminescence; Dynatech Laboratories Ltd., Billingshurt, U.K.) to produce mean lucigenin-dependent chemiluminescence values from triplicate wells.
Measurement of 3H-Deoyglucose Uptake in Human Neutrophils
3H-DG was used to measure DG uptake by neutrophils. 3H-DG and 18F-FDG give qualitatively identical information in that both agents gain cellular access by the same glucose-specific uptake mechanism and are converted to the 6-phosphate form, which can be metabolized no further and remains within the cell (13,14). Moreover, both exogenous radioligands have been shown to accumulate within metabolically active cells in proportion to their glucose requirements (13). Neutrophils (106 cells) were suspended in 90 μL PBS containing Ca2+ and Mg2+ cations and incubated with TNFα (200 U/mL) or PBS for 30 min at 37°C. Immediately after the addition of 370 kBq 3H-DG, the cells were stimulated with fMLP (100 nmol/L), PMA (0.1–1,000 nmol/L), or PBS (total volume, 1 mL). Uptake of 3H-DG was stopped by the addition of iodoacetic acid (100 nmol/L). After centrifugation, the supernatants were decanted and the cells were washed twice with PBS to remove all unincorporated 3H-DG. Cell pellets were dissolved in Soluene-350, and radioactivity in the cell pellets and supernatants was measured by liquid scintillation counting. The uptake of 3H-DG by neutrophils was calculated from the ratio of 3H-DG in the cell pellet to that in the supernatant. For inhibitor studies, cells were preincubated with wortmannin (100 nmol/L) or vehicle (0.01% dimethyl sulfoxide [DMSO]) for 10 min before 3H-DG addition and agonist stimulation.
Measurement of Degranulation in Human Neutrophils
Myeloperoxidase (MPO) release was determined by the 3,3-dimethoxybenzidine method as described previously (15), with the following minor modifications. Briefly, 106 neutrophils were suspended in PBS containing Ca2+ and Mg2+ ions and incubated with TNFα (200 U/mL) or PBS at 37°C (30 min), followed by stimulation with fMLP (100 nmol/L) or PBS (10 min). For inhibitor studies, cells were preincubated with wortmannin (100 nmol/L) or DMSO (0.01%) for 10 min before agonist stimulation. One of each set of quadruplicate determinations included Triton X-100 (0.2%) to estimate the total enzymatic activity. Reactions were quenched by the addition of ice-cold PBS and immersion in ice. After centrifugation (12,500g, 2 min, 4°C), the supernatants were incubated with phosphate buffer (pH 6.2), 3,3-dimethoxybenzidine (0.125 mg/mL), and H2O2 (0.001%) for 20 min at 37°C. Reactions were terminated by the addition of 0.1% NaN3, and the samples were cooled to room temperature. The MPO activity was measured by spectrophotometric assessment (460 nm) (Ultrospec III; Pharmacia, Amersham Biosciences UK Ltd., Little Chalfont, Bucks, U.K.), and the amount of MPO released was expressed as a percentage of the total MPO available from cells lysed by Triton X-100.
Assessment of Agonist-Stimulated Change in Neutrophil Shape
Neutrophils were isolated as detailed above, equilibrated in PBS at 37°C for 5 min, and incubated with PBS or wortmannin (100 nmol/L) for 10 min before a further 10-min incubation with varying concentrations of fMLP. This time point was determined in preliminary experiments that indicated that the fMLP-induced change in neutrophil shape was maximal at 2–5 min and did not vary over the subsequent 30 min. Reactions were stopped by the addition of an equal volume of 2.5% glutaraldehyde. The shape of cells (minimum of 300 cells per condition) was then visually assessed for change using a light microscope (10).
Statistics
All values are expressed as the mean ± SEM of the number of independent experiments. The data were evaluated statistically by ANOVA, with comparisons between groups made using the Newman-Keuls test. Differences were considered significant when P was <0.05.
