


Abstract
The transport of MIBG by the human norepinephrine transporter (hNET) seems to be the critical step in the treatment of MIBG-concentrating tumors. Therefore, we investigated whether the accumulation of MIBG may be induced by retroviral transfection of the hNET gene in Morris hepatoma cells. Methods: A bicistronic retroviral vector for the transfer of the hNET coding sequence and the hygromycin resistance gene was generated. Morris hepatoma cells (MH3924A) were infected with the respective retroviral particles, and hNET-expressing cell lines MHhNEThyg1 to MHhNEThyg9 were obtained through hygromycin selection. The uptake of 3H-norepinephrine or 131I-MIBG and the efflux of 131I-MIBG were determined in transfected and wild-type cells. In addition, the 131I-MIBG distribution was monitored in nude mice and rats bearing wild-type and hNET-expressing hepatomas. Results: hNET-expressing hepatoma cell lines accumulated up to 36 times more norepinephrine than did wild-type cells and 8 times more than did hNET-expressing neuroblastoma cell line SK-N-SH. The addition of nisoxetine, a selective inhibitor of noradrenaline uptake, inhibited norepinephrine uptake. Maximal 131I-MIBG accumulation was observed 2 h after incubation and was followed by 43% efflux within 4 h after the 131I-MIBG-containing medium had been removed. In vivo experiments performed with nude mice bearing both hNET-expressing and wild-type tumors showed a 10-fold-higher accumulation of 131I-MIBG in transfected tumors than in wild-type tumors. The ex vivo calculations revealed doses of 605 and 75 mGy in hNET-expressing and wild-type tumor tissues, respectively. Conclusion: Transduction of the hNET gene enables Morris hepatoma cells to accumulate norepinephrine and MIBG. However, the retention of MIBG is brief; therefore, the absorbed dose of radiation in vivo is not expected to be therapeutically effective.
Radioiodinated MIBG, a metabolically stable false analog of norepinephrine, has been widely used for imaging and targeted radiotherapy in patients with neural crest-derived tumors, such as neuroblastomas or pheochromocytomas. In the adrenal medulla and in pheochromocytomas, MIBG is stored in the chromaffin neurosecretory granules (1). The mechanism of MIBG uptake, which is qualitatively similar to that of norepinephrine, has been studied in a variety of cellular systems, and 2 different uptake systems have been postulated. Whereas most tissues accumulate MIBG by a nonspecific, nonsaturable diffusion process, cells of the neuroadrenergic tissues and malignancies derived from them exhibit active uptake of the tracer that is mediated by the noradrenaline transporter (2–4). Reverse transcription-polymerase chain reaction was used to detect the expression of the norepinephrine transporter gene and revealed a correlation between human norepinephrine transporter (hNET) expression and the uptake of MIBG in human neuroblastoma cell lines (3,5). Moreover, the differentiation of neuroblastoma cells induced by γ-interferon is associated with increased hNET gene expression followed by increased MIBG uptake (6). The clinical use of MIBG radiotherapy has been restricted so far to neural crest-derived malignancies and, because of insufficient 131I-MIBG uptake therapy in patients with these tumors, is not curative.
The present study was performed to gain more information about hNET function and the application of MIBG radiotherapy in tumor cells not related to the neural crest. For this purpose, we investigated the effect of hNET gene transduction on 131I-MIBG transport capacity and doses in rat hepatoma cells in vitro and in vivo.
MATERIALS AND METHODS
Cell Culture
Transient packaging cell line BOSC23 (7) was used for the production of ecotropic retroviral particles. Rat Morris hepatoma cell line MH3924A and human neuroblastoma cell line SK-N-SH, expressing the norepinephrine transporter, and the BOSC23 cell line were cultured in RPMI 1640 medium (Gibco BRL) supplemented with 292 mg of glutamine per milliliter, 100,000 IU of penicillin per liter, 100 mg of streptomycin per liter, and 10% (BOSC23 and SK-N-SH) or 20% (MH3924A) fetal calf serum. The cell lines were cultured at 37°C in 5% CO2.
Cloning and Expression of hNET Gene and Hygromycin Gene
For gene transfection, a bicistronic retroviral vector based on Moloney murine leukemia virus (MMLV) M48 (obtained from O. Danos (8)) was constructed: The hNET coding sequence (obtained from S. Amara (9)) and the hygromycin resistance gene, which was taken from pIRES1hyg (Clontech), were cloned downstream of the elongation factor 1α (EF1α) promoter, taken from the pShooter vector pEF/myc/cyto (Invitrogen) (Fig. 1). The simultaneous expression of both the hNET coding sequence and the hygromycin resistance gene and stabilization of the respective messenger RNAs were ensured by insertion of a synthetic Intron (Schering Corp.) and the encephalomyocarditis virus (EMCV) internal ribosomal entry site between the genes.

