


Abstract
Our objective was to determine whether multiple clinically useful radiotracers accumulate in brown adipose tissue (BAT) and to assess their uptake in rats kept at room temperature or exposed to a cold environment. Methods: The following radiotracers were injected intravenously into groups of 6 female Wistar rats: 201Tl-chloride (TlCl), 123I-metaiodobenzylguanidine (MIBG), 99mTc-sestamibi (MIBI), 18F- or 3H-FDG, 3H-l-methionine, and 3H-thymidine. BAT-stimulated animals were maintained at 4°C for 4 h before tracer injection, whereas control animals were kept at approximately 22.5°C. The animals were sacrificed at 20–60 min after tracer injection, and BAT, major organs, and blood were extracted, weighed, and measured for radioactivity. The localization of uncoupling protein-1, glucose transporter-1, and norepinephrine transporter was evaluated with immunohistochemical staining in both groups. Results: We determined the percentage injected dose (%ID) per gram of each radiotracer in interscapular BAT, normalized to blood %ID/g. In control animals, this uptake ratio (±SD) was 8.44 ± 3.39 for 201TlCl, 9.77 ± 6.06 for 123I-MIBG, 37.30 ± 14.42 for 99mTc-MIBI, 5.47 ± 4.44 for 18F- or 3H-FDG, 1.93 ± 0.87 for 3H-l-methionine, and 1.22 ± 0.74 for 3H-thymidine. Compared with uptake at room temperature, uptake after exposure to cold increased 26.4-fold (P < 0.01) for 18F- or 3H-FDG and increased significantly (P < 0.05) for 201Tl (2.04-fold), 123I-MIBG (3.25-fold), and 3H-l-methionine (3.11-fold). Immunohistochemical staining revealed increased glucose transporter-1 and norepinephrine transporter expression in BAT cell membranes and blood vessels after exposure to cold, whereas uncoupling protein-1 was expressed in the cytoplasm under both control and cold-stimulated conditions. Conclusion: BAT uptake of 18F- or 3H-FDG, 123I-MIBG, and 3H-l-methionine was significantly increased over the control state by exposure to cold. Increased uptake of 201TlCl relative to blood in cold-stimulated BAT suggests that blood flow in BAT is increased by exposure to cold. The greater increased uptake with 18F- or 3H-FDG, 123I-MIBG, and 3H-l-methionine, and the immunohistostaining findings, suggest that other factors in addition to blood flow (e.g., increased metabolism, increased transport, or metabolic trapping of the tracers) are involved in cold-stimulated BAT activation. Knowledge that high uptake in BAT may possibly be observed on clinical scans using several radiotracers, especially after patients are exposed to the cold, may lead to more accurate interpretation of clinical studies.
Brown adipose tissue (BAT) differs from white adipose tissue morphologically. BAT is so named because of the visible brownish color resulting from its rich vascularization and increased mitochondrial content. BAT is generally located in deep cervical regions, including the supraclavicular area, the interscapular and paravertebral regions, and areas near large blood vessels. It is characterized by a unique metabolic pathway that results in the generation of heat. This heat generation by BAT is initiated by activation of the sympathetic nervous system, especially via activation of the β3-adrenergic receptor. Signals of decreasing body temperature, recent feeding, and high reserves of body energy have been reported to be the initial triggers that stimulate BAT (1).
BAT has recently received considerable attention in clinical nuclear medicine (2). High uptake of 18F-FDG in nonmalignant supraclavicular, thoracic costovertebral, and upper abdominal regions has been explained as uptake in BAT (3,4). This high uptake may sometimes be indistinguishable from uptake in tumors or lymph nodes. Increased uptake in the supraclavicular region has been reported to occur in 2.5%–4.0% of patients undergoing 18F-FDG PET/CT studies (5), is more common in women, and in some reports is more common in winter (3,5). There is also evidence that 18F-FDG uptake is reduced by controlling environmental temperature before a PET scan (6). Understanding the function of BAT and the influence of exposure to cold on BAT activity is potentially important for the interpretation of 18F-FDG PET/CT studies.