RESULTS
As previously shown (16), the ability of secretagogue agonists such as fMLP to stimulate respiratory burst activity in isolated neutrophils is highly dependent on the priming status of these cells. Hence, preincubation of neutrophils with TNFα, which alone has little effect on superoxide anion generation, caused a major increase in fMLP-stimulated respiratory burst activity (Fig. 1A). In contrast, when 3H-DG uptake was assessed under identical conditions, incubation of neutrophils with TNFα or fMLP alone resulted in a near identical increase in 3H-DG, with no significant augmentation of fMLP-stimulated 3H-DG uptake observed after TNFα priming (Fig. 1B). Moreover, detailed analysis of the time course of superoxide anion generation in neutrophils indicated a clear dissociation between the kinetics of fMLP-stimulated respiratory burst activity and 3H-DG uptake, with the former response being maximal at 2 min (Fig. 1C) and the latter response occurring between 5 and 30 min after addition of fMLP (Fig. 1B).

TNFα priming of fMLP-stimulated O2− generation and 3H-DG uptake in human neutrophils. (A) Measurement of O2− release by superoxide dismutase-inhibitable reduction of cytochrome C during 10-min incubation with fMLP (100 nmol/L) after priming with TNFα (200 U/mL, 30 min). Data points represent mean ± SEM of 3 separate experiments, each performed in triplicate. (B) 3H-DG uptake in TNFα (200 U/mL, 30 min)-primed neutrophils. After priming, or incubation with PBS, cells were stimulated with fMLP (100 nmol/L) or PMA for indicated time. Data shown are for control unprimed cells (○), TNFα-primed (200 U/mL, 30 min) unstimulated cells (□), unprimed fMLP (100 nmol/L)-stimulated cells (•), TNFα-primed fMLP-stimulated cells (▪), and PMA (▵). Results are mean ± SEM of 5 separate experiments. (C) Quantification of O2− release with lucigenin-dependent chemiluminescence monitored at 9-s-cycle intervals in control unprimed cells (○), unprimed fMLP (100 nmol/L)-stimulated cells (•), and TNFα-primed fMLP-stimulated cells (▪). Data are from single experiment, representative of 3 separate experiments.
When neutrophils were stimulated with the phorbol ester PMA, which causes direct, nonreceptor-dependent activation of respiratory burst activity, a marked difference was again apparent between respiratory burst activity and 3H-DG uptake. PMA (100 nmol/L) was found to increase O2− generation to levels considerably higher than those observed in TNFα-primed, fMLP-stimulated cells (Fig. 2A). However, whereas both fMLP and TNFα stimulation caused a sustained increase in 3H-DG accumulation, PMA caused only a transient increase in 3H-DG uptake to levels equivalent to those seen with TNFα or fMLP stimulation (Fig. 1B). At later time points (>5 min), cell-associated 3H-DG was reduced by PMA to less than control levels (Fig. 1B). In subsequent experiments, PMA was found to cause a concentration-dependent reduction in cell-associated 3H-DG at 30 min (Fig. 2B).

(A). Superoxide anion release from control unstimulated cells. Cells were primed with TNFα (200 U/mL) for 30 min, stimulated with fMLP (100 nmol/L) for 10 min, and treated with 100 nmol/L PMA for 30 min. O2− release was measured by the superoxide dismutase-inhibitable reduction of cytochrome C. Data points represent mean ± SEM of 3 separate experiments, each performed in triplicate. (B) Effect of PMA concentration on 3H-DG uptake. Cells were incubated with 3H-DG; radioactivity in cells was measured at 30 min.
Respiratory burst activity occurs through a phosphoinositide 3 (PI3)-kinase-dependent signaling pathway (17,18). This was confirmed by preincubating neutrophils with the PI3-kinase inhibitor wortmannin, which abolished fMLP-stimulated O2− generation and MPO release (Figs. 3A and 3B). Despite this inhibition of agonist-stimulated O2− generation, wortmannin had no effect on fMLP-stimulated 3H-DG uptake in either TNFα-primed or TNFα-unprimed cells (Fig. 3C). The dissociation between neutrophil secretory events and 3H-DG uptake was further confirmed by measuring the release of MPO as a marker of neutrophil degranulation. This response displays rapid kinetics that parallel respiratory burst activity. In these experiments, significant degranulation was observed in only TNFα-primed, fMLP-stimulated cells and was again inhibited by wortmannin (Fig. 3B). Hence, degranulation responses appeared to be similar to the O2− responses and to fail to mimic the pattern of 3H-DG uptake. In contrast, the agonist-stimulated change in neutrophil shape or polarization responses is not dependent on priming and can be initiated independently by TNFα and fMLP alone (10). Furthermore, the agonist-stimulated change in shape is also one of the few neutrophil responses not influenced by wortmannin (Fig. 4).
