


Abstract
Major depressive disorder (MDD) is a debilitating condition that affects over 14 million Americans. Remission occurs only in a minority of individuals after first-line antidepressant treatment (∼35%); predictors of treatment outcome are therefore needed. Using PET imaging with a radiotracer specific for the serotonin transporter (5-HTT), 11C-McN5652, we found that patients with MDD who did not achieve remission after 12 mo of naturalistic treatment had lower pretreatment midbrain and amygdala binding than healthy volunteers. Here, using a superior 5-HTT tracer, 11C-DASB, we repeated this study with a prospective design with 8 wk of standardized treatment with escitalopram. As this same cohort also underwent 11C-WAY100635 scans (serotonin-1A receptor [5-HT1A]), we examined whether using both pretreatment 5-HTT and 5-HT1A binding could improve prediction of posttreatment remission status. Methods: Thirty-one healthy controls (Hamilton Depression Rating Scale-24 item [HDRS-24] = 1.7) and 26 medication-free patients with MDD (HDRS-24 = 24.8) underwent PET scanning using 11C-DASB. MDD subjects then received 8 wk of standardized pharmacotherapy with escitalopram. The relationship between pretreatment binding and posttreatment clinical status was examined. Arterial blood samples were collected to calculate the metabolite-corrected arterial input function. The outcome measure was VT/fP (VT is volume of distribution in region of interest, fP is free fraction in plasma). Remission was defined as a posttreatment depression score of less than 10 as well as 50% or more reduction in the score from baseline, resulting in 14 nonremitters (HDRS-24 = 17.6) and 12 remitters (HDRS-24 = 5.3). Results: A linear mixed-effects model comparing group differences in the a priori regions of interest (amygdala and midbrain) revealed a significant difference in amygdala binding between controls and remitters (P = 0.03, unadjusted), where remitters had an 11% lower amygdala binding than controls. Differences in amygdala binding between remitters and nonremitters approached significance (P = 0.06). No additional differences were found between any groups (all P > 0.05). Additionally, we found no relationship between pretreatment amygdala binding and posttreatment depression score, and were unable to predict posttreatment depression severity using both pretreatment 5-HTT (in the amygdala) and 5-HT1A binding (in the raphe). Conclusion: These results suggest 5-HTT amygdala binding should be examined further, in conjunction with other measures, as a potential biomarker for remission after standardized escitalopram treatment.
Major depressive disorder (MDD) is a debilitating condition that affects over 14 million Americans (1,2) and is now the second leading cause of disability worldwide (3,4). MDD is a heterogeneous disorder comprised of clusters of symptoms; treating this disorder is, therefore, complex. Optimizing medication treatment is challenging and involves weeks of trial and error, which is stressful, uncomfortable, and sometimes life-threatening for patients. Remission occurs in a minority of individuals after first-line antidepressant treatment (30%–45%) (5), whereas 10%–30% of patients will not respond to any treatment (5,6). What is needed is a better understanding of the biology underlying this disorder, allowing for patient-specific treatment selection and improved response rates.
The serotonergic system has been implicated in the pathophysiology of depression and is a major target of antidepressant treatment (7,8). 5-HTT (serotonin transporter), the target of selective serotonin reuptake inhibitors (SSRIs), has been examined in MDD extensively (9–13) in postmortem and in vivo investigations. A metaanalysis including 25 in vivo studies found reduced 5-HTT binding in both the midbrain and the amygdala as well as the striatum, but not in the thalamus or hippocampus, in MDD compared with controls (9). Evaluations of postmortem studies also revealed decreases in overall 5-HTT levels in the amygdala and striatum (though these were few and did not achieve statistical significance), but not in the frontal cortex, brain stem, or hippocampus (9). Furthermore, low levels of 5-HTT binding in the amygdala have been associated with depression and anxiety (14).