High tracer uptake in BAT has been reported for clinical radiotracers other than 18F-FDG. Okuyama et al. demonstrated that 123I-metaiodobenzylguanidine (MIBG) accumulated in the BAT of experimental animals and that 123I-MIBG uptake in these regions in children occurred more often in cold weather than in warm weather (7,8). These results are consistent with the hypothesis that the increased incidence of high radiotracer uptake in BAT is related to activation of BAT. Fukuchi et al. reported increased uptake of 99mTc-tetrofosmin in BAT in pediatric patients (9). Higuchi et al. described a patient who displayed high 99mTc-sestamibi (MIBI) uptake in BAT but no uptake of 201Tl-chloride (TlCl) in a dual-tracer SPECT study (10). These studies indicate that 99mTc-labeled agents for cardiac perfusion may also reflect BAT activity, especially its rich mitochondrial function. Our current study originated from the question of what happens to BAT uptake when common SPECT and PET radiotracers are used in animals at room temperature (approximately 22°C–23°C) or under cold conditions.
BAT is known to be stimulated by physiologic or pharmacologic intervention (11,12), and activation of BAT may result in an increase in radiotracer uptake. This uptake may represent the metabolic activity of BAT. However, it is still unknown which radiotracers accumulate in BAT and how this uptake will be changed by BAT activation.
We previously reported that uptake of 18F-FDG in BAT can be increased by a short period of exposure to cold (4°C for 4 h), suggesting that the procedure is suitable for a simple BAT-activation model (11). The purpose of this study was to systematically evaluate the BAT uptake of various clinically useful radiotracers using a cold-activation method in rodents. We also evaluated the degree of expression and the localization of uncoupling protein-1 (UCP-1), glucose transporter-1 (Glut-1), and norepinephrine transporter (NET) by immunohistostaining. An awareness of the distribution of commonly used clinical radiotracers in BAT, normally and after exposure to cold, should help avoid errors in the interpretation of various nuclear medicine studies.
MATERIALS AND METHODS
Animals
Eight- to 12-wk-old female Wistar rats (mean body weight, 225 g) were used in this study and were kept in the animal housing facilities at least 1 wk before the start of experimentation. Food and water were given ad libitum. The study was performed as part of a project approved by the animal research committee at the Johns Hopkins Medical Institutions.
BAT Activation
To stimulate BAT activity, we maintained the experimental animals at 4°C for 4 h before tracer injection, whereas control animals were kept at room temperature (approximately 22°C–23°C). We chose this condition (4°C for 4 h) according to previous reports to ensure that the BAT of all animals was stimulated adequately and reproducibly (1,11,13). To avoid the effect of anesthesia on BAT activity, no anesthetic agent was used in any procedure throughout the study except for euthanasia.
Biodistribution Study
Tracers were injected intravenously into groups of 6 rats each. The uptake time was 20 min for 201TlCl, 20 min for 123I-MIBG, 60 min for 99mTc-MIBI, 60 min for 18F- or 3H-FDG, 60 min for 3H-l-methionine, and 20 min for 3H-thymidine. A dose of 7,400 kBq (200 μCi) and 185 kBq (5 μCi) per animal was injected for γ-emitters and tritium-labeled tracers, respectively. In some experimental groups, γ-emitting and tritium-labeled isotopes were simultaneously given by dual injection (e.g., 18F-FDG plus 3H-l-methionine, or 99mTc-MIBI plus 3H-FDG). Both γ-emitters and tritium-labeled tracers were used in the FDG group. Animals in the cold-exposure group were kept in the cold after tracer injection. After the uptake time appropriate for each tracer, the animals were sacrificed; BAT, major organs, and blood were extracted and weighed; and tissue radioactivity was measured with a γ-counter. For the dual-tracer injection groups, tissue samples were then dissolved in solution (Solvable; Perkins Elmer) and the radioactivity of tritium was measured using a liquid scintillation counter. To avoid contamination from the γ-radiotracer, the tritium samples were counted after at least 20 half-lives of the coinjected γ-emitter.
