


Abstract
123I-Metaiodobenzylguanidine (123I-MIBG) is used for lung scintigraphy to assess pulmonary endothelial cell integrity, but its processing at the cellular level has not been investigated to date. We thus characterized the mechanisms that mediate 123I-MIBG transport in pulmonary endothelial cells and investigated the effects of stimuli associated with pulmonary dysfunction. Methods: Calf pulmonary artery endothelial (CPAE) cells were examined for 123I-MIBG uptake and efflux rates and evaluated for the presence of norepinephrine (NE) transporters by Western blotting. The specificity of 123I-MIBG uptake was investigated with inhibitors of the uptake 1 and uptake 2 transport systems. In addition, we tested the effects of hypoxia (1% O2), phorbol 12-myristate 13-acetate (PMA, a protein kinase C [PKC] activator), and NG-nitro-l-arginine methyl ester (l-NAME) (a nitric oxide synthase inhibitor) treatments on CPAE cell 123I-MIBG uptake. Results: CPAE cells demonstrated a time-dependent increase in 123I-MIBG uptake that reached a relative plateau (mean ± SD) at 4 h of 375.6% ± 5.9% the 30-min level. When the culture medium was changed after 30 min of uptake, 123I-MIBG gradually was eluted from the cells at an efflux rate of 43.8% over 2 h. Western blotting confirmed the presence of NE transporters in CPAE cells. The uptake 1 inhibitors desipramine, imipramine, and phenoxybenzamine at 50 μmol/L reduced 123I-MIBG uptake to 55.3% ± 2.7%, 62.4% ± 3.5%, and 48.0% ± 2.2% control levels, respectively, whereas none of the uptake 2 inhibitors had an effect. Exposure to hypoxia resulted in a reduction in 123I-MIBG uptake to 77.5% ± 0.2% and 50.0% ± 3.4% control levels at 0.5 and 4 h, respectively. PMA (10 ng/mL) and l-NAME (2 nmol/L) decreased 123I-MIBG uptake to 76.7% ± 9.0% and 86.5% ± 5.6% control levels, respectively. Conclusion: Pulmonary endothelial cells express NE transporters and actively take up 123I-MIBG through the specific uptake 1 system. Furthermore, 123I-MIBG transport can be reduced by hypoxia, PKC activation, and nitric oxide deficiency, which may contribute partly to the lower levels of lung uptake observed in diseases that compromise pulmonary endothelial cell integrity.
Metaiodobenzylguanidine (MIBG) is a guanethidine analog that shares structural features and biologic behavior with the adrenergic neurotransmitter norepinephrine (NE). Scintigraphy with 123I-MIBG is widely used for the clinical evaluation of neuroendocrine tumors and myocardial sympathetic dysfunction (1,2), and 131I-MIBG is used for the treatment of neural crest tumors (1). 123I-MIBG imaging also is applied for the assessment of pulmonary integrity in patients with various forms of lung injuries, including chronic obstructive pulmonary disease, lung fibrosis, vasculitis, irradiation, high-altitude pulmonary edema, and chemotherapeutic lung toxicity (3–8).
The pattern of 123I-MIBG uptake by the lungs provides information on pulmonary integrity because it reflects the ability to clear circulating NE to help maintain the homeostasis of circulating levels (9–11). Investigations of this metabolic lung function have shown that NE clearance takes place in pulmonary endothelial tissues (12,13) and that it is mediated through a saturable, energy-dependent, imipramine-sensitive carrier mechanism (14). Studies with isolated animal lung models suggested that 123I-MIBG uptake may occur through a similar process. Subsequent clinical scintigraphic imaging studies demonstrated altered processing of 123I-MIBG by the lungs in patients with diverse forms of pulmonary injuries. Therefore, it is hoped that clinical 123I-MIBG scintigraphy will provide useful information regarding the functional status of the pulmonary endothelium, but accurate interpretation of such imaging studies will require a thorough understanding of the mechanism and regulation of tracer accumulation. To date, however, the properties of pulmonary endothelial cell 123I-MIBG transport have not been elucidated at the cellular level. Therefore, in the present study, we investigated the characteristics of 123I-MIBG transport in calf pulmonary artery endothelial (CPAE) cells and further evaluated the effects of stimuli that have been implicated in pulmonary endothelial cell dysfunction.
MATERIALS AND METHODS
Cell Culture Conditions
CPAE cells were maintained in RPMI 1640 medium (Gibco BRL) supplemented with 20% fetal bovine serum and penicillin–streptomycin (100 U/mL) at 37°C in a 5% CO2 incubator. The culture medium was changed every 3–4 d, and 123I-MIBG uptake experiments and Western blotting were performed when cell confluence reached 80%. Human neuroblastoma SK-N-SH cells cultured in minimum essential medium (Gibco BRL) supplemented with 10% fetal bovine serum and penicillin–streptomycin (100 U/mL) were used as positive control cells for Western blot analysis of NE transporters.
