


Abstract
Activated monocytes with a high 18F-FDG accumulation can affect the results of clinical PET studies. To better understand the mechanisms regulating monocytic 18F-FDG uptake, we investigated the effect of priming and respiratory-burst generation and further evaluated the role of potential protein kinase pathways. Methods: Purified human monocytes were primed with interferon-γ (IFN-γ), and respiratory burst was generated by stimulation of primed cells with phorbol-12-myristate-13-acetate (PMA). Oxygen-intermediate generation was assessed by luminescence measurements after the addition of lucigenin. 18F-FDG uptake after 30 min of incubation was measured for unprimed control cells, primed cells, and PMA-stimulated cells. The role of protein kinases was investigated using respective inhibitors. Results: PMA stimulation of primed monocytes dramatically increased oxygen-intermediate generation, leading to a 42.2 ± 1.1 fold higher level of cumulative luminescence compared with unprimed control cells, whereas IFN-γ priming alone resulted in low luminescence levels (13.9% ± 4.6% of PMA-stimulated cells). In contrast, priming alone was sufficient to augment monocytic 18F-FDG uptake to 273.3% ± 16.7% of control levels (P < 0.001), and it was not further increased by PMA stimulation. The tyrosine kinase inhibitor, genistein, and the specific protein kinase C inhibitor, staurosporine, completely abolished the priming-induced enhancement of 18F-FDG uptake and lowered uptake to control levels. Under the same conditions, wortmannin, a phosphatidylinositol 3 kinase (PI3 kinase)-specific inhibitor, and cycloheximide, a protein synthesis inhibitor, were associated with only minor reductions in the enhanced-uptake effect of priming. Conclusion: IFN-γ priming alone, without stimulation of respiratory-burst activity, is sufficient to induce maximal augmentation of 18F-FDG uptake in monocytes. Furthermore, this metabolic effect appears to involve tyrosine kinases and the protein kinase C pathway but is independent of the PI3 kinase pathway.
Monocytes are key effector cells of the immune system and have a repertoire of functional responses, including chemotaxis, phagocytosis, and the generation of toxic reactive oxygen intermediates (1). Because monocytes are one of the principal 18F-FDG-accumulating cells within inflammatory lesions and in certain tumors, their level of glucose metabolism may influence the interpretation of PET studies (2–4). Consequently, an improved understanding of the mechanisms involved in the regulation of 18F-FDG uptake in activated monocytes would be helpful.
Complete activation of monocytes results from 2 sequential steps: priming and a second triggering stimulus. The T cell-produced cytokine interferon-γ (IFN-γ) is considered the major priming signal, whereas the triggering signal includes various stimuli such as bacterial endotoxin, tumor necrosis factor-α, and chemical agents such as phorbol-12-myristate-13-acetate (PMA) (5). A remarkable trait of monocytes is their ability to enhance their oxidative metabolism in response to activating factors, leading to the production of superoxide ions, hydrogen peroxide, and hydrogen radicals referred to as respiratory burst. This response by monocytes is characterized by a dramatic increase in oxygen consumption and glucose metabolism (6). However, the precise conditions and machinery through which activated monocytes acquire augmented levels of 18F-FDG uptake are not clear. We thus investigated the effect of IFN-γ priming and PMA stimulation of monocytes on respiratory-burst generation and 18F-FDG uptake. We further evaluated the role of potential signal transduction pathways by using their respective protein kinase inhibitors.
MATERIALS AND METHODS
Cell Preparation
Fresh circulating monocytes were purified from the peripheral venous blood of healthy human volunteers. Whole blood, collected in acid citrate dextrose-treated syringes and transferred to 16 × 150 mm screw-cap Falcon round-bottom tubes, was diluted in equal volumes of phosphate-buffered saline and mixed with Ficoll/Hypaque 1077 (Sigma) at a ratio of 1:4. The mononuclear cell layer was collected, washed twice, and resuspended in 10% fetal bovine serum-supplemented RPMI-1640 medium (Gibco BRL) containing 2 g of glucose per liter and 100 U of penicillin-streptomycin per milliliter.
After the cells were incubated in a flat-bottomed 150-mm plastic dish at 37°C with 5% CO2 for 1 h, free-floating lymphocytes were collected and transferred to a separate dish. Monocytes attached to the plastic surface were collected with careful pipetting and washing with phosphate-buffered saline. The cell purity of the monocyte preparation was determined with flow cytometry and microscopic inspection after Wright staining.