Structure of recombinant retroviral vector based on MMLV. Vector is designed to transfer hNET and hygromycin resistance (hyg) activity. Coexpression is achieved by use of internal ribosomal entry site (IRES) from EMCV. Synthetic Intron (IVS) is inserted to stabilize messenger RNA. Gene expression is regulated by EF1α promoter. LTR = long terminal repeat.
Retroviral Infection and Generation of Recombinant Cell Lines
For transient packaging of the recombinant retroviral vector, DNA was introduced into BOSC23 cells by use of Lipofectamine (Invitrogen). After 2 d, the culture medium was centrifuged to remove detached BOSC23 cells, and the supernatant was used for infection of MH3924A cells in the presence of 8 μg of Polybrene (Abbott Laboratories) per milliliter overnight. For the generation of stable cell lines, the infected cells were treated with 425 μg of hygromycin per milliliter for 4 wk, and hNET expression was determined by Northern blot analysis. Briefly, total cellular RNA was isolated by the acid guanidinium thiocyanate-phenol-chloroform method, and 5 μg of RNA were electrophoresed in 1% agarose-formaldehyde gels and blotted on nylon membranes (Hybond N; Amersham) in 20× SSC (0.3 mol/L sodium citrate, 3 mol/L NaCl pH 7.0) buffer. The hybridization of 32P-deoxycytidine triphosphate-labeled complementary DNA probes corresponding to either the hNET or the β-actin coding sequence was performed by the method described by Church and Gilbert (10). Autoradiography was done with Kodak X-OMAT TM AR films (Eastman Kodak Co.) for 4 h (β-actin) or 24 h (hNET).
Determination of 3H-Noradrenaline Uptake
The uptake of noradrenaline was determined in wild-type and hNET-expressing MH3924A (MHhNEThyg) cell lines as well as in human neuroblastoma cell line SK-N-SH by a modification of the method of Lingen et al. (11). Briefly, after being washed with phosphate-buffered saline (PBS), the cells were cultured for 15 min in preincubation buffer (125 mmol of NaCl per liter, 2.4 mmol of K2SO4 per liter, 1.2 mmol of KH2PO4 per liter, 1.2 mmol of MgSO4 per liter, 25 mmol of N-(2-hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) [HEPES]-Tris [pH 7.4] per liter, 5.6 mmol of d-(+)-glucose per liter, 1 mmol of ascorbic acid per liter, 10 μmol of pargyline [inhibitor of monoamine oxidase] per liter, 0.02 μmol of 3,5-dinitrocatechol [inhibitor of catechol-O-methyltransferase; OR486; RBI] per liter). Subsequently, the cells were incubated for 10 min in uptake buffer (125 mmol of NaCl per liter, 2.4 mmol of K2SO4 per liter, 1.2 mmol of KH2PO4 per liter, 1.2 mmol of MgSO4 per liter, 25 mmol of HEPES-Tris [pH 7.4] per liter, 5.6 mmol of d-(+)-glucose per liter, 1 mmol of ascorbic acid per liter, 10 μmol of pargyline per liter) containing 74 kBq of 3H-noradrenaline (specific activity, 1.92 TBq/mmol; radiochemical purity, 98.5%; Amersham) and cold noradrenaline at a final concentration of 10 nmol/L in the presence or absence of 10 μmol of nisoxetine, a selective inhibitor of noradrenaline uptake, per liter (11,12). After incubation, the cells were washed 3 times with ice-cold PBS and lysed with 500 μL of NaOH (0.3 mol/L).
Measurement of 131I-MIBG Uptake and Efflux
For the evaluation of MIBG uptake, MH3924A cells and the MHhNEThyg2 cell line were incubated in 1 mL of culture medium supplemented with 74 kBq of 131I-MIBG (specific activity, 591.4 TBq/mmol; Amersham) per well in 6-well plates for 1, 2, 5, 10, 30, 120, 240, or 480 min. Because MIBG is metabolically stable, no inhibitors of monoamine oxidase and catechol-O-methyltransferase were added. The cells were washed twice with ice-cold PBS and lysed with 500 μL of NaOH (0.3 mol/L) on ice. For the determination of 131I-MIBG efflux, MHhNEThyg2 and MH3924A cells were cultured for 1 h in medium containing 74 kBq of 131I-MIBG in 6-well plates. After the plates were washed twice with PBS, the cells were lysed as described above or fresh nonradioactive medium was added; after 5, 10, 30, 60, 120, or 240 min, the radioactivity in cell lysates and in the medium was measured by use of an automated NaI(Tl) well counter (Cobra II; Canberra Packard). The viable cell number was determined in parallel in different wells by use of a Coulter Counter (Coulter Electronics) and by trypan blue staining (more than 94% viable cells). The median cell volume was determined by use of the Coulter Counter.