A major advance in the study of psychiatric illness is the use of neuroimaging techniques. Using PET imaging with a 5-HTT tracer, 11C-McN5652, our group has found that 5-HTT binding differs between healthy controls and MDD patients in a depressive episode (11). Additionally, we found that nonremission from MDD after 12 mo of naturalistic treatment was associated with lower pretreatment midbrain and amygdala binding as compared with controls. When using a superior 5-HTT tracer (15,16), 11C-DASB, we were unable to replicate the differences between controls and patients with MDD (10); however, we found that suicide attempters had lower midbrain binding than nonattempters and controls. We postulated that perhaps the differences we initially observed within the MDD group may have been due to a heterogeneous population comprised of multiple clinical characteristics.
Here, we improve on the previous (naturalistic) study design using standardized treatment with SSRI escitalopram to investigate whether pretreatment 5-HTT binding (11C-DASB) is predictive of subsequent nonremission in the midbrain and amygdala. These previously examined regions of interest (ROIs) have been identified in several studies, highlighting their importance in the pathophysiology of MDD and treatment response. Additionally, we have examined serotonin-1A receptor (5-HT1A) binding in this same cohort (17) and found pretreatment raphe 5-HT1A binding significantly differs between remitters and nonremitters (where remitters had higher 5-HT1A binding) yet was unable to predict posttreatment depression severity (17). Here we examine the potential of 5-HT1A and 5-HTT binding to predict posttreatment depression severity.
MATERIALS AND METHODS
Subjects/Treatment
This study was approved by the Institutional Review Boards of the New York State Psychiatric Institute and Columbia University Medical Center. Thirty-one healthy volunteers and 26 subjects who met the DSM-IV criteria for MDD were included in this study. Data from all controls and 24 of the 26 MDD subjects were reported in a previous study using a different PET tracer, 11C-WAY10035 (17). Recruitment, consent, and eligibility were as previously described (17).
Clinical Procedures
MDD subjects were not taking medication (n = 19) or were weaned from ineffective medication (n = 6) before enrollment into the study (one subject’s previous medication status was unknown). After baseline PET and MRI scans, treatment was initiated with escitalopram. At week 4, the escitalopram dose was increased for nonresponders (<50% decrease in Hamilton Depression Rating Scale [HDRS]), whereas responders remained at 10 mg/d. At week 6, any subject who was a nonremitter (>10 HDRS) and was still taking 10 mg daily had the escitalopram dose increased to 20 mg per day. HDRS-24 was used to assess depression severity at baseline and at 8 wk. Remission status was defined as a 50% or more decrease in HDRS-24 score as well as a final HDRS-24 score of less than 10. Using these criteria, we classified 12 subjects as remitters and 14 subjects as nonremitters.
Radiochemistry
11C-DASB was prepared as previously described (18).
PET Scanning Protocol
PET scanning was conducted as previously described (19). In brief, PET images were acquired on an ECAT EXACT HR+ camera (Siemens/CTI). After a 10-min transmission scan, 11C-DASB was injected as an intravenous bolus, and emission data were collected for 100 min.
Image Processing
Image analysis was performed as previously described (10,12). In brief, motion correction of PET data was performed where frame-by-frame rigid-body registration was performed to a reference frame. A mean of the motion-corrected frames was then coregistered to the MRI. ROIs were hand drawn on the MR images of 18 healthy subject by experienced technicians trained to reliably approximate these regions using brain atlases (20,21) and published reports (22,23). To generate probabilistic ROIs, each subject’s MR image was preprocessed, and each of the 18 template images was registered with the Automated Registration Toolbox to the subject’s brain. The probability of a regional label was defined as the percentage of the 18 templates that assign the same regional label to that voxel.
Quantitative Analysis
Arterial samples were collected continuously for the first 4 min, and manually thereafter (every 10 s until 2 min, every 20 s until 4 min, every 2 min until 8 min, every 4 min until 20 min, and every 10 min until the end of the scan). The metabolite-corrected arterial input function was calculated by multiplying interpolated plasma radioactivity counts by the percentage parent compound and was fit using a straight line before the peak and the sum of 3 exponentials after the peak (19). The fitted values were used as the metabolite-corrected arterial input function. The volume of distribution in a region of interest (VT) estimates were obtained using likelihood estimation in graphical analysis and, with the arterial analysis, were used to calculate our primary outcome measure: VT/free fraction in plasma (fP) (10,19,24,25). Using this measure, estimated that binding does not rely on a reference region. It has been shown that an optimal reference region does not exist for 11C-DASB (26). Standard errors were computed for each estimated VT value using a bootstrapping algorithm that accounts for errors in plasma, metabolite, and brain data (27).