Cell Priming and Respiratory-Burst Activation
For priming, 0.2–1 × 106 cells were transferred to culture tubes containing 500 μL of culture medium and treated with a human recombinant IFN-γ (Sigma) concentration of 100 U/mL for 3 d. Respiratory-burst activity was induced by stimulating primed cells with 100 nmol of PMA (Sigma) per liter for 20 min. Development of respiratory burst was confirmed by chemiluminescent measurements of oxygen intermediates for 30 min immediately after the addition of a lucigenin (Sigma) concentration of 250 μmol/L. The luminometer was programmed to output raw data of relative light units every 30 s, and measurement of control tubes using this method generated only background numbers.
Measurement of Cellular 18F-FDG Uptake
Monocytes were suspended in 500 μL of Hanks’ balanced salt solution and incubated with 370 kBq of 18F-FDG at 37°C in a CO2 incubator for 30 min. The cells were centrifuged and then washed twice with 500 μL of phosphate-buffered saline. Radioactive counts from the pellets were measured on a high-energy γ-counter (Wallac). 18F-FDG uptake levels were corrected for protein content in each sample, as determined by the Bradford method. Results were expressed as percentage uptake relative to the mean radioactivity for unprimed control cells.
Protein Kinase Inhibition Experiments
To determine the relationship between the effect of IFN-γ priming on 18F-FDG uptake and protein kinase activity, cells were treated with staurosporine (4 μmol/L), a specific protein kinase C (PKC) inhibitor; wortmannin (200 nmol/L), a specific phosphatidylinositol 3 kinase (PI3 kinase) inhibitor; genistein (500 μmol/L), a tyrosine kinase inhibitor; or cycloheximide (100 ng/mL), a protein synthesis inhibitor. The inhibitors were added to monocytes 1 h before the initiation of priming with IFN-γ.
Statistical Analysis
All experiments were performed with either duplicate or triplicate samples. The results were expressed as mean ± SD, and P values < 0.05 were considered significant.
RESULTS
PMA stimulation of primed monocytes dramatically increased the generated oxygen-intermediate levels, which peaked approximately 6 min after reaction with lucigenin (Fig. 1A). The level of cumulative relative light units for PMA-stimulated cells was 42.2 ± 1.1 fold higher than that for unprimed control cells (P < 0.01). In comparison, cumulative relative light units generated from monocytes that were only IFN-γ primed were low and reached only 13.9% ± 4.6% of the level for PMA-stimulated cells (P < 0.02; Fig. 1B).