Cellular 123I-MIBG Uptake and Washout Rate Measurements
123I-MIBG uptake experiments were performed by adding 74 kBq (20 mCi/mL) of 123I-MIBG (Korean Atomic Energy Research Institute) to cells in 12-well plates containing 1 mL of culture medium. The specific activity of 123I-MIBG preparations used in the cell experiments was 5–11 GBq/mg, and there was negligible 123I-MIBG dehalogenation, as indicated by quality assurance tests. After incubation at 37°C in 5% CO2 for various times up to 8 h, cells were rapidly washed twice with cold phosphate-buffered saline (PBS) and measured for radioactive counts with a high-energy γ-counter (Wallac). To measure 123I-MIBG washout rates, cells were incubated with 123I-MIBG for 30 min, after which the culture medium was removed and replaced with the same volume of fresh medium. After further incubation for various times up to 8 h, cells were rapidly washed with cold PBS and measured for radioactivity as described above. All cell uptake results were corrected for protein content, as determined by the Bradford method, and expressed as mean ± SD (triplicate samples) of percent uptake relative to that of control cells.
Western Blot Analysis for NE Transporter Expression
CPAE and SK-N-SH cells grown in 150-mm culture plates were washed twice with cold PBS and solubilized in 1 mL of cold radioimmunoprecipitation buffer (Tris at 65 mmol/L [pH 7.4], NaCl at 150 mmol/L, ethylenediaminetetraacetic acid at 100 mmol/L, 10% Nonidet P-40, and 10% sodium deoxycholate) supplemented with protease inhibitors (aprotinin, leupeptin, and pepstatin each at 1 μg/mL and phenylmethylsulfonyl fluoride at 500 μmol/L) for 15 min at 4°C with constant shaking. Lysates were centrifuged at 14,000g for 15 min at 4°C, and the supernatants were appropriately diluted after protein assays (Bio-Rad). Samples (500 μL) were incubated with 5 μL of a monoclonal antibody directed against NE transporter proteins (antibody NET17-1; Mab Technologies, Inc.) for 2 h at 4°C and then mixed with 100 μL of protein A–Sepharose CL-4B beads (Sigma) overnight at 4°C. Samples were washed 3 times with cold PBS for 5 s each time, eluted into 50 μL of 2× Laemmli sample buffer (Sigma) for 30 min at room temperature, and heated for 1 min at 90°C. Supernatants then were subjected to Western blotting by separation on a sodium dodecyl sulfate–10% polyacrylamide gel and transfer to a polyvinylidene difluoride membrane (Bio-Rad). The membrane was incubated with antibody NET17-1 at a dilution of 1:1,000 and then incubated with an antimouse horseradish peroxidase–conjugated secondary antibody (Amersham Bioscience) at a dilution of 1:2,000. NE transporter proteins were visualized with detection reagents and high-performance chemiluminescence films.
Effects of Uptake 1 and Uptake 2 Transport Inhibitors on 123I-MIBG Uptake
The effect of blocking uptake 1–mediated transport on 123I-MIBG uptake was evaluated by use of a 30-min pretreatment of CPAE cells with concentrations of 0, 1, 50, or 100 μmol/L of the specific uptake 1 inhibitor desipramine (Sigma) or imipramine (Sigma) or of phenoxybenzamine (Sigma), a nonselective adrenergic blocking agent with uptake 1 inhibitory action. The effect of blocking uptake 2–mediated transport was evaluated by use of a 30-min pretreatment of cells with the well-known uptake 2 inhibitor corticosterone, clonidine, or normetanephrine (Sigma) at 0, 5, or 50 μmol/L. Cellular 123I-MIBG uptake levels were measured after 30 min of incubation at 37°C as described above.
Effects of Hypoxia, Phorbol 12-Myristate 13-Acetate (PMA), and NG-Nitro-l-Arginine Methyl Ester (l-NAME) on Cellular 123I-MIBG Uptake
To evaluate the effect of hypoxia on 123I-MIBG transport, CPAE cells were exposed to hypoxic conditions for 0.5, 1, or 4 h by placement in an anaerobic chamber containing 1% O2, 5% CO2, and 94% N2 (Anaerobic System-1029; Forma Scientific). Cells were incubated with 74 kBq of 123I-MIBG inside the hypoxic chamber for 30 min, after which they were rapidly washed with cold PBS, lysed, and measured for radiouptake as described above. Cells incubated under standard oxygen conditions (95% air, 5% CO2) were used as controls. The effects of PMA (5 or 10 nmol/L) and l-NAME (10 μmol/L and 2 mmol/L) were evaluated by pretreatment of cells with the respective agents for 16 h before 30 min of incubation with 74 kBq of 123I-MIBG and radiouptake measurements as described above.