The percentage injected dose (%ID) per gram of tissue normalized by kilogram of body weight (%ID/g × kg [%ID/body weight]), as well as tissue %ID/g standardized by %ID/g of blood (%ID/blood) and by %ID/g of muscle (%ID/muscle), was calculated and compared among tissues. Except for cold activation, all procedures including radiotracer injection were performed at room temperature (approximately 22°C–23°C) in a climate-controlled room.
Immunohistostaining
To evaluate cold-induced histologic changes of BAT, histologic (hematoxylin–eosin staining) and immunohistochemical evaluations were performed on samples obtained from animals under the control and the cold-stimulated conditions. The rabbit antimitochondrial UCP-1 antibody, anti–Glut-1 antibody, and anti-NET antibody were used for immunohistochemical evaluation. Antibody staining was performed using the avidin-biotin-peroxidase (avidin–biotin complex) method on dewaxed 5-μm-thick sections from a formalin-fixed, paraffin-embedded sample. The details of this procedure have been described elsewhere (14). The polyclonal rabbit anti-UCP-1 antibody (Sigma), the polyclonal rabbit anti–Glut-1 antibody (Chemicon), and the polyclonal rabbit anti-NET antibody (Chemicon) were used as primary antibodies. Each of these antibodies was used in a 1:300 dilution in separate studies. Samples were reviewed by light microscopy to determine the presence and location of these proteins.
Statistical Analysis
The data for the cold-stimulated group (%ID/body weight, %ID/blood, and %ID/muscle) were compared with the data for the control group using the Mann–Whitney U test. A P value of less than 0.05 was considered statistically significant.
RESULTS