Oxygen-intermediate generation. (A) Serial 30-s luminescence levels from monocytes that were unprimed (▪), IFN-γ primed (▵), and PMA stimulated after priming (•). (B) Cumulative luminescence levels from monocytes. Results are mean ± SD of triplicate samples obtained from a single experiment representative of 3 separate experiments.
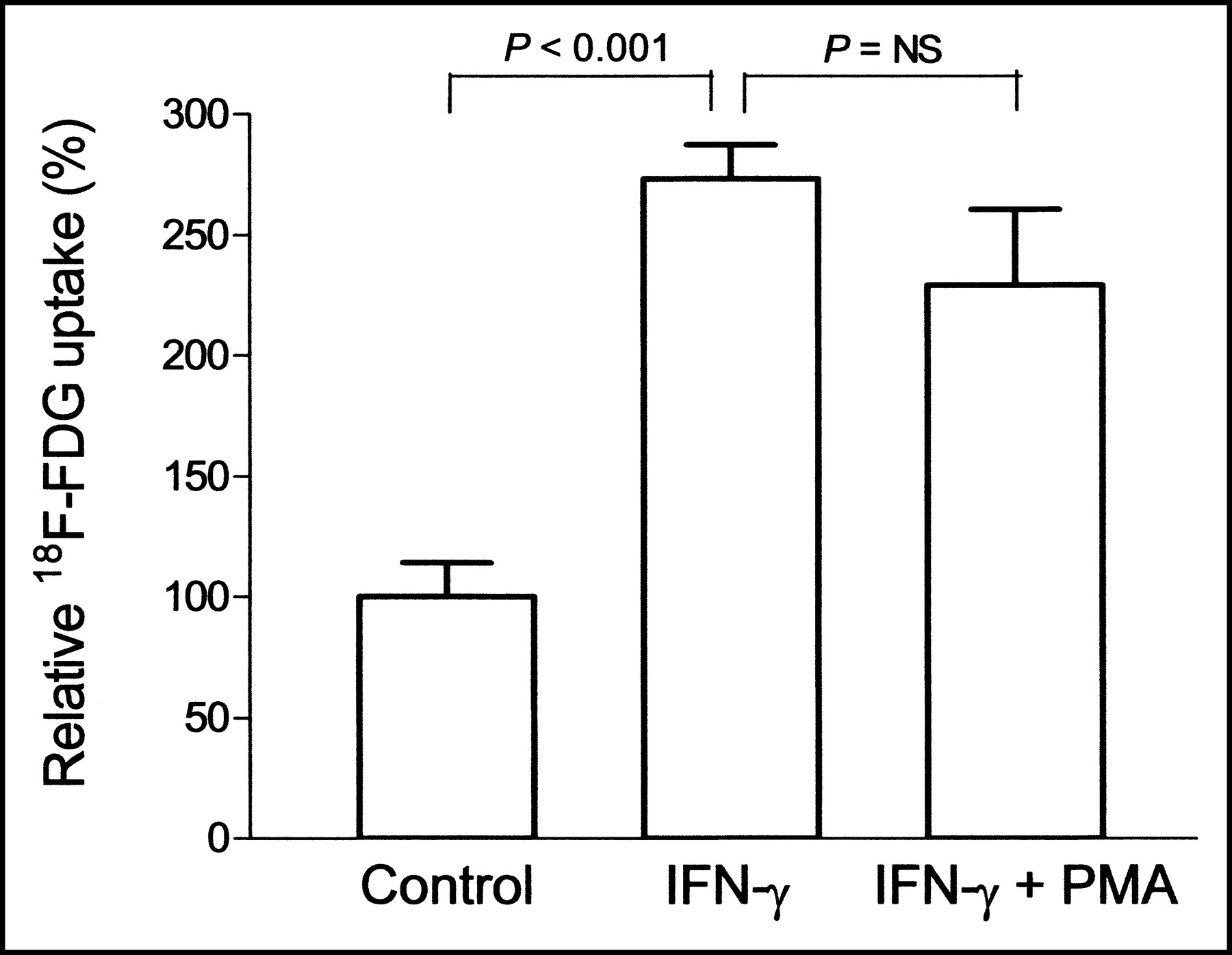
Priming with IFN-γ significantly increased 18F-FDG uptake to 273.3% ± 16.7% of the levels for unprimed control monocytes (P < 0.001). PMA stimulation of primed cells, however, did not further increase 18F-FDG uptake over the level for cells that were primed only (83.9% ± 11.5%, P = NS; Fig. 2). Cell number as assessed by the protein content of the samples showed no significant change after 3 d of treatment with IFN-γ, compared with the number for unprimed control cells (108.4% ± 14.8%; P = NS). The protein content was slightly decreased for primed cells after stimulation with 100 nmol of PMA per liter, compared with the content for unstimulated cells (24.1% ± 0.6% reduction; P < 0.001).

Relative 18F-FDG uptake in monocytes that were unprimed, IFN-γ primed, and PMA stimulated after priming. Results are expressed as percentage uptake relative to the mean of unprimed control cells. Data are mean ± SD of triplicate samples obtained from a single experiment representative of 3 separate experiments.
In the presence of either genistein or staurosporine, the priming-induced enhancement of monocytic 18F-FDG uptake was completely abolished (Fig. 3). The resultant uptake levels were substantially lower than those for cells primed in the absence of inhibitors (both P = 0.005) and were even slightly lower than those of unprimed control cells (89.6% ± 2.0% and 79.9% ± 3.8%, respectively; both P < 0.05). Under the same conditions, wortmannin and cycloheximide caused reductions in 18F-FDG uptake of only 26.7% ± 3.4% and 20.6% ± 9.5%, respectively, compared with the levels for cells primed in the absence of inhibitors (Fig. 4).

Effect of staurosporine and genistein on 18F-FDG uptake in unprimed control monocytes and IFN-γ primed monocytes. Results are percentage uptake relative to the mean of unprimed cells. Data are mean ± SD of triplicate samples from single experiment representative of 2 separate experiments. P value is for comparison with monocytes primed in absence of inhibitors.