RESULTS
CPAE Cell 123I-MIBG Uptake and Washout Kinetics
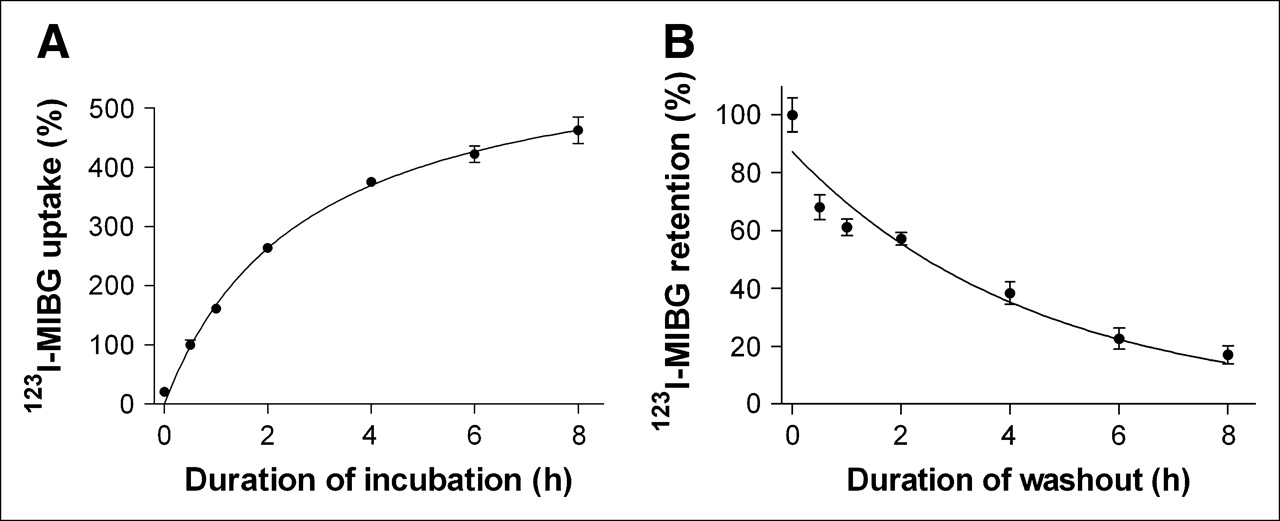
CPAE cells incubated with 123I-MIBG demonstrated a gradual increase in radiouptake over time. Uptake after 1 and 2 h of incubation increased to 161.3% ± 4.7% and 263.7% ± 7.3% that at 30 min, respectively, and a relative uptake plateau of 375.6% ± 5.9% was reached at 4 h (Fig. 1A). When the culture medium was changed after 30 min of 123I-MIBG incubation, radioactivity initially taken up by CPAE cells was gradually released into the fresh medium over time in an exponential manner. As a result, 1-, 2-, and 4-h retention rates were 61.1% ± 1.9%, 57.2% ± 2.2%, and 38.3% ± 3.9%, respectively (Fig. 1B). The washout curve was analyzed with Prism version 3.02 software (GraphPad Software Inc.). Nonlinear regression curve fitting with 1-phase exponential decay (goodness of fit, R2 = 0.9268) revealed a retention half-life of 2.7 ± 0.2 h
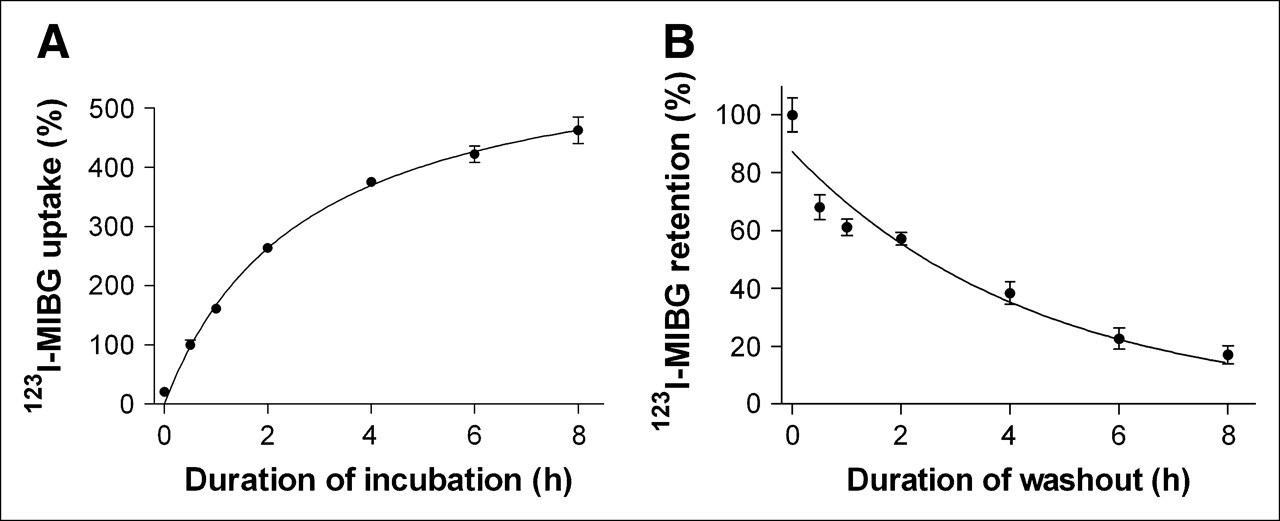
Time course of 123I-MIBG uptake and washout in CPAE cells. (A) 123I-MIBG uptake levels according to duration of incubation, expressed as cellular radioactivity relative to that after 30 min of incubation. (B) 123I-MIBG retention levels according to duration of time in 123I-MIBG-free fresh medium after 30 min of 123I-MIBG uptake, expressed as cellular radioactivity relative to that immediately after medium change. All results represent mean ± SD of triplicate samples from 1 experiment representative of 2 separate experiments.