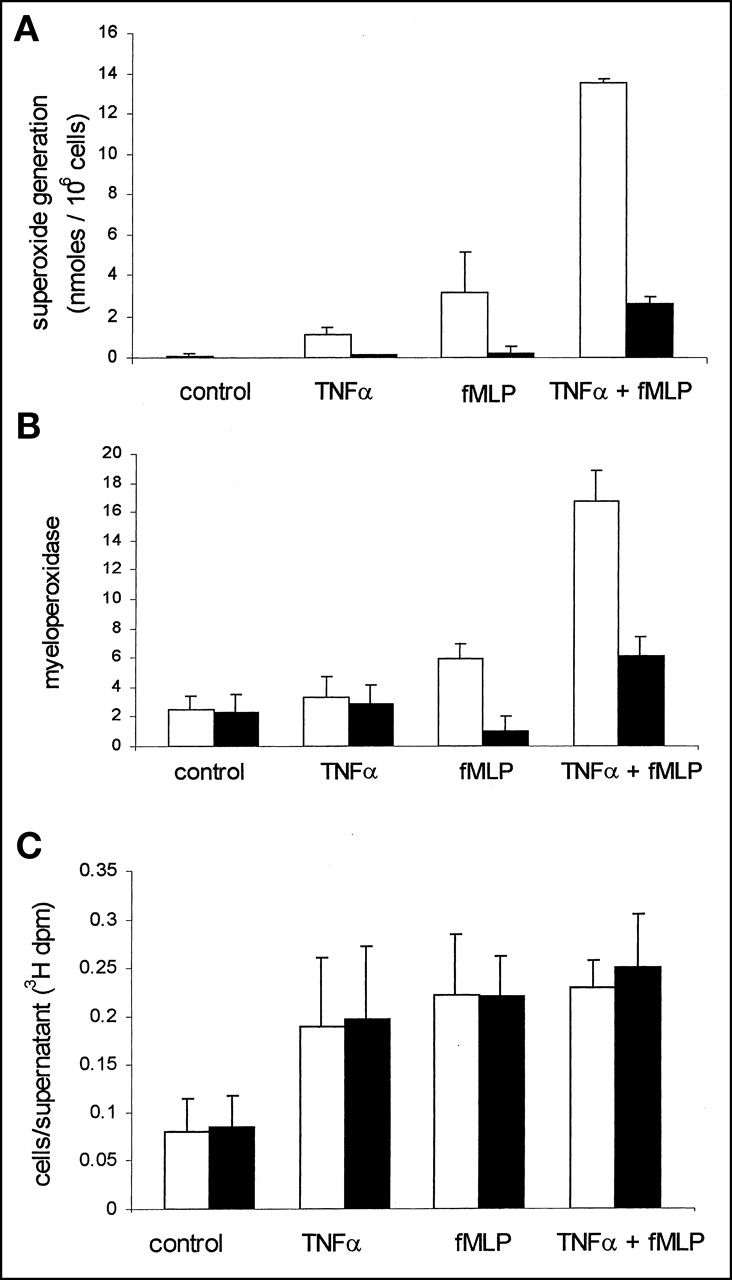
Effect of PI3-kinase inhibitor wortmannin on O2− release, degranulation, and DG uptake. (A) Human neutrophils were incubated with vehicle (0.01% DMSO) (open bars) or 100 nmol/L wortmannin (filled bars) for 10 min before TNFα priming (200 U/mL for 30 min) and fMLP stimulation (100 nmol/L, 10 min). O2− release was quantified by superoxide dismutase-inhibitable reduction of cytochrome C. Data points represent mean ± SEM of 3 separate experiments, each performed in triplicate. (B) Wortmannin inhibits neutrophil degranulation. fMLP-induced MPO release in TNFα-primed human neutrophils was performed as described. Human neutrophils (12.5 × 106/mL) were suspended in PBS with Ca2+ and Mg2+ and treated with TNFα (200 U/mL) or PBS at 37°C for 30 min. Cells were then incubated with 100 nmol/L wortmannin (filled bars) or vehicle (0.01% DMSO) (open bars) for 10 min before stimulation with fMLP (100 nmol/L) or PBS. Reaction was terminated at 10 min by addition of ice-cold PBS and then immersing tubes in ice. MPO release was quantified by spectrophotometric assessment (460 nm) as described previously (15). Data represent mean ± SEM of 3 separate experiments, each performed in triplicate. (C) Wortmannin does not affect 3H-DG uptake. Human neutrophils (12.5 × 106/mL) were suspended in PBS with Ca2+ and Mg2+ and treated with TNFα (200 U/mL) or PBS at 37°C for 30 min. Cells were then incubated with 100 nmol/L wortmannin (filled bars) or vehicle (0.01% DMSO) (open bars) for 10 min before incubation with 3H-DG and stimulation with fMLP (100 nmol/L) or PBS. Data are shown for uptake over 30 min. dpm = disintegrations per minute.