Voxelwise Average Images
VT/fP binding maps were calculated on a voxel-by-voxel level using empiric Bayesian estimation in graphical analysis, as previously described (28,29). For display, images were equally thresholded and displayed at the plane of the amygdala.
Statistical Analysis
Group comparisons of VT/fP between remitters, nonremitters, and controls were performed using a linear mixed-effects model with subject as the random effect and diagnosis, subject group, and age as covariates. The midbrain and amygdala were a priori ROIs. Additionally, a linear mixed model was used to determine the potential of pretreatment 11C-DASB binding covaried by pretreatment HDRS-24 to predict posttreatment HDRS-24. As a secondary analysis, we examined 4 additional ROIs (thalamus, dorsal putamen, hippocampus, and anterior cingulate cortex).
In the same cohort, our group found significantly higher pretreatment 5-HT1A binding in the raphe of remitters as compared with nonremitters (17), though we were unable to use 5-HT1A binding to predict posttreatment HDRS. Our group has previously shown that there is no correlation between 5-HTT binding and 5-HT1A binding (25), and accordingly we did not find a correlation here. This suggests that 11C-WAY-100635 and 11C-DASB quantification may provide complementary information. We postulated that a combined model using both raphe 5-HT1A and amygdala 5-HTT binding may better predict remission than either measure alone. Linear regression was used to determine whether pretreatment raphe (5-HT1A) binding and amygdala (5-HTT) binding, covaried for pretreatment HDRS-24, predicted posttreatment HDRS-24.
To allow for a test in proportional differences, to stabilize variance across regions, and to alleviate skewness, we and other groups used log-transformed data in all analyses involving multiple regions (10,11,26). Data presented graphically are displayed as actual (nonlog-transformed) data. Reported P values are unadjusted for multiple comparisons. Statistical tests were performed in SAS 9.3 (SAS Institute Inc.) and R 2.1.0 (http://cran.r-project.org)
RESULTS
Sample Characteristics
Summary statistics for clinical variables are presented in Table 1. No differences in HDRS-24 pretreatment score were found between the remitters and nonremitters. Significant differences in fP (P = 0.02) and trending differences in reference region (P = 0.07) binding were found between controls and remitters.
Sample Characteristics
5-HTT Binding in Remission
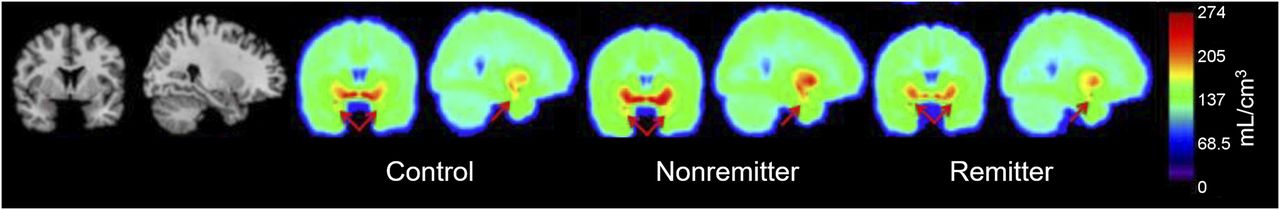
Remitters had lower 5-HTT pretreatment binding (VT/fP) than controls in the amygdala (P = 0.03, unadjusted; Figs. 1 and 2). Remitters had a trend of lower 5-HTT pretreatment binding in the amygdala than nonremitters, (P = 0.06, unadjusted). No differences in amygdala binding were found between nonremitters and controls (P = 0.97), or in the midbrain, thalamus, dorsal putamen, hippocampus, or anterior cingulate cortex between any groups (P > 0.05).