NE Transporter Protein Expression in SK-N-SH and CPAE Cells
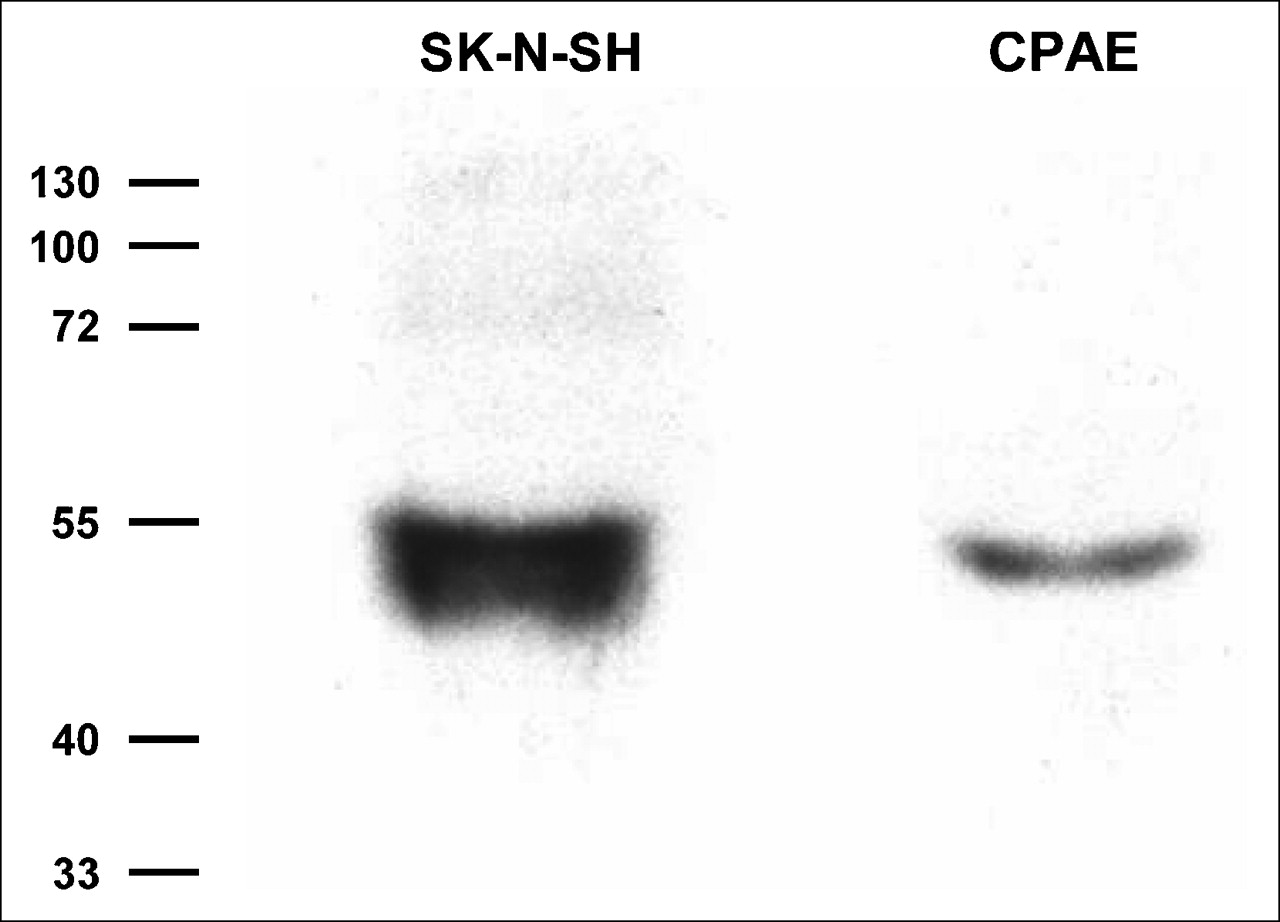
When the cellular expression of NE transporters was examined by Western blot analysis with monoclonal antibodies, SK-N-SH cells demonstrated a clear band at approximately 55 kd, consistent with previous reports for identical cells. CPAE cells also showed a clear band at the same molecular mass region, indicating the presence of NE transporters expressed in the cells (Fig. 2).