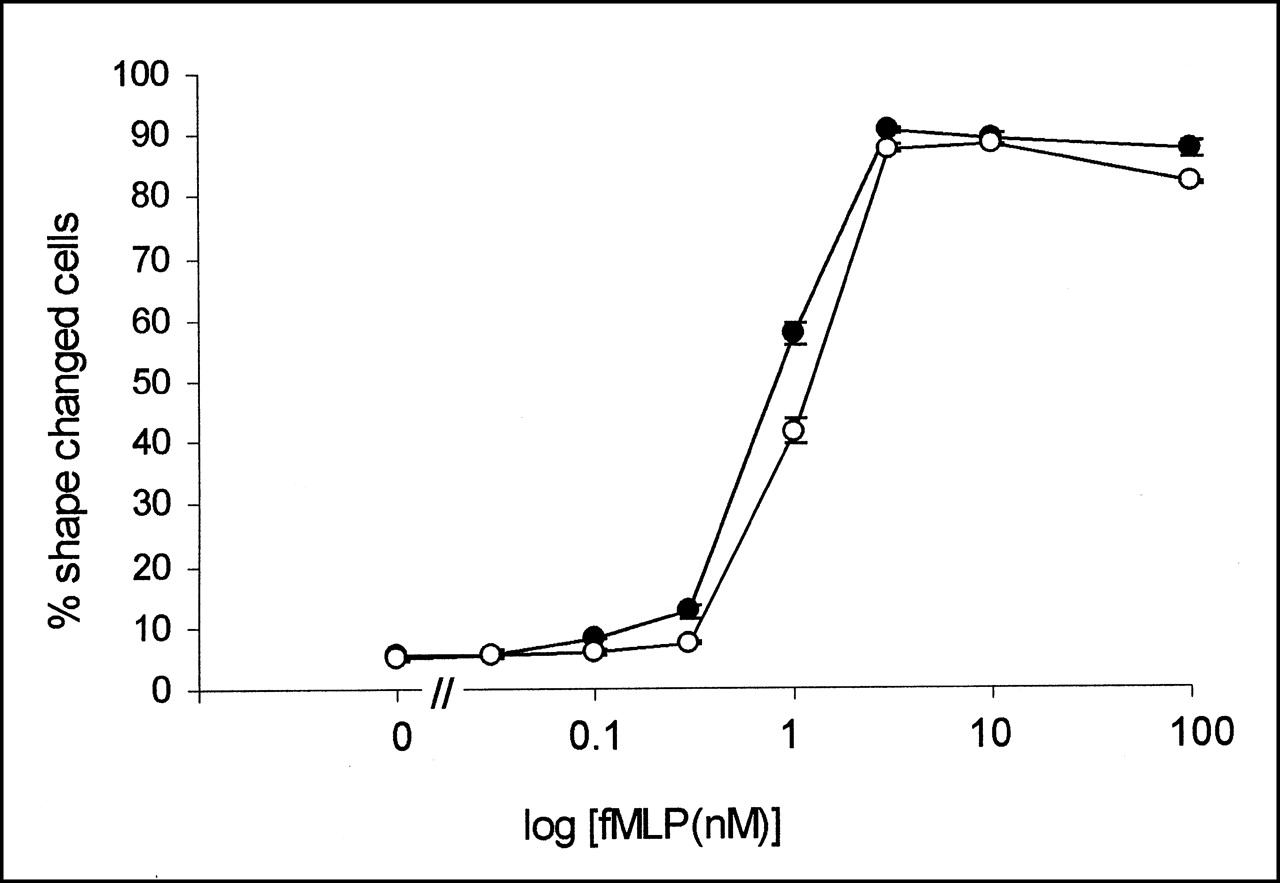
Wortmannin has no effect on fMLP-stimulated change in neutrophil shape. Neutrophils were incubated with increasing concentrations of fMLP with (○) or without (•) wortmannin (100 nmol/L). After addition of glutaraldehyde, shape of cells was visually assessed for change using light microscope. Data represent mean ± SEM of 3 separate experiments, each performed in triplicate.