Comparison of binding potential by group in amygdala; weighted means and weighted errors are shown with black lines. *Significant difference between control and remitter where P < 0.05.

Averaged voxel images of control, remitter, and nonremitter 5-HTT binding (VT/fP) at plane of amygdala (red arrows).
Pretreatment Binding to Predict Posttreatment Depression Severity
Using a linear mixed model, we did not find a significant relationship between 5-HTT amygdala binding and posttreatment HDRS-24 score (P = 0.39). Linear regression revealed no correlation of raphe 5-HT1A and amygdala 5-HTT binding in predicting posttreatment HDRS-24 score (P = 0.24).
DISCUSSION
Pretreatment 5-HTT Amygdala Binding Differs in Remission
We observed lower pretreatment 5-HTT binding in the amygdalae of remitted patients than healthy controls, which remained a trend after correction for multiple comparisons. There was a trend toward lower 5-HTT binding in remitters than nonremitters. The lack of significance between remitters and nonremitters may have been due to the large variance and modest sample size in these groups. Additionally, we found no difference in midbrain, dorsal putamen, thalamic, hippocampal, and anterior cingulate cortex binding between any groups. These findings are at odds with previous results from Kuyaga et al., who reported higher pretreatment diencephalonic (midbrain subdivision) 5-HTT binding (SPECT) associated with improved response to acute fluoxetine or paroxetine treatment (4 and 6 wk, respectively) (30). It is possible that the discrepant findings are due to the use of 123I-β-CIT, a SPECT tracer known to bind to both 5-HTT and DAT (compared with 11C-DASB in the current study), as well as the use of an outcome measure that relies on reference region. These findings are in partial agreement with a metaanalysis that showed consistent reduction in 5-HTT levels in the amygdala in depression (31).
Pretreatment Binding to Predict Posttreatment Depression Severity
We did not find a relationship between pretreatment 5-HTT amygdala binding and posttreatment HDRS-24. Additionally, we were unable to predict depression severity using the 5-HT1A and 5-HTT combined data. These results are unsurprising as pretreatment 5-HTT amygdala binding between nonremitters and remitters was not significantly different. The addition of 5-HTT amygdala binding was, therefore, unable to strengthen the model. Interestingly, Lazenberger et al. found that pretreatment 5-HTT binding ratios (amygdala to raphe nucleus) were able to predict 3 wk posttreatment response using 11C-DASB (32). It is possible this discrepancy is due to the different measure that was used (ratio of amygdala to raphe nucleus) and the length of treatment before assessing outcome.
Choice of Binding Potential Measure
Here, we use the outcome measure VT/fP, which is conservative by not correcting for reference region binding and normalizes the data by fP. We chose this outcome measure for a few reasons. We report significantly lower free fraction in remitters than controls. This could be due to an elevation in tracer binding to peripheral plasma proteins, and could result in inaccurate 5-HTT density determination, as subjects with greater fp could have higher binding because more tracer is free to bind. An outcome measure that normalizes for free fraction differences at the individual level is needed.
We found that the difference in reference region binding between controls and remitters approaches significance, where remitters have lower reference region binding than controls. This further corroborates that the reference region (cerebellar gray matter) may not be devoid of specific binding (11). In this case, correcting VT by reference region binding could incorrectly estimate VT.
Differences in free fraction and reference region binding have been observed in other studies that use different tracers (33). These measures alone cannot provide us with any information about the specific neurotransmitter abnormalities in MDD and cannot be used as a surrogate for binding studies.
Interpretation of Findings
We previously found higher raphe 5-HT1A binding in remitters than controls and postulated that higher raphe 5-HT1A levels at baseline would cause low basal firing rate (17). Normal 5-HT transmission in the amygdala is thought to inhibit amygdala activity via activation of γ-aminobutryic acid (GABA) interneurons (34). A reduced basal firing rate may remove amygdala inhibition, resulting in hyperactivity. Hyperactivity in the amygdala has been shown to increase the salience of an aversive event and is observed in depression and anxiety (35–40).