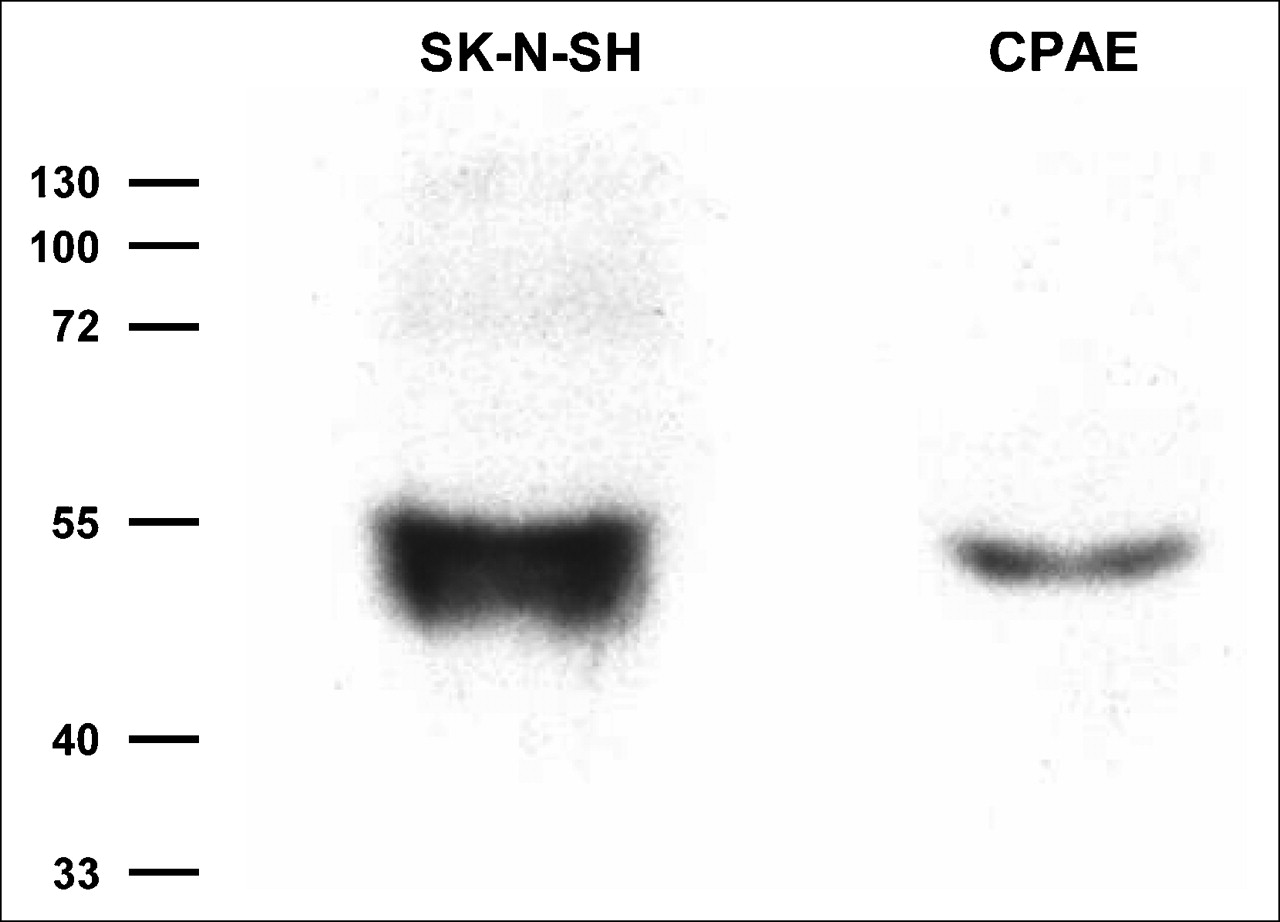
Representative Western blot for NE transporters. SK-N-SH cells are human neuroblastoma cells used as positive control cells. Molecular masses are given in kilodaltons.