131I-MIBG Uptake in Tumor Tissue
Experiments for evaluating 131I-MIBG uptake in tumor tissue were performed in compliance with German laws relating to the conduct of animal experimentation. A total of 4 × 106 MHhNEThyg2 or MH3924A cells were transplanted subcutaneously into the right (MHhNEThyg2) or left (MH3924A) thigh of young male ACI rats weighing 200–250 g or Swiss CD1 nu/nu mice (Charles River Laboratories; delivered by Iffa Credo) weighing 20–25 g. For imaging studies, which were performed under general gaseous anesthesia (40% O2–60% N2O–1% halothane), only animals bearing tumors with a minimum diameter of 15 mm were selected. Immediately after injection of 200 μL of 131I in 0.9% NaCl (14.8 MBq for mice and 37 MBq for rats) into the tail vein of the animals, a dynamic scintigraphic image was obtained over 30 min to evaluate the radioactivity in the blood circulation. The general body distribution and the early uptake into tumor tissues of 131I-MIBG were evaluated by use of a 25.4-cm (10-in.) scintillation camera (Searle-Siemens). Additional images were obtained 1, 2, 16, and 24 h after injection.
The absolute amount of radioactivity (percentage of injected dose per gram of wet tissue) was determined in 3 rats at 4 h after injection and in 20 mice at 2, 4, 16, 24, and 48 h after injection by analyzing the organs by use of an automated NaI(Tl) well counter. Dosimetric calculations were done with MIRDOSE3* (13) by assuming a unit-density sphere with a mass of 1 g and considering only the value of the absorbed dose in the sphere. Therefore, the sum of 2 exponential decay functions was fitted to the data.
RESULTS
Generation and Functional Characterization of hNET-Expressing MH-Series Cell Lines
MH3924A cells were infected with recombinant retroviruses (Fig. 1), and 9 stable hNET-expressing cell lines (MHhNEThyg1 to MHhNEThyg9) were established by hygromycin selection. Northern blot analysis revealed transcription of the hNET gene in all cell lines (data not shown). The transport activity of the hNET gene product in the MHhNEThyg cell lines was investigated by noradrenaline uptake experiments (Fig. 2). The MHhNEThyg cell lines accumulated significantly more radiotracer than did the MH3924A cell line or the SK-N-SH positive control cell line. The noradrenaline uptake of the SK-N-SH cells was 5-fold higher than that of the MH3924A cells. Between 5- and 40-fold more noradrenaline was transported into the MHhNEThyg cell lines than into the wild-type counterparts. Uptake experiments carried out in the presence of nisoxetine showed a decrease in noradrenaline accumulation to the level in wild-type cells, indicating that the hNET gene product was functionally active. For further experiments, the MHhNEThyg2 cell line, characterized by a reproducible high level of substrate uptake, was used. The in vitro doubling times were calculated to be 15.2 ± 0.6 h (mean ± SD) for the MH3924A cells and 15.8 ± 0.2 h for the MHhNEThyg2 cells.

3H-Noradrenaline uptake in SK-N-SH neuroblastoma cells, wild-type Morris hepatoma cells, and hNET-expressing Morris hepatoma cells after 10 min of incubation with 3H-noradrenaline in presence or absence of nisoxetine. Values are means ± SDs (n = 3).
131I-MIBG Uptake and Efflux in MHhNEThyg2 Cell Line
MHhNEThyg2 cells and MH3924A cells were incubated in 131I-MIBG-containing medium for different periods of time. A significant accumulation of the radiotracer by the MHhNEThyg2 cells as a function of incubation time was observed (Fig. 3), with up to 36-fold-higher 131I-MIBG uptake in the transfected cells than in the wild-type cells. The maximal uptake of 131I-MIBG by the MHhNEThyg2 cells was measured after 2 h of incubation, with a plateau level implicating steady-state uptake. At that time, the intracellular radioactivity in 105 recombinant cells was calculated to be 12.2% of the total radioactivity in cell lysates and in the medium. Given that the median cell volumes of MH3924A cells and MHhNEThyg2 cells were 1.4 fL (range, 1.02–2.6) and 1.2 fL (range, 0.91–2.0), respectively, the intracellular 131I-MIBG concentration was calculated to be 1.3 × 106-fold higher than the radioactivity in the medium for the MHhNEThyg2 cells.