Interscapular BAT was easily located in each rat at the time of dissection. Interscapular BAT in the cold-exposure group was reddish brown without exception on direct visual inspection, whereas interscapular BAT in the control group was whitish brown (Fig. 1A). On staining of interscapular BAT with hematoxylin–eosin, numerous lipid vacuoles were seen in the BAT cells from the control animals (Fig. 1B), but most of these lipid vacuoles were not present in the animals exposed to cold (Fig. 1C).
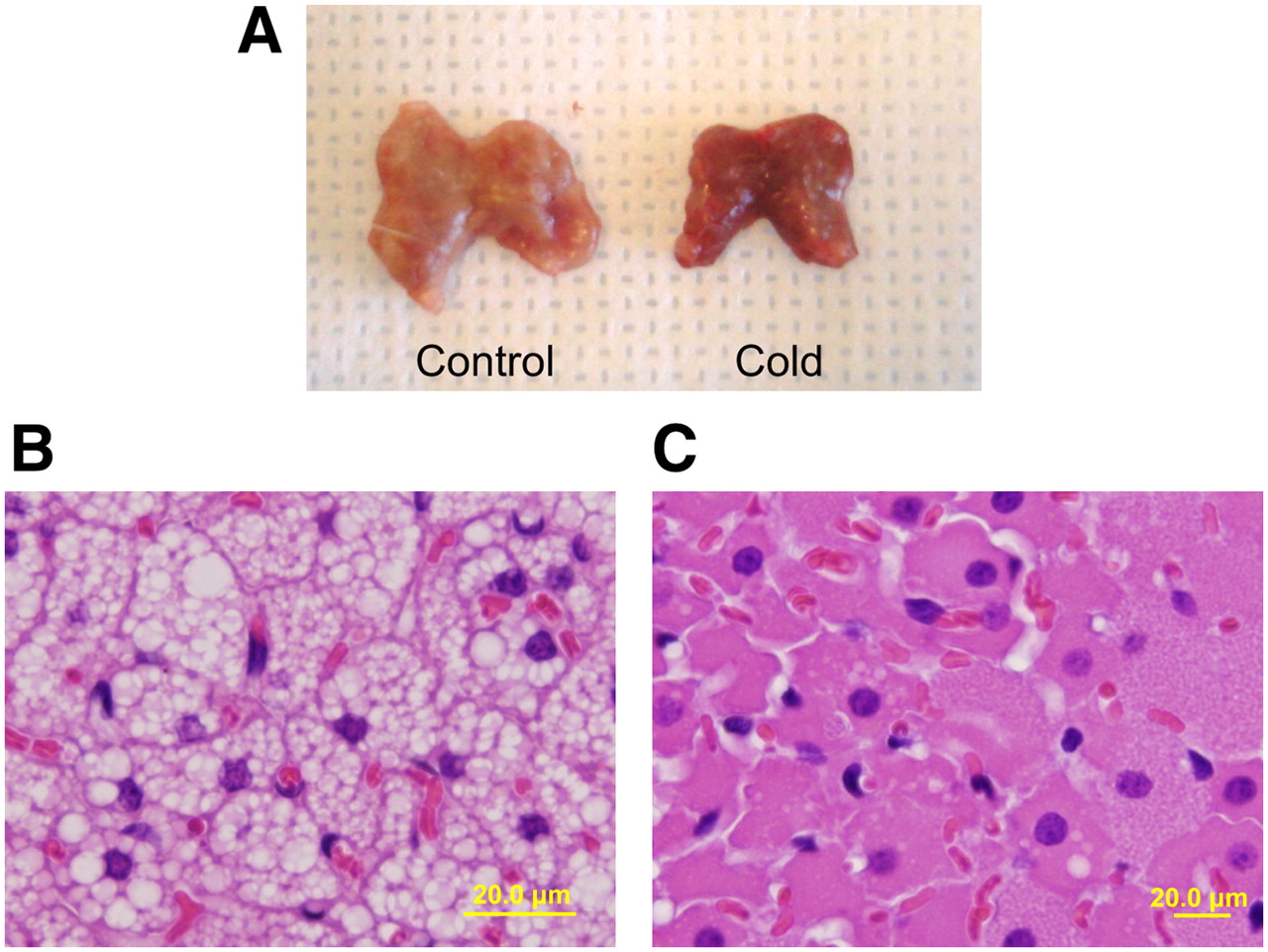
(A) Resected interscapular BAT after 4 h of exposure to cold (right) becomes darker and redder than control BAT kept at room temperature (left). (B and C) On light microscopy, major structural changes are seen in BAT cells stained with hematoxylin–eosin. Many large lipid vacuoles are seen in control BAT cells (B), but vacuoles are almost nonexistent in BAT cells exposed to cold (C). Effect of exposure to cold is grossly visible.
%ID/blood is summarized in Figure 2A. Under control conditions, 99mTc-MIBI had the highest uptake among these tracers. %ID/blood was lower for 18F- or 3H-FDG than for 201TlCl, 123I-MIBG, and 99mTc-MIBI under control conditions. After stimulation by cold, %ID/blood was increased, compared with %ID/blood under nonstimulated conditions: 2.04-fold for 201TlCl, 3.25-fold for 123I-MIBG, 1.39-fold for 99mTc-MIBI, 26.4-fold for 18F- or 3H-FDG, 3.11-fold for 3H-l-methionine, and 2.17-fold for 3H-thymidine (Fig. 2A). A statistically significant (P < 0.05) increase was seen for 201TlCl, 123I-MIBG, 18F- or 3H-FDG, and 3H-l-methionine. BAT uptake of 99mTc-MIBI tended to increase with exposure to cold (P = 0.07). Uptake of 3H-thymidine did not change significantly (P = 0.17). Changes in %ID/blood for other major organs are summarized in Table 1. Changes occurred in brain, stomach, and spleen (decreased uptake with 18F- or 3H-FDG); white adipose tissue (increased uptake with 3H-l-methionine); and kidney (increased uptake with 99mTc-MIBI).