Effects of Uptake 1 and Uptake 2 Inhibitors on Cellular 123I-MIBG Transport
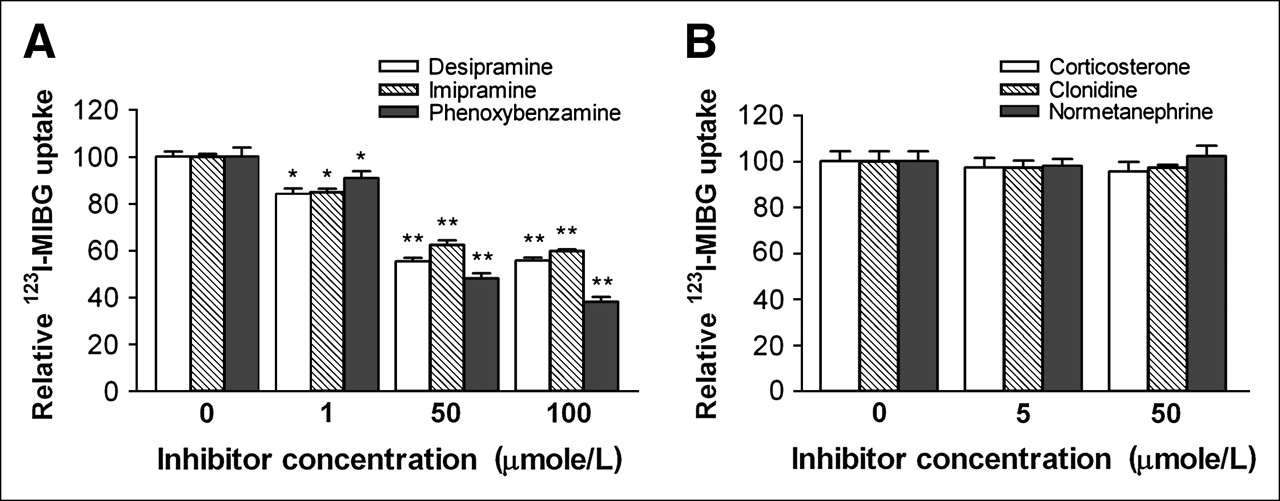
When CPAE cells were incubated with 123I-MIBG in the presence of the uptake 1 transport inhibitor desipramine or imipramine, there were significant and dose-dependent reductions in radiouptake. Desipramine, at concentrations of 1 and 50 μmol/L, reduced cellular 123I-MIBG uptake to 84.1% ± 4.0% and 55.3% ± 2.7% control levels, respectively, whereas the same concentrations of imipramine reduced the uptake to 84.8% ± 2.5% and 62.4% ± 3.5% control levels, respectively (Fig. 3A). Phenoxybenzamine, a nonselective adrenergic blocking agent with uptake 1 inhibitory action, also blocked 123I-MIBG uptake in a dose-dependent manner at 50 μmol/L, with uptake decreasing to 48.0% ± 2.2% control levels (Fig. 3A).

Effects of uptake 1 and uptake 2 inhibitors on CPAE cell 123I-MIBG uptake. Cells were pretreated with uptake 1 inhibitors desipramine, imipramine, and phenoxybenzamine at 0, 1, 50, or 100 μmol/L (A) and with uptake 2 inhibitors corticosterone, clonidine, and normetanephrine at 0, 5, or 50 μmol/L (B) for 30 min and then incubated with 123I-MIBG at 37°C for 30 min. Results are expressed as mean ± SD (triplicate samples) of percent cellular radioactivity relative to that of control cells and are from 1 experiment representative of 3 separate experiments. *P < 0.01. **P < 0.005.
In contrast to the uptake 1 inhibitors, none of the inhibitors of the uptake 2 mechanism tested had an effect on 123I-MIBG transport. As such, there was no difference in 123I-MIBG uptake levels between control cells and cells treated with corticosterone, clonidine, or normetanephrine at 5 or 50 μmol/L (Fig. 3B).
Effects of Hypoxia, PMA, and l-NAME on Cellular 123I-MIBG Uptake
When CPAE cells were exposed to hypoxic conditions, there was a time-dependent reduction in 123I-MIBG uptake, which decreased to 77.5% ± 0.2%, 57.8% ± 3.5%, and 50.0% ± 3.4% control (standard oxygen conditions) levels after 0.5, 1, and 4 h of hypoxia, respectively (Fig. 4A). When we evaluated the effect of PMA, concentrations of 5 and 10 ng/mL significantly reduced cellular 123I-MIBG uptake to 81.8% ± 5.8% and 76.7% ± 9.0% control levels, respectively (Fig. 4B). In addition, l-NAME at 2 nmol/L also was found to cause a mild but significant reduction in 123I-MIBG uptake to 86.5% ± 5.6% control levels (Fig. 4B).