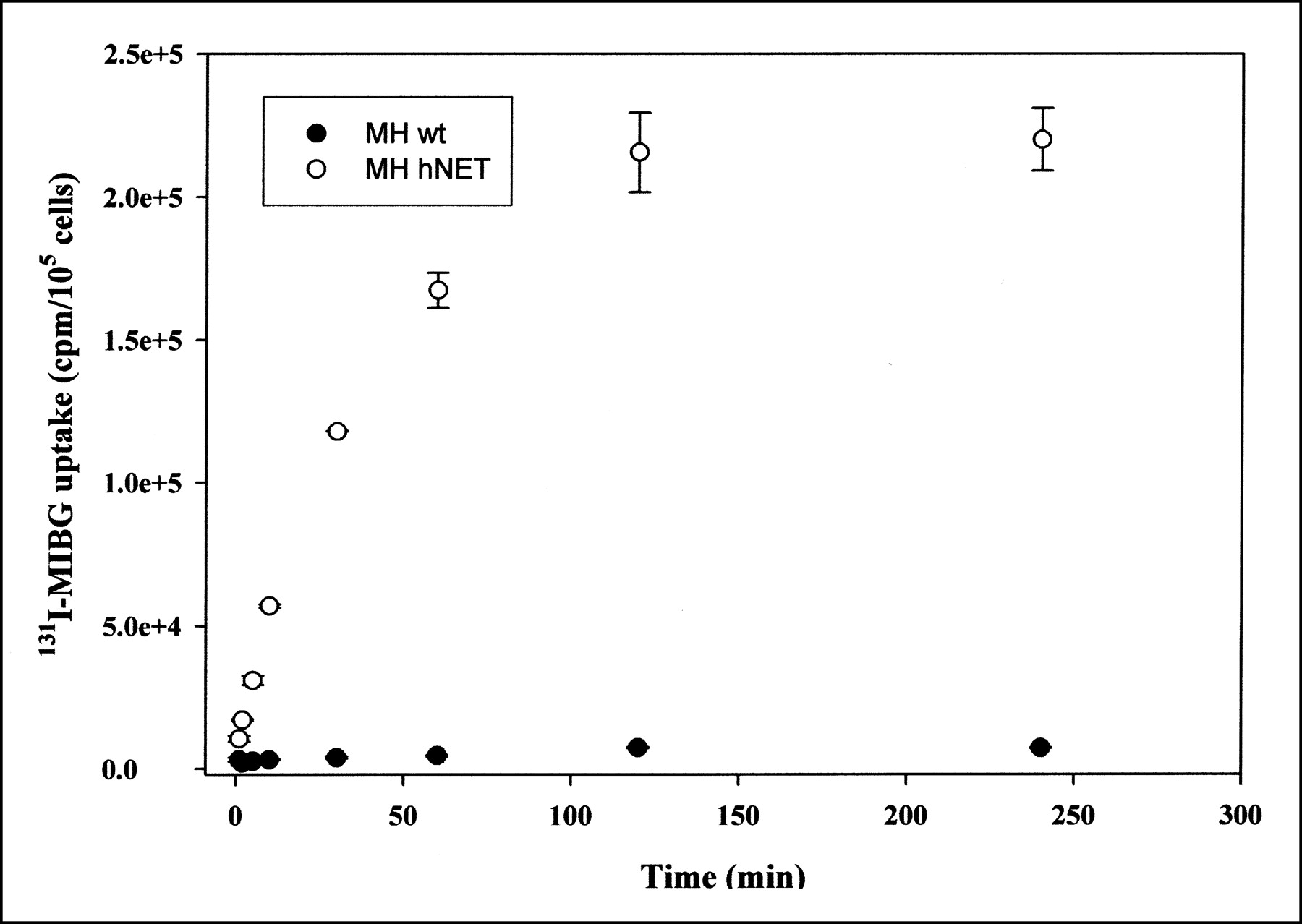
Time dependence of 131I-MIBG uptake in wild-type (wt) Morris hepatoma cells and hNET-expressing cell line MHhNETShyg2. Values are means ± SDs (n = 3).
For investigation of the efflux of the radiotracer, the uptake of 131I-MIBG by the MHhNEThyg2 and MH3924A cells was allowed to proceed for 1 h before the medium was replaced with nonradioactive medium. Thereafter, the amount of radioactivity present in the cell lysates was determined as a function of time, as shown in Figure 4. After 1 h, MHhNEThyg2 cells released about 38% of the cellular radioactivity into the medium, and 43% efflux was observed after 4 h, indicating a slow effusion of the radiotracer from these cells. From the MH3924A cells, 75% and 95% of the radioactivity was released after 1 h and after 4 h, respectively.