%ID/blood (A), %ID/muscle (B), and %ID/body weight (%ID/g × kg [b.w.]) (C) for various tracers in interscapular BAT of control rats (white) and cold-stimulated rats (black). Under control conditions, highest uptake was seen in 123I-MIBG or 99mTc-MIBI. After cold activation, average uptake increased for all tracers. A, B, and C show statistically significant increases (P < 0.05) in tracer uptake in rats injected with 123I-MIBG, 18F- or 3H-FDG, and 3H-l-methionine. Increase in uptake of 201TlCl, compared with control, was also significant when normalized to blood.
%ID/Blood Values for Major Organs
%ID/muscle and %ID/body weight are summarized in Figures 2B and 2C, respectively.
Under control conditions, 123I-MIBG had the highest value for %ID/body weight among the tracers. Uptake of 18F- or 3H-FDG was relatively low among the tracers under control conditions. After stimulation by cold, statistically significant (P < 0.05) increases were seen for 123I-MIBG, 18F- or 3H-FDG, and 3H-l-methionine. No significant differences in uptake were seen between 18F-FDG and 3H-FDG (data not shown). Changes in %ID/muscle and %ID/body weight for other major organs are summarized in Tables 2 and 3, respectively. Changes occurred in the stomach and spleen (decreased uptake with 18F- or 3H-FDG) and in the liver (increased uptake with 3H-l-methionine).
%ID/Muscle Values for Major Organs
%ID/Body Weight Values for Major Organs
Immunohistolocalization of UCP-1, Glut-1, and NET in BAT under nonstimulated and cold-stimulated conditions is shown in Figure 3. Dense cytoplasmic staining with UCP-1 was seen in BAT under both the control and the cold-stimulated conditions. Partly because of the markedly reduced lipid content in cold-stimulated BAT, the total cytoplasmic UCP-1 content appeared somewhat higher. Staining with GLUT-1 was faint in control BAT cells, whereas dense staining was seen evenly in the plasma membrane of cold-stimulated BAT cells and blood vessels. NET expression was also increased by exposure to cold. However, the distribution of NET was different from that of GLUT-1: Dense staining was seen along the capillary blood vessels, and mild staining was seen in the plasma membrane of BAT cells.
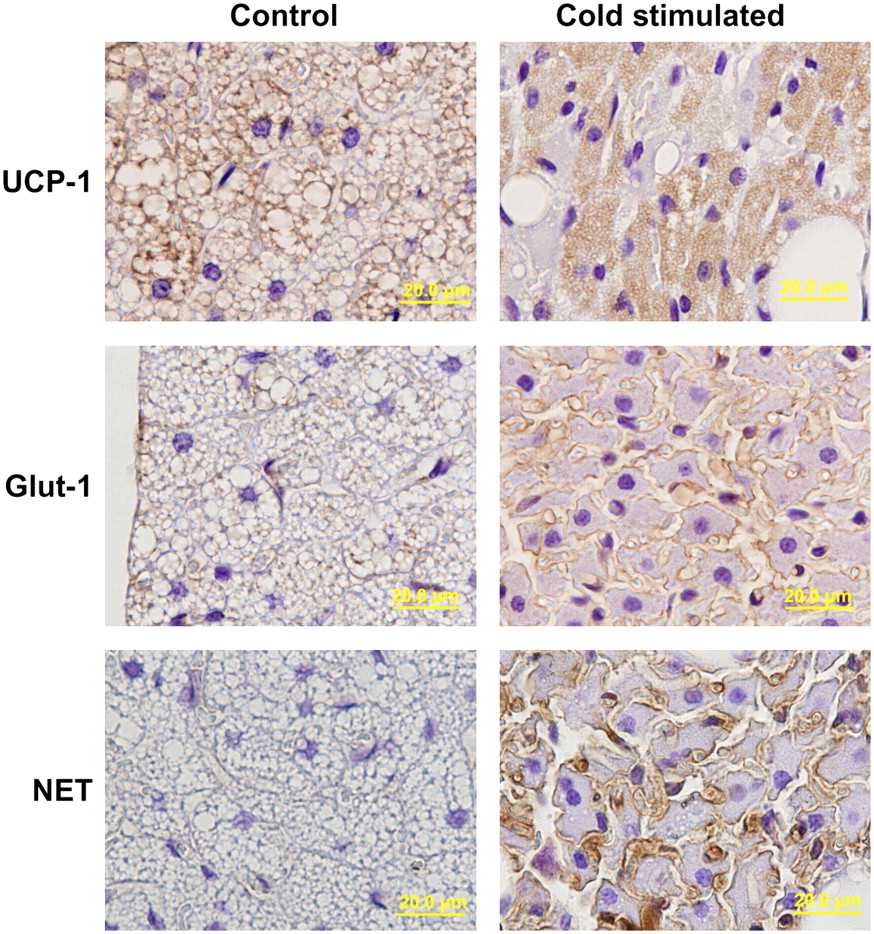
Immunohistolocalization of UCP-1, Glut-1, and NET in BAT under control and cold-stimulated conditions. Dense cytoplasmic staining of UCP-1 is seen in BAT under both conditions. Staining of GLUT-1 is faint under control conditions, whereas definite staining is evenly distributed in plasma membrane of BAT under stimulated conditions. NET is also stimulated by cold; however, its distribution is different from that of GLUT-1. Dense staining is seen along with capillary blood vessel, and mild staining was seen in plasma membrane of BAT cells.