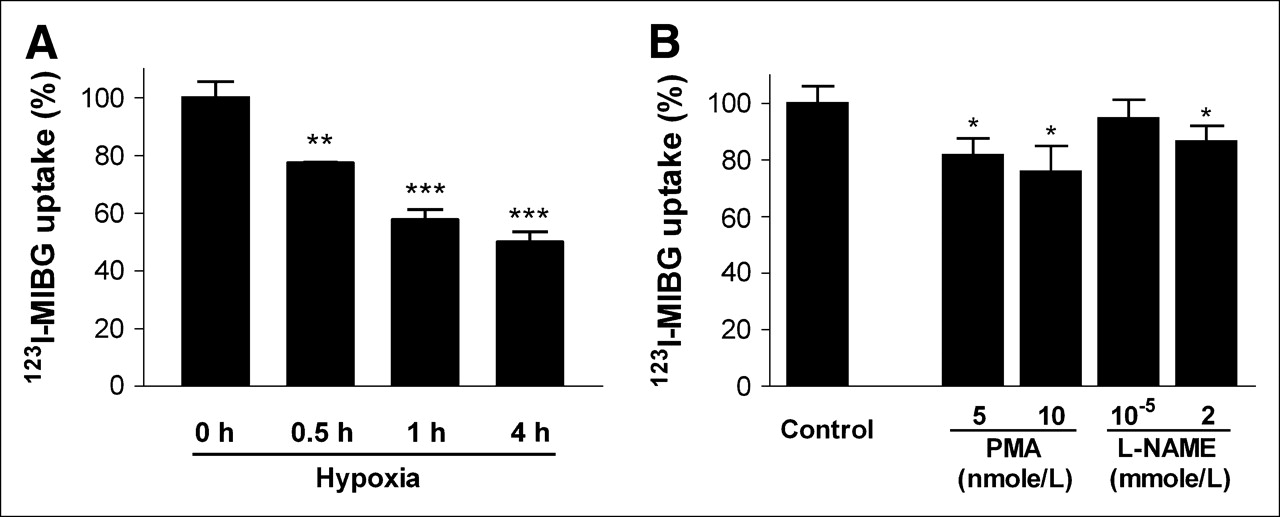
Effects of hypoxia, PMA, and l-NAME on CPAE cell 123I-MIBG uptake. (A) Cells were exposed to 1% oxygen for 0, 0.5, 1, or 4 h and then measured for uptake of 123I-MIBG after 30 min. (B) Cells were preincubated with PMA at 5 and 10 nmol/L or l-NAME at 10 μmol/L and 2 mmol/L for 16 h and then measured for uptake of 123I-MIBG after 30 min. Results are expressed as mean ± SD (triplicate samples) of percent cellular radioactivity relative to that of control cells and are from 1 experiment representative of 2 separate experiments. *P < 0.05. **P < 0.005. ***P < 0.001.
DISCUSSION
The lungs take up NE as part of an important metabolic function of regulating circulating levels of biogenic amines. The site for pulmonary removal of NE has been localized by fluorescence histochemistry (12) and autoradiography (13) to endothelial tissues that line the pulmonary microvasculature. However, whereas the characteristics of 123I-MIBG transport in neuroendocrine tumor cells have been elucidated extensively, studies on pulmonary 123I-MIBG processing have been limited to in vivo lung imaging in patients and uptake measurements in explanted lung tissue. In the present study, we investigated the characteristics of 123I-MIBG uptake in cultured pulmonary endothelial cells, and the results of our experiments confirmed that these cells express NE transporter proteins and actively take up 123I-MIBG through the specific uptake 1 mechanism. These findings reiterate, in several ways, what has been found in studies of isolated lung tissue. However, the distinct aspect of the present study is that 123I-MIBG processing as an indicator of pulmonary endothelial cell function was explored for the first time at the cellular level.
Previous studies on the kinetics of 123I-MIBG transport showed that human neuroblastoma cell lines, such as SK-N-SH cells, vigorously take up 123I-MIBG through both a nonspecific diffusion mechanism and a specific active uptake system (15–18). In the present study, CPAE cells were found to take up 123I-MIBG avidly in a time-dependent manner, and the uptake reached a relative plateau after 4 h of incubation. Washout studies demonstrated a gradual release of cellular 123I-MIBG after the initial uptake, with an efflux rate of 43% over 2 h. This rate of 123I-MIBG release is greater than the 24-h efflux rates of 20%–30% reported for SK-N-SH cells (15) but is comparable to the efflux rates of 30%–40% over 2–4 h that have been observed for NE transporter gene–transduced nonneuronal cells (19,20). Thus, the 123I-MIBG storage and release mechanism for CPAE cells appears to be more similar to that of other nonneuronal cells that lack chromaffin storage granules than to that of neuroblastoma cells (21).
As for NE, the specific transport of 123I-MIBG in SK-N-SH neuroblastoma cells occurs through a transport system characterized by high-affinity energy dependency and imipramine sensitivity (15–18). In our experiments, whereas none of the uptake 2 inhibitors tested had an effect, the 123I-MIBG uptake of CPAE cells was effectively blocked by the specific uptake 1 inhibitors desipramine and imipramine. Uptake also was reduced significantly by the nonselective adrenergic blocking agent phenoxybenzamine, which has known suppressive effects on uptake 1–mediated NE reuptake into nonneuronal tissues (22–24). These results indicate that a substantial portion of 123I-MIBG uptake in CPAE cells is mediated through the specific uptake 1 mechanism.