MIBG efflux from hNET-expressing cells after 1 h of incubation with 131I-MIBG. Values are means ± SDs (n = 3).
131I-MIBG Uptake and Efflux in Tumor-Bearing Animals
For investigation of the function of the hNET gene product in vivo, 131I-MIBG imaging studies of nude mice and ACI rats transplanted with both MHhNEThyg2cells and MH3924A cells were performed.
In scintigraphic images of mice, clear 131I-MIBG uptake in the hNET-expressing tumors from 1 h to 23 h after injection was accompanied by a high background in the abdomen (Fig. 5). The ex vivo quantification of radioactivity revealed maximal accumulation of 131I-MIBG in the hNET-expressing tumors 2 h after administration of the tracer. Increased activities were also observed in the liver, the spleen, the kidneys, and the bowels. Compared with the wild-type tumors, the hNET-expressing tumors showed a 10-fold-higher accumulation of 131I-MIBG over 48 h (Table 1). Figure 6 shows efflux of the radiotracer similar to that observed in the respective cell culture experiment. After the application of 14.8 MBq of 131I-MIBG, dosimetry revealed absorbed doses of 605 mGy in the hNET-expressing tumors and 75 mGy in the wild-type tumors. In most of the nontumor tissues investigated, even higher absorbed doses were observed (spleen, 399 mGy; liver, 741 mGy; kidneys, 1,655 mGy; bowels, 802 mGy). With respect to 131I-MIBG uptake in tumor-bearing rats (n = 6), no significant difference between the hNET-expressing tumors and the wild-type tumors was revealed by scintigraphy, even at 24 h after injection (Fig. 7). The radioactivities calculated ex vivo 4 h after injection (n = 3) were 22.02 ± 16 kBq/g of wet tissue in transfected tumors and 8.1 ± 5.9 kBq/g of wet tissue in wild-type tumors.

Scintigraphic images of tumor-bearing young male nu/nu mice subcutaneously transplanted with hNET-expressing cells (right thigh) or wild-type Morris hepatoma cells (left thigh) at 30 min (A), 1 h (B), 2 h (C), and 24 h (D) after injection of 131I-MIBG.
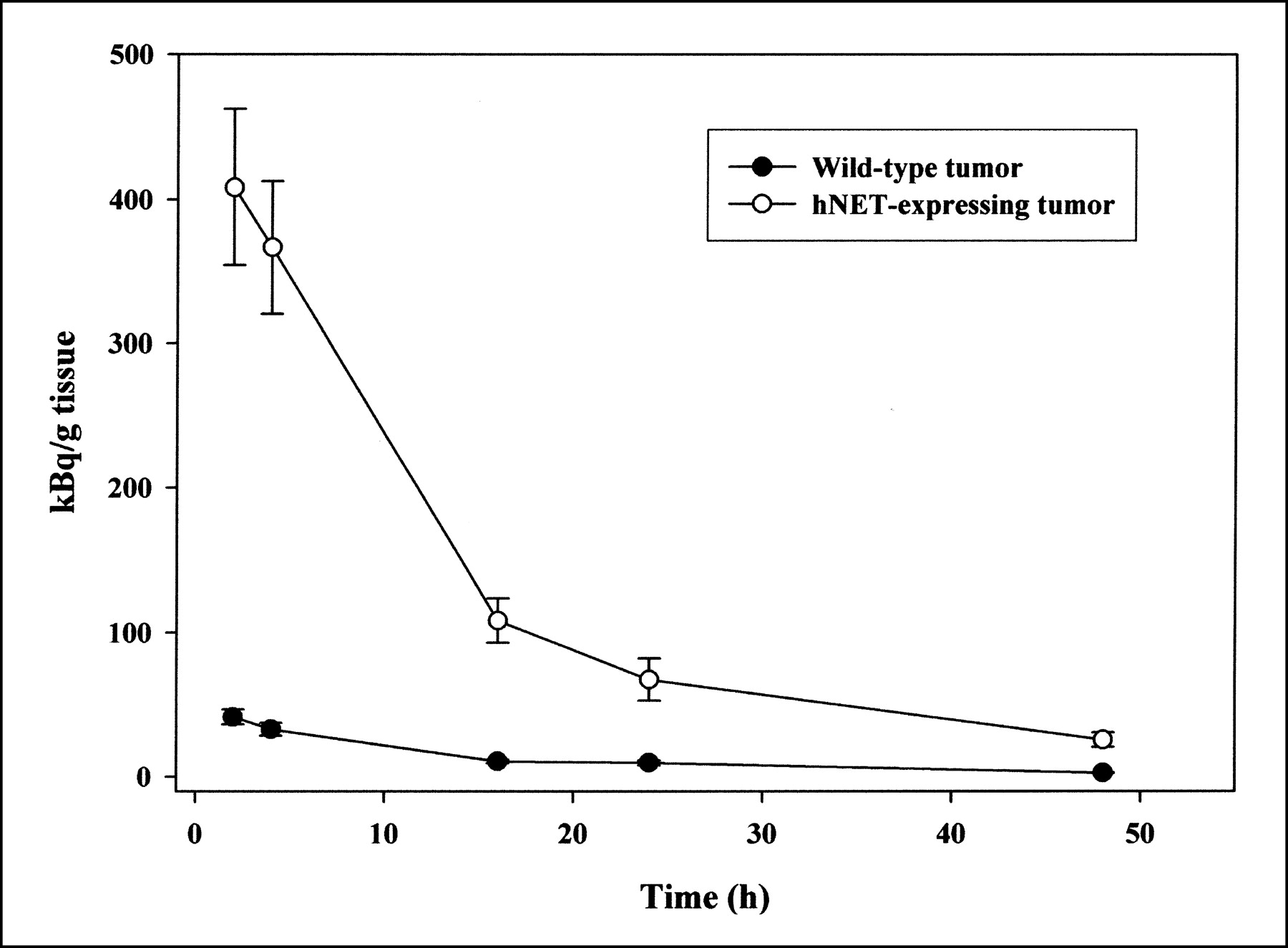
MIBG uptake in tumor tissue at different times after 131I-MIBG administration in nu/nu mice. Values are means ± SDs (n = 4).
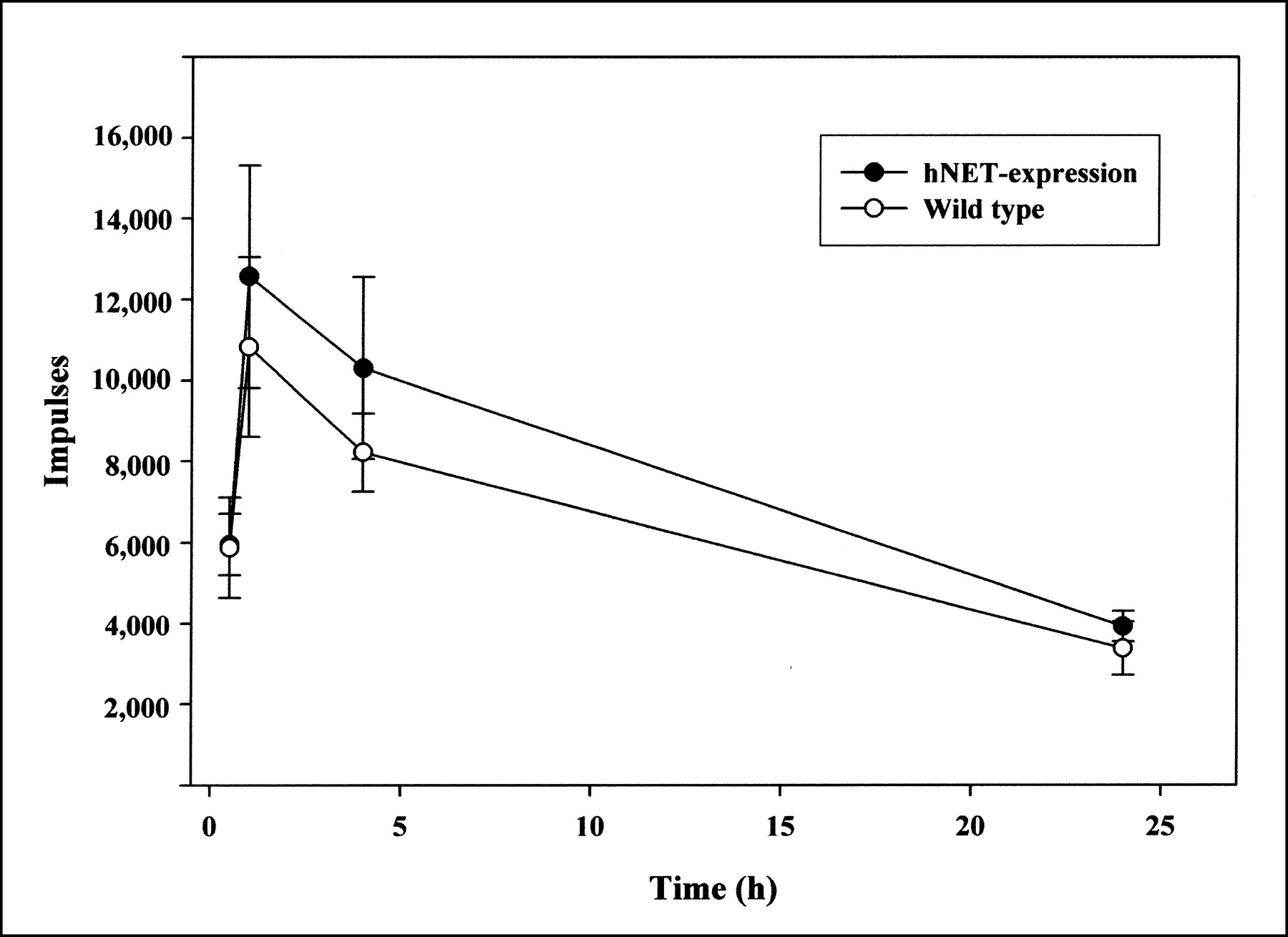
131I-MIBG uptake (impulses) in hNET-expressing and wild-type Morris hepatoma cells at different times after radiotracer application in rats. Values are means ± SDs (n = 6).