DISCUSSION
The realization that the relatively high number of false-positive findings encountered in 18F-FDG PET imaging of cancer, especially lung cancer, is the result of uptake in inflammatory lesions has prompted an increasing interest in the use of 18F-FDG PET for imaging and locating inflammation itself. Previous work by our group focused on the mechanism of 18F-FDG uptake in pulmonary inflammation, both the acute inflammation from experimental streptococcal lobar pneumonia or C5a instillation and the chronic inflammation from instillation of a range of mediators of chronic inflammation, including bleomycin and silica particles (7). That study clearly implicated the neutrophil as the principal cell accumulating DG in such inflammatory lesions and, in the case of acute lobar pneumonia, suggested that the cellular process occurred after the neutrophils had migrated into the air spaces (6).
Extension of these studies (6,7) to human lung disease produced some surprising results. Thus, whereas 18F-FDG uptake in acute lobar pneumonia was avid, reflecting the findings in experimental lobar pneumonia, no uptake was seen in bronchiectasis (8), even though this is a condition associated with heavy neutrophil migration as shown with 111In-labeled neutrophil scintigraphy (19). Hence, transendothelial migration of neutrophils into tissue per se is not necessarily accompanied by a significant increase in 18F-FDG uptake. Negative 18F-FDG PET findings in bronchiectasis suggests a dissociation between neutrophil accumulation and activation events, although it is possible that, because the migrating neutrophils in bronchiectasis gain early access to the bronchial lumen, they may not be accessible to circulating 18F-FDG at a time when they are glucose avid. This possibility is unlikely, because the 18F-FDG signal is strongly positive in inflammatory bowel disease (20), a condition also associated with early migration of neutrophils into an inaccessible lumen (the gut). Other inflammatory cells in inflammatory bowel disease, namely lymphocytes and macrophages, may also contribute to a positive 18F-FDG PET signal. To date, no studies have been published that address the direct effect of various inflammatory conditions on DG uptake and retention in circulating neutrophils.
Against this background, it is apparent that an improved understanding of the mechanisms by which neutrophils accumulate 18F-FDG in inflammation would be helpful. The neutrophil has a repertoire of functional responses, which are largely determined by the cell-to-cell contact required for successful transendothelial neutrophil migration at a site of inflammation and the influence of locally generated cytokines and other stimuli. We predicted that the major contributor to the increased DG uptake by stimulated neutrophils would be respiratory burst activity or degranulation events. Neutrophils depend on glucose as their major energy source, and a dramatic increase in uptake and phosphorylation of DG has previously been observed after stimulation in vitro. The current findings show clearly that the cellular events driving cell DG uptake are not related to superoxide generation or degranulation but most likely reflect cell polarization and shape change. This conclusion is based partly on the finding that neutrophil priming alone causes maximal DG uptake in the absence of stimulated superoxide anion generation or MPO release. Thus, DG uptake was closely associated with priming, initiated either by TNFα or by fMLP, and was not significantly increased in response to fMLP exposure after priming with TNFα, a sequence of events that leads to a marked increase in respiratory burst activity and degranulation. Moreover, inhibition of superoxide and enzyme release by the PI3-kinase inhibitor wortmannin was not accompanied by inhibition of DG uptake. The failure of wortmannin to inhibit DG uptake in response to sequential TNFα and fMLP was accompanied by a failure to prevent shape change, again pointing to a link between shape change and DG uptake. The association of cell depolarization with increased glucose uptake when cell migration itself is not inevitably accompanied by an increased 18F-FDG PET signal, as in bronchiectasis, implies that neutrophils can migrate without necessarily first being primed.