Effect of wortmannin and cycloheximide on 18F-FDG uptake in monocytes. Results are percentage uptake relative to the mean of unprimed cells. Data are mean ± SD of triplicate samples from a single experiment representative of 2 separate experiments. P value is for comparison with unprimed control monocytes.
The design and results of our experiments are summarized in Figure 5.

Schematic representation of experimental design and results. Purified human monocytes that were activated with PMA after priming with IFN-γ showed enhanced 18F-FDG uptake and respiratory-burst activation. Priming alone did not stimulate respiratory-burst activity but was sufficient to enhance 18F-FDG uptake. This effect was completely abolished by staurosporine and genistein, inhibitors of protein kinase C (PKC) and tyrosine kinases (TyrK), respectively, whereas wortmannin, a PI3 kinase inhibitor, and cycloheximide, a protein synthesis (Psyn) inhibitor, had little effect.
DISCUSSION
The results of our study confirm that stimulation of primed monocytes with PMA, which causes a dramatic increase in oxygen-intermediate generation, is accompanied by a significant augmentation of 18F-FDG uptake compared with unprimed cells. However, it was also revealed that IFN-γ priming alone, which led to only minor levels of oxygen-intermediate generation, was sufficient to achieve a comparable if not higher level of 18F-FDG uptake. This implies that augmented 18F-FDG uptake seen in activated monocytes is driven by the cellular events associated with the process of priming rather than those that occur during development of respiratory-burst activity. Our results further provide evidence that the increase in monocytic 18F-FDG uptake induced by IFN-γ priming involves tyrosine kinases and the protein kinase C pathway but not the PI3 kinase pathway.
Priming is considered the in vitro analog of activation in vivo, in which hosts exposed to pathogens acquire resistance to subsequent infection through acquisition of aggressive phagocytes (1). IFN-γ is a well-established priming agent for studying monocyte activation states (7,8). IFN-γ-primed monocytes resemble macrophages activated in vivo and acquire a dramatic increase in responsiveness to activating agents, a critical trait for monocyte-mediated immune reactions (9). The molecular mechanisms regulating the priming effect of IFN-γ are not fully explained. It is known that IFN-γ bound to its cell-surface receptor activates the receptor-associated Janus tyrosine kinases, leading to the activation of the signal transducer and activator of transcription (STAT) pathway, which in turn induces the expression of various responsive genes (5). The wide variety of effects by IFN-γ and the complex patterns of cell-specific gene regulation preclude illumination of the precise mechanism through which IFN-γ priming increases 18F-FDG uptake in monocytes. Although glucose uptake has been related to proliferation in growth factor-stimulated macrophages (10), it appears from the lack of change in protein content after priming that the metabolic effect of IFN-γ is not coupled to proliferative response pathways.
We selected 30 min as the incubation time for 18F-FDG uptake because this time frame has frequently been used in previous in vitro experiments measuring radiolabeled deoxyglucose uptake in monocytes. In a previous study, we observed that 18F-FDG uptake in freshly purified monocytes gradually increases with incubation time but only to a modest degree—37% ± 3%—from 20 to 80 min of incubation (4). Therefore, although a lack of data using longer durations of 18F-FDG uptake (as is commonly used for 18F-FDG PET studies of clinical infection) is a limitation of our study, we believe that this factor would not have significantly affected the results of our study.
The observed augmentation in 18F-FDG uptake independent of respiratory-burst generation appears to diverge from the well-known metabolic effect of the latter. PMA stimulation of IFN-γ-primed monocytes directly activates reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, which results in a major release of superoxide anions. Such respiratory-burst activation imposes an acute metabolic demand to fuel the NADPH oxidase activity, and this demand is met by an increased use of both internal energy stores and exogenous metabolites such as glucose (11). A previous study on monocytic RAW cells confirmed an increase in 2-deoxyglucose uptake associated with high superoxide production after PMA stimulation (6). Our results also confirm the ability of PMA to stimulate respiratory-burst activity accompanied by an increase in monocytic 18F-FDG uptake, although a comparable metabolic response was found with priming alone. It may thus be that glucose uptake rates are increased in advance during the priming phase so that the high-energy demand during respiratory-burst generation can later be met. Similar to our findings, dissociation between an increase in glucose uptake and respiratory-burst activity has recently been observed for human circulating neutrophils. Priming of neutrophils with tumor necrosis factor-α caused an increase in deoxyglucose uptake identical in magnitude to that caused by N-formyl-methionyl-leucyl-phenylalanine-stimulated respiratory-burst activation, and the events were temporally dissociated (12).