Radioactivity in Various Tissues at 2-48 Hours After Injection of 131I-MIBG
DISCUSSION
The clinical value of 131I-MIGB for the therapy of cancer patients is limited to neural crest-derived tumors expressing hNET. These tumors accumulate the radiotracer, but often a rather low radiation dose is achieved. In the present study, we have shown that transduction of the hNET coding sequence into rat Morris hepatoma cells induces the expression of a functional norepinephrine transporter, leading to the active uptake of 3H-noradrenaline and 131I-MIBG. The effect of hNET gene transfection was investigated previously with COS-1 cells, HeLa cells, and glioblastoma cells, and 3-fold (COS-1 cells), 9-fold (HeLa cells), and 15-fold (glioblastoma cells) increases in 131I-MIBG accumulation were achieved (4,9,14). The data that we obtained from in vitro experiments indicated up to 36-fold-higher MIBG uptake in hepatoma cells transfected with the hNET gene. The scintigraphic images of nude mice bearing both wild-type and hNET-expressing Morris hepatoma cells confirmed the in vitro data, revealing a significant accumulation of 131I-MIBG in the genetically modified tumors. The in vivo results obtained from rats transplanted with hNET-expressing tumors, however, were inconsistent with those obtained from nude mice. The transport of MIBG has been described to be influenced by a variety of processes and factors, such as N-glycosylation of the transport protein, perfusion, and competition of the substrate with naturally occurring catecholamines (15–22). Given that plasma catecholamine levels are even higher in mice than in rats, competition seems not to be the reason for the lack of MIBG accumulation in rats (23–25). The production of inactivating antibodies against the hNET gene product in rats receiving transplants but not in immune-deficient nude mice is possible. As a result, the functional activity of the transport molecule and, consequently, MIBG uptake could be blocked (26).
Compared with previous studies concerning the efflux of 131I from MH-series cells expressing the human sodium iodide symporter (27), this study revealed longer retention of MIBG in hNET-transfected cells. Nevertheless, at 4 h after incubation with MIBG, efflux of 43% of the radioactivity was determined for the recombinant cells, whereas the wild-type cells had lost 95% of the radioactivity. With regard to MIBG radiotherapy for nonneuroectodermal tumors, intracellular trapping of the tracer is required to achieve therapeutically sufficient doses of radioactivity in genetically modified tumor cells. A positive correlation has been observed between the content of chromaffin neurosecretory granules and the uptake of radiolabeled MIBG (28).
Human glioblastoma cells transfected with the bovine NET gene were killed by doses of 0.5–1 MBq of 131I-MIBG per milliliter in monolayer cell cultures as well as in spheroids (14). Accordingly, the authors expected the intratumoral activity in a 70-kg patient to be 0.021%. This value corresponds to the range of MIBG uptake usually achieved in neuroblastoma cells. However, data obtained from in vitro experiments cannot be applied to the in vivo situation. In contrast to what is achieved under stable in vitro conditions, radioactive doses delivered to a tumor in vivo differ because of decreasing radioactivity in the serum and heterogeneity within the tumor tissue. For calculation of the radiation dose in a particular tumor more precisely, we propose in vivo dosimetry. Using 14.8 MBq of 131I-MIBG for tumor-bearing mice (corresponding to 2,200 MBq/m2 in humans), we calculated radiation doses of 605 mGy for hNET-expressing tumors and 75 mGy for wild-type tumors. With regard to the treatment of patients with nonneuroectodermal tumors transfected with the hNET gene, we consider the absorbed dose to be too low to lead to any tumor response. In addition, the in vivo experiments presented here were performed with animals that had been transplanted with 100% stable hNET-expressing cells. Therefore, because of the low in vivo infection efficiency of viral particles, infection of tumor cells in vivo will result in even lower radiation doses.