The phorbol ester PMA causes direct activation of the reduced nicotinamide adenine dinucleotide phosphate oxidase and stimulated a major release of superoxide anions even in unprimed cells (Fig. 2A). At 5 min, PMA caused an early DG uptake of similar magnitude to the other stimuli. This uptake was followed by a loss of cell-associated label during the ensuing 25 min (Fig. 1B). The cause of this loss from intact neutrophils is unclear but may stem from increased cellular permeability to DG or DG-6-P, upregulation of dephosphorylation enzymes, or even the release of DG or the DG-6-P form in secretory granules. Although the precise mechanism of this effect of PMA on DG uptake by neutrophils is unclear, these data provide further evidence of a dissociation between respiratory burst activity and DG uptake.
If, as suggested by this study, DG uptake by neutrophils is a marker of neutrophil priming, one would expect to see diffuse pulmonary activity after administration of 18F-FDG in patients with conditions such as inflammatory bowel disease, graft-versus-host disease, and systemic vasculitis, which have an elevated circulating level of shape-changed (primed) neutrophils and which usually show an increased, reversible diffuse pulmonary accumulation of radiolabeled neutrophils in the absence of any respiratory symptoms (21). Although such an increase in the 18F-FDG signal has not as yet been studied under such conditions, we have seen increased diffuse uptake of 18F-FDG in patients with acute head injury but no respiratory symptoms (9).
CONCLUSION
As 18F-FDG becomes more frequently used in the investigation of inflammatory disease, we will need to understand its mechanism of uptake by, and its selectivity for, the major cellular players in the inflammatory response. Although we have shown that the neutrophil is the cell responsible for the larger part of the 18F-FDG PET signal in both acute and chronic inflammatory responses in the lung (7), we need to define the relationship between DG uptake and activation status in other cells, including endothelial cells, as well as inflammatory cells such as lymphocytes and macrophages. The current study strongly suggests that as far as neutrophils are concerned, priming is the cellular event predominantly responsible for 18F-FDG uptake. A greater knowledge of the mechanisms that result in increased DG uptake improves our ability to interpret data acquired by 18F-FDG PET. The ability to monitor this marker of neutrophil activation status repeatedly and noninvasively provides a powerful opportunity to evaluate different therapeutic strategies in patients with inflammatory disease.
Acknowledgments
This study was supported by the Wellcome Trust, Medical Research Council, U.K., and by the Papworth Hospital NHS Trust, Papworth Everard, U.K.
Footnotes
Received Sep. 17, 2001; revision accepted Jan. 23, 2002.
For correspondence contact: Hazel A. Jones, PhD, Respiratory Medicine, Imperial College School of Medicine, Hammersmith Campus, London W12 0NN, U.K.
E-mail: hazel.jones{at}ic.ac.uk
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Imaging Pulmonary Inflammation
- PET/CT in nononcological lung diseases: current applications and future perspectives
- Can the Inflammatory Response Be Evaluated Using 18F-FDG Within Zones of Microvascular Obstruction After Myocardial Infarction?
- 18F-FDG Kinetics Parameters Depend on the Mechanism of Injury in Early Experimental Acute Respiratory Distress Syndrome
- Noninvasive Quantitative Assessment of Pulmonary Blood Flow with 18F-FDG PET
- 18F-FDG Uptake Rate Is a Biomarker of Eosinophilic Inflammation and Airway Response in Asthma
- Idiopathic Pulmonary Fibrosis and Diffuse Parenchymal Lung Disease: Implications from Initial Experience with 18F-FDG PET/CT
- Noninvasive Assessment of Crohn's Disease Intestinal Lesions with 18F-FDG PET/CT
- Effect of Corticosteroids on 18F-FDG Uptake in Tumor Lesions After Chemotherapy
- 18F-FDG PET in Rheumatoid Arthritis: There Still Is a Long Way to Go
- Tetraphenylphosphonium as a Novel Molecular Probe for Imaging Tumors
- Augmented 18F-FDG Uptake in Activated Monocytes Occurs During the Priming Process and Involves Tyrosine Kinases and Protein Kinase C
- In vivo assessment of lung inflammatory cell activity in patients with COPD and asthma