DISCUSSION
Information about body temperature, feeding status, and body energy reserve is coordinated in the ventromedial hypothalamic nucleus, which signals the release of norepinephrine via the sympathetic nervous system and initiates triglyceride breakdown in BAT primarily via β3-adrenergic receptors. Stimulation of norepinephrine induces increased lipoprotein lipase, leading to the release of triglycerides and fatty acids, which are both the acute substrate for thermogenesis and the regulators of UCP-1 activity (1). Triglycerides stored in BAT itself are a major resource for thermogenesis, as well as for glucose to cope with stimulation by acute cold. Our results show that the number of lipid vacuoles in BAT was substantially decreased after stimulation by cold (Figs. 1B and 1C). Thomson et al. reported that a reduction in fat droplet size in BAT was observed during the first 3 d of stimulation by cold and was followed by a gradual increase in size over a 6-wk period (15). In our study, BAT changed to a reddish and dark color after exposure to cold (Fig. 1A). This change may be due to both an increase in blood flow and a decrease in the lipid content of BAT.
After stimulation by cold, statistically significant (P < 0.05) increases in radiotracer uptake were seen for 123I-MIBG, 18F- or 3H-FDG, and 3H-l-methionine. The increase in uptake of 201TlCl, compared with control, was also significant when normalized to blood. In humans, the major structures surrounding BAT are muscle and blood vessels. Thus, activity in these nearby tissues is highly relevant to clinical imaging results. Detection of abnormal signals requires comparison with these normal structures.
We have provided data on tracer uptake normalized to both blood and muscle, as well as the absolute %ID/body weight data. Because normalization to the blood background value also reduces any variability that may be due to differences in injection technique, we have emphasized the tracer uptake normalized by blood in this study.
A cold-stimulated increase in uptake of 201TlCl may result from increased blood flow in BAT. Astrup et al. reported that BAT blood flow was increased by ephedrine stimulation and resulted in a high temperature in the interscapular subcutaneous region (16). This result of Astrup et al. suggests that blood flow in BAT may be increased by changes in metabolic activity induced not only by ephedrine but also by exposure to cold.