The basal efflux of 131I-MIBG from human neuroblastoma cells is highly temperature dependent and is inducible by high potassium concentrations (in a calcium-dependent manner), noradrenaline, cold MIBG, and imipramine (29). Ca2+-independent release, which involves the cytoplasmic neurotransmitter pool, is mediated by reverse action of the transport system and has been described for tissues lacking storage granules. For different neurotransmitter transport systems, carrier-mediated release has been described as being triggered by depolarization and transstimulation by high ligand concentrations (29–34). Because no additive effects were observed when the different stimuli were combined and because the stimulation of MIBG release could be inhibited by the competitive inhibitor imipramine, it was suggested that the uptake system works in reverse mode for efflux (29,35,36). Exocytosis is assumed to play a minor role, and it has been suggested that carrier-mediated efflux occurs at low intracellular concentrations and that passive diffusion occurs at high intracellular concentrations (35,36). Pharmacologic modulation of MIBG retention by inhibition of efflux through calcium channel blockage by nifedipine may not be successful in tumors without granules, such as neuroblastomas, or in other tumors, such as the hepatomas used in this study (37). As an alternative, transport inhibitors, such as imipramine, may be used to prolong intracellular MIBG accumulation. However, for human neuroblastoma cells, Lashford et al. observed that 66% of the radioactivity remained within the cells after 24 h, whereas after the addition of desipramine, the intracellular radioactivity decreased to 10% (38). These results were interpreted as evidence that MIBG is lost rapidly and taken up again. The levels of competing plasma catecholamines may be reduced by use of inhibitors of biosynthetic enzymes, such as tyrosine hydroxylase or dopamine β-hydroxylase, but no data are available at this time. Therefore, pharmacologic modulation may be difficult. In addition, there has been speculation about whether the transfer of the hNET gene into pheochromocytomas or neuroblastomas may enhance the efficiency of MIBG therapy (14). Future developments may include pharmacologic modulation of MIBG retention, interaction with competing catecholamines, or the use of the hNET coding sequence as an in vivo reporter gene.
CONCLUSION
The transfer of the hNET gene into rat Morris hepatoma cells resulted in enhanced uptake of MIBG in vitro and in vivo. We also showed considerable efflux of the radiotracer from the cells, resulting in a low absorbed dose in the tumor. Future experiments should address the question of how to achieve longer retention of MIBG in recombinant tumors to provide a therapeutically sufficient radiation dose. The use of the recombinant hNET gene product as an in vivo reporter, as has been proposed for the human sodium iodide symporter (27,39), is not promising, because images showed a high background and a relatively faint appearance of the genetically modified tumor.
Acknowledgments
The authors thank Rainer Kühnlein for his help in performing the animal experiments. This study was supported by a grant from the Tumorzentrum Heidelberg/Mannheim.
Footnotes
Received Oct. 24, 2002; revision accepted Feb. 13, 2003.
For correspondence or reprints contact: Annette Altmann, PhD, Clinical Cooperation Unit Nuclear Medicine, German Cancer Research Center, Im Neuenheimer Feld 280, D-69120 Heidelberg, Germany.
E-mail: A.Altmann{at}dkfz-heidelberg.de
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Norepinephrine Transporter as a Target for Imaging and Therapy
- Dynamic imaging for CAR-T-cell therapy
- Noninvasive Molecular Imaging Using Reporter Genes
- Engineered Antibody Fragments with Infinite Affinity as Reporter Genes for PET Imaging
- Molecular-Genetic Imaging Based on Reporter Gene Expression
- Comparison of Uptake of Multiple Clinical Radiotracers into Brown Adipose Tissue Under Cold-Stimulated and Nonstimulated Conditions
- Imaging hNET Reporter Gene Expression with 124I-MIBG
- Radioiodine Therapy of Hepatoma Using Targeted Transfer of the Human Sodium/Iodide Symporter Gene
- Characteristics and Regulation of 123I-MIBG Transport in Cultured Pulmonary Endothelial Cells
- The Human Norepinephrine Transporter in Combination with 11C-m-Hydroxyephedrine as a Reporter Gene/Reporter Probe for PET of Gene Therapy