In this study, we showed that this same cohort of subjects (remitters) has lower 5-HTT binding in the amygdala. We hypothesized that lower amygdala binding in the remitter group might be correlated with decreased 5-HTT levels. As partial validation of this, 5-HTT null mice do exhibit reduced firing rate in dorsal raphe neurons as compared with control mice (41), further supporting our previous hypothesis that low 5-HTT levels could contribute to a low basal firing rate.
Postmortem studies have found decreased 5-HTT levels in MDD (42–45). Interestingly, lower 5-HTT binding in the amygdala has been associated with high anxiety (46). Functional MRI studies have revealed amygdala hyperactivity in response to negative stimuli (36,47), similar to hyperactivity seen in subjects with MDD. Importantly, SSRI administration has been shown to alleviate this hyperactivity (36–38,48).
With acute SSRI administration raphe autoreceptors, sensitive to the increase in serotonin, are thought to decrease serotonergic firing. With repeated administration, the desensitization of autoreceptors may enhance serotonin release (49). An increase in transmission, coupled with previously low levels of 5-HTT, could enhance the effects of serotonin; with saturated transporters, prolonged effects of 5-HT are possible in the synapse. These data suggest that patients with lower amygdala 5-HTT levels coupled with elevated raphe 5-HT1A levels—patients with greater serotonergic abnormality—are more likely to benefit from serotonergic intervention (escitalopram). Interestingly, 1 study reported that remitted depressed individuals did not have lower 5-HTT binding in the amygdala and several other regions than controls, suggesting that these may not be trait properties, and instead state markers of a subset of the MDD population (50).
We found that MDD nonremitters, after 1 y of naturalistic treatment, had lower pretreatment amygdala binding than controls (12). These discrepant findings of lower amygdala binding in nonremission (previous study (12)) and remission (current study) could be due to study design (naturalistic vs. prospective), length after which treatment outcome was assessed (1 y vs. 8 wk), radiotracer (11C-McN5652 vs. 11C-DASB), and possible differences in the sampling of depressed patients.
This group of MDD subjects who remit after 8 wk of escitalopram treatment have higher raphe 5-HT1A levels and lower amygdala 5-HTT levels. It is unclear which is primary, though Gobbi et al. showed a decreased firing rate in dorsal raphe nuclei in 5-HTT null mice; thus, it is possible that 5-HTT may play a role in the regulation of 5-HT neurotransmission (41). Either way, these findings suggest that subjects with greater serotonergic abnormality benefit most from serotonergic intervention. As nonremitters do not exhibit serotonergic abnormalities, it is possible they require nonserotonergic intervention. We postulate that amygdala hyperactivity in nonremitters may be mediated by deficient GABAergic transmission and an inability to maintain sufficient control in the amygdala. There exists considerable evidence for low GABAergic transmission as a feature of MDD as well as anxiety and mood disorders (51,52). Alternatively, it is possible that hyperactivity in the amygdala is a result of elevated glutamatergic transmission (35), as there is increasing support for glutamatergic intervention in psychiatric disorders including stress and anxiety, as well as MDD (53,54). Further studies investigating GABAergic-mediated disinhibition and glutamatergic hyperactivity in MDD are needed.
CONCLUSION
These results, coupled with previous findings investigating 5-HT1A binding in the same cohort, suggest that subjects with greater serotonergic abnormality benefit most from serotonergic intervention. These findings move toward using PET to improve treatment selection. Follow-up PET studies would benefit from pre-/posttreatment scans, increased sample size, and comparison with nonserotonergic treatment and placebo (to determine whether this finding is specific to escitalopram). These measures will better allow us to validate the possibility of the amygdala and raphe as biomarkers of remission after 8 wk of standardized SSRI treatment.
DISCLOSURE
No potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Dr. Hao Chen from Stony Brook University and Drs. Jeff Miller and Todd Ogden from Columbia University for their comments and help in data acquisition and processing. We acknowledge the support from the Biostatistical Consulting Core at the School of Medicine, Stony Brook University.
Footnotes
Published online Sep. 21, 2017.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication January 8, 2017.
- Accepted for publication July 25, 2017.





{kind=link}
{kind=link}