With 18F- or 3H-FDG, BAT uptake was quite low under control conditions and increased significantly (26.5-fold; P < 0.01) after stimulation by cold. This increase could be interpreted as a result of increased glucose uptake and consumption by BAT, as is also supported by the Glut-1 immunohistostaining result indicating that the facilitated glucose transporter was stimulated and contributed to increased uptake of 18F- or 3H-FDG. These findings are consistent with previous reports (17,18).
123I-MIBG is known to accumulate in the sympathetic nerve terminals, and BAT activation is substantially controlled by the sympathetic nervous system (1). Some reports suggest activation of the sympathetic nervous system is an essential step to activating BAT. Okuyama et al. reported high 123I-MIBG uptake in BAT (8). Our results showed that the increase in 123I-MIBG uptake was 3.25-fold higher after exposure to cold than under control conditions. This increase is somewhat higher than that of 201TlCl, implying that factors other than blood flow are likely to be involved. The results of immunohistostaining with an anti-NET antibody revealed that expression of the NET protein was induced in capillary vessels after cold activation and might facilitate uptake of norepinephrine into BAT. Wakayama et al. reported that NET is expressed in the endothelial cells of capillary vessels in the brain (19). NET also appears to be expressed in the endothelial cells of capillary vessels in BAT. We believe this phenomenon indicates that the peripheral sympathetic nervous system in BAT is stimulated by a signal from the hypothalamus under cold conditions. As a result, the activity of NET increases and results in increased uptake of 123I-MIBG. 123I-MIBG uptake and NET expression have previously been reported to correlate well (20). This process may be related to the enlargement of vascular vessels and increase in blood flow that occur in stimulated BAT.
99mTc-MIBI is a tracer that was originally designed to investigate myocardial perfusion. This tracer is taken up in normal mitochondria via passive or active transport into the cell (21–23), and images of this tracer represent the function of the mitochondria, their density, and tracer delivery, especially with longer uptake times. Fukuchi et al. reported that high 99mTc-MIBI uptake is seen in the cervical region of children, especially in winter (9), and suggested that 99mTc-MIBI uptake into BAT was stimulated by cold temperatures. No significantly increased uptake was observed in our cold-activation model for 99mTc-MIBI. This discrepancy may partly be due to the mechanism of BAT activation. Chronic exposure to cold, such as during a seasonal change in climate, leads to an increase in the mass of BAT. From our results, uptake of 99mTc-MIBI into BAT relative to blood was the highest among the 6 tracers we evaluated in this study, even in the nonstimulated state, and BAT was the organ with the third highest uptake in the body, less than uptake in the kidney and heart (Table 1). These findings indicate that BAT may likely be visualized on clinical 99mTc-MIBI scans even in the nonstimulated state, especially in children, who have relatively more BAT than do adults. If the total amount of BAT increases, the incidence of high uptake seen on a scintigram may also increase. Another explanation is that 99mTc-MIBI is known as an agent with high lipophilicity (24) and may wash out when cold activation produces an acute decrease in the fat content of BAT. As shown in Figure 1, the total lipid content in BAT was dramatically reduced after acute exposure to cold. Increased 99mTc-MIBI uptake in BAT may have been offset by rapid lipid clearance.
Significantly increased uptake of 3H-l-methionine was observed after stimulation by cold in our experiment. BAT has glycolytic capabilities, as well as enzymes involved in amino acid metabolism (25). Uptake and release of several amino acids in BAT was evaluated by Lopez-Soriano et al., who studied arteriovenous differences in vivo (13). Our results indicate that increased 3H-l-methionine metabolism in BAT is also measurable with radiolabeled tracers and may potentially be visualized as high uptake in 3H-l-methionine PET studies.
Although not statistically significant (P = 0.17), increased tracer uptake after stimulation by cold was also seen in the animal groups that received 3H-thymidine. The blood-normalized uptake ratio of activation/nonactivation was 2.17 for 3H-thymidine—similar to that of 201TlCl (2.04). A longer observation time may have increased the differences in 3H-thymidine uptake.