The uptake 1 mechanism selectively takes up NE through specific transporters (25), and 123I-MIBG uptake levels have been shown to correlate with NE transporter expression levels in both cultured cells (26,27) and neuroblastoma tumor models (28). Although the expression of NE transporters is known to be restricted almost exclusively to cells of the sympathetic nervous system, messenger RNA for NE transporters previously was detected by Northern blot analysis of pulmonary endothelial cells obtained from near-term fetal sheep (29). In the present study, we confirmed that NE transporter proteins are also present in CPAE cells. Western blots of proteins from CPAE cells revealed a distinct band that immunoreacted with an antibody against NE transporters; the location of this band was identical to the location of the band obtained from SK-N-SH cells and consistent with the known molecular mass of NE transporter proteins (30,31). Taken together, the results of the present study demonstrate that pulmonary endothelial cells actively take up 123I-MIBG, predominantly through an NE transporter–mediated uptake 1 mechanism.
The features of pulmonary processing of 123I-MIBG are of clinical interest because the metabolic function of biogenic amine extraction is recognized as a sensitive biochemical marker of lung pathology (14). Since its introduction as a radiotracer for the monitoring of amine processing by the lungs, 123I-MIBG scintigraphy has gained acceptance as a noninvasive method for assessing a variety of diseases that are accompanied by pulmonary endothelial cell damage (3–8). Therefore, we further investigated the effect on 123I-MIBG transport of exposure of CPAE cells to stimuli that have been associated with pulmonary endothelial cell dysfunction.
Low oxygen tension can affect endothelial cellular physiology in several ways (32). Animal experiments showed that newborns raised in a hypoxic environment have significantly reduced pulmonary extraction of radiolabeled NE (33), and healthy human subjects exposed to high-altitude hypoxia showed decreased lung 123I-MIBG activity in the early recovery phase; these results appear to reflect impaired pulmonary endothelial cell metabolic function (7). Similar findings also have been observed for the myocardium: rats submitted to hypoxia showed significantly reduced cardiac 123I-MIBG activity predominantly attributable to decreased NE uptake 1 transporter function (34). Another study demonstrated a hypoxia-induced reduction in the adrenergic neurotransmitter reserve in both the myocardium and the lungs by showing significantly reduced cardiac and pulmonary 123I-MIBG uptake in humans exposed to high-altitude hypoxia (35). In the present study, we found that short-term exposure to hypoxia was sufficient to induce a time-dependent reduction in CPAE cell 123I-MIBG uptake. Because these cells were deprived of oxygen for relatively short durations of time, this finding likely was not caused by energy depletion but rather reflected impaired NE transport function induced by hypoxia.
In addition to exposure to hypoxia, we also evaluated the effects of protein kinase C (PKC) activation by PMA and the inhibition of nitric oxide synthesis by l-NAME. The involvement of PKC activity in the regulation of endothelial cell monolayer integrity is well documented (36), and PKC-linked receptors have been shown to modulate the NE transport of neuroblastoma cells (37). In our experiments, PMA led to a dose-dependent reduction in 123I-MIBG uptake, suggesting the possibility that the NE transport capacity of pulmonary endothelial cells also may be modulated by PKC. Nitric oxide is a biologic vasoactive agent important to normal endothelial cell function. Nitric oxide synthase activity is expressed constitutively in endothelial cells, and insufficient nitric oxide production by dysfunctional endothelial cells has been implicated in the development of various pulmonary vascular diseases (32). In the present study, nitric oxide synthase inhibition by l-NAME caused a mild but significant reduction in 123I-MIBG uptake in CPAE cells.
Taken together, the results of our stimulation studies suggest that impairment of pulmonary endothelial cell transport function by hypoxia, nitric oxide deficiency, and PKC activation may contribute partly to the decreased lung 123I-MIBG uptake that is observed in various lung diseases. However, because the conditions of our cell-based experiments do not accurately reflect the conditions of lung tissues in living subjects, caution should be applied to generalizing our observations to clinical settings in patients with lung diseases.
CONCLUSION
In conclusion, our study confirms that pulmonary endothelial cells express NE transporters and actively take up 123I-MIBG; a significant portion of this activity occurs through the uptake 1 system. Furthermore, the cells show significantly reduced 123I-MIBG transport when exposed to potentially injurious stimuli, such as hypoxia, decreased availability of nitric oxide, and PKC activation. These experiments support the validity of lung 123I-MIBG imaging for monitoring pulmonary endothelial cell integrity and demonstrate that pulmonary endothelial cell models may be useful for investigating factors that affect pulmonary 123I-MIBG kinetics.
Acknowledgments
The authors thank Byung-Joo Yoo for technical assistance. This work was supported by Samsung grant SBRI C-A6-419-1. This work was presented in part at the 52nd Annual Meeting of the Society of Nuclear Medicine, Toronto, Ontario, Canada, June 18–22, 2005.
References
- Received for publication October 20, 2005.
- Accepted for publication December 1, 2005.





{kind=link}
{kind=link}
{kind=link}
{kind=link}