The mechanisms that govern cellular glucose metabolism are diverse, and multiple signaling cascades might be involved in its regulation in different cells. In cultured skeletal muscle cells, tumor necrosis factor-α and IFN-γ have been shown to stimulate glucose transport and glucose transporter 1 protein levels, presumably through signaling pathways that induce nitric oxide production (13). We investigated the role of potential signaling cascade pathways on the regulation of monocytic glucose uptake by evaluating the effect of protein kinase inhibitors. Genistein is an inhibitor of tyrosine kinases and is used in experiments to terminate IFN-γ signaling by deactivation of STAT1 through inhibition of Janus tyrosine kinase activity (14). Staurosporine is a potent inhibitor of protein kinase C (15), which has been implicated in the regulation of glucose transport in phagocytes (16). As a result, both genistein and staurosporine completely inhibited the augmented 18F-FDG uptake response in monocytes treated with IFN-γ. These findings are similar to previous observations on the effect of these inhibitors on deoxyglucose uptake in monocytic RAW cells (6) and in human neutrophils (17).
We also investigated the PI3 kinase pathway, which is essential for generation of respiratory-burst activity (18) and is the major pathway through which insulin signals an increase of glucose use. IFN-γ induces the phosphorylation of the serine/threonine kinase Akt through PI3 kinase in human monocytes, and abrogation of this activation by PI3 kinase inhibitors has been shown to prevent some of the IFN-γ-induced effects such as monocyte adhesion (19). However, we found that wortmannin, a PI3 kinase inhibitor (20), had little effect on 18F-FDG uptake in primed monocytes. Previous studies have also demonstrated results that are consistent with our findings (6,12,17). Taken together, the results of our inhibition study suggest that IFN-γ-primed monocytes might circumvent a requirement for PI3 kinase in enhancing 18F-FDG uptake by activation of other pathways that involve tyrosine kinases or protein kinase C.
CONCLUSION
The results of our study demonstrate that priming is the cellular event responsible for the enhanced 18F-FDG uptake seen in activated monocytes and that this regulation involves both tyrosine kinases and protein kinase C activity. This study demonstrates how the functional response of monocytes may influence their level of 18F-FDG uptake and suggests that an enhanced uptake level may thus be considered a marker of cell priming.
Acknowledgments
The authors thank Jong-Ho Park for technical assistance. This work was supported by grant M20329010001-03A0723-00110 from the National Mid- and Long-Term Nuclear R&D Program of the Korean Ministry of Science and Technology.
Footnotes
Received May 19, 2003; revision accepted Sep. 25, 2003.
For correspondence or reprints contact: Kyung-Han Lee, MD, Department of Nuclear Medicine, Samsung Medical Center, 50 Ilwondong, Kangnamgu, Seoul, Korea.
E-mail: khleenm{at}samsung.co.kr
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Imaging Pulmonary Inflammation
- Prospective Evaluation of 18F-Fluorodeoxyglucose Uptake in Postischemic Myocardium by Simultaneous Positron Emission Tomography/Magnetic Resonance Imaging as a Prognostic Marker of Functional Outcome
- Oxidized Low-Density Lipoprotein Stimulates Macrophage 18F-FDG Uptake via Hypoxia-Inducible Factor-1{alpha} Activation Through Nox2-Dependent Reactive Oxygen Species Generation
- Hypoxia But Not Inflammation Augments Glucose Uptake in Human Macrophages: Implications for Imaging Atherosclerosis With 18Fluorine-Labeled 2-Deoxy-D-Glucose Positron Emission Tomography
- Imaging Atherosclerosis With F18-Fluorodeoxyglucose Positron Emission Tomography: What Are We Actually Seeing?
- Noninvasive Pulmonary [18F]-2-Fluoro-Deoxy-D-Glucose Positron Emission Tomography Correlates with Bactericidal Activity of Tuberculosis Drug Treatment
- Noninvasive Assessment of Crohn's Disease Intestinal Lesions with 18F-FDG PET/CT
- In Vitro Studies on the Signal Transduction of Thyroidal Uptake of 18F-FDG and 131I-Iodide