Some significant changes in radiotracer uptake relative to blood after exposure to cold were seen in other major organs. For example, changes occurred in the brain, stomach, and spleen (decreased uptake with 18F- or 3H-FDG), white adipose tissue (increased uptake with 3H-l-methionine), and kidney (increased uptake with 99mTc-MIBI). We cannot fully explain these findings. A possible explanation is decreased uptake of 18F- or 3H-FDG in the stomach because of a decrease in gastric peristalsis via cold-stimulated activation of the sympathetic nervous system. Lower brain 18F- or 3H-FDG uptake in cold-stimulated animals may partly be due to decreased sensory stimulation during exposure to cold. The P values for most organs were relatively high for all tracers, suggesting that BAT is the major organ influenced by exposure to cold (Table 1).
UCP-1 is expressed only in BAT and is a key protein that enables mitochondrial oxidation not by coupling with adenosine triphosphate synthesis but rather by promoting heat generation (1). Initially, we anticipated some differences in UCP-1 expression between the control group and the cold-stimulated group. However, UCP-1 expression was seen under both control and stimulated conditions, suggesting that UCP-1 is likely constitutively expressed in BAT. Thus, BAT expresses UCP-1 protein under both nonstimulated and stimulated conditions. Substantial morphologic changes make it difficult to assess small changes in absolute UCP-1 expression. In this study, we did not perform a quantitative analysis of this protein. It is therefore difficult to say whether a true change in the expression level of this protein occurs with stimulation by cold. We believe that this result assists our understanding of how UCP-1 is expressed under control conditions. Staining with UCP-1 is also important by providing clear evidence that what we were evaluating was truly BAT.
To our knowledge, no previous reports have described cold-stimulated changes in the uptake of various clinical radiotracers by BAT under identical conditions. Our results indicate that high tracer uptake would be expected to be observed in clinical SPECT studies using 201TlCl or 123I-MIBG and in PET studies using 11C-methionine or 18F-FDG. It is important to know the behavior of various clinical tracers in BAT—especially when stimulated by a low-temperature environment—for correct interpretation of nuclear medicine images.
CONCLUSION
BAT uptake (%ID/blood) of 18F- or 3H-FDG, 123I-MIBG, 201TlCl, and 3H-l-methionine was significantly higher after exposure to cold than in the control state. The greatest increase in tracer uptake was for 18F- and 3H-FDG. Increased uptake of 201TlCl in cold-stimulated BAT suggests that blood flow in BAT is increased by exposure to cold, but the greater increased uptake for 123I-MIBG, 3H-l-methionine, and 18F- or 3H-FDG (P < 0.05) and the immunohistostaining findings indicate that other factors in addition to blood flow (e.g., increased metabolism) are involved in cold-stimulated BAT activation. Knowledge of the distribution of commonly used clinical radiotracers in BAT, normally and after exposure to cold, will help avoid errors in the interpretation of various nuclear medicine studies.
Acknowledgments
We thank Judy Buchanan for her assistance in the preparation of the manuscript.
Footnotes
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication March 14, 2007.
- Accepted for publication June 27, 2007.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Perspectives on Brown Adipose Tissue Imaging: Insights from Preclinical and Clinical Observations from the Last and Current Century
- The significance of beige and brown fat in humans
- Preclinical Evaluation of 18F-LMI1195 for In Vivo Imaging of Pheochromocytoma in the MENX Tumor Model
- Quantification of Human and Rodent Brown Adipose Tissue Function Using 99mTc-Methoxyisobutylisonitrile SPECT/CT and 18F-FDG PET/CT
- CT Hounsfield Units of Brown Adipose Tissue Increase with Activation: Preclinical and Clinical Studies
- Preliminary PET/CT Study of 18F-FDG Uptake in Cervical and Supraclavicular Brown Adipose Tissue
- Visualization of Brown Adipose Tissue with 99mTc-Methoxyisobutylisonitrile on SPECT/CT