


Abstract
αvβ6 integrin is overexpressed by several carcinomas and thus considered a target for diagnostic imaging and anticancer therapies. Recently, we presented the αvβ6 integrin-binding peptide SFITGv6 as a novel potential tracer for imaging and targeted therapy of αvβ6 integrin–positive carcinomas. Here, we analyzed the affinity and specificity of 5 native αvβ6 integrin–specific binders in comparison to SFITGv6. Methods: Sunflower trypsin inhibitor 1 (SFTI1)–based peptides containing arginine-glycine-aspartic acid (RGD) motif-spanning octamers of fibronectin (SFFN1), tenascin C (SFTNC), vitronectin (SFVTN), and latency-associated peptides (LAP) 1 (SFLAP1) and 3 (SFLAP3) were synthesized, and their binding potential to αvβ6 integrin–expressing head and neck squamous cell carcinoma (HNSCC) cell lines was evaluated. Subsequently, stability, affinity, and specificity were assessed in vitro using radio–high-pressure liquid chromatography, surface plasmon resonance assay, and binding experiments including competition, kinetics, internalization, and efflux. αvβ6 integrin binding specificity was further evaluated by peptide histochemistry. Finally, in vivo binding properties were assessed using small-animal PET imaging and biodistribution experiments in HNSCC-bearing mice, and 68Ga-DOTA-SFLAP3 was applied for diagnostic PET/CT of an HNSCC patient. Results: When the newly designed peptides were compared, significant binding (>20%) to several HNSCC cell lines (HNO97, HNO399, and HNO223) and a fast internalization of up to 60% and 70% were observed for SFLAP3 (GRGDLGRL) and SFITGv6 (FRGDLMQL). In contrast, the other peptides displayed binding that was moderate (SFLAP1, 4.1%–12.1%) to marginal (SFFN1, SFTNC, and SFVTN, <1%) and were therefore excluded from further analysis. Notably, SFLAP3 exhibited improved affinity for αvβ6 integrin (mean half-maximal inhibitory concentration, 3.5 nM; dissociation constant, 7.4). Moreover, small-animal PET imaging and biodistribution studies of HNSCC xenograft mice revealed an increased tumor-specific accumulation 30–60 min after injection of 68Ga-labeled or 177Lu-labeled DOTA-SFLAP3. Peptide staining further demonstrated binding specificity for SFLAP3 to HNSCC tumor cells. Finally, PET/CT scanning of an HNSCC patient showed specific SFLAP3 accumulation in the primary tumor lesion (SUVmax, 5.1) and in corresponding lymph node metastases (SUVmax, 4.1). Conclusion: SFLAP3 represents a promising tracer for prognostic assessment, diagnostic imaging, and possibly targeted therapy of αvβ6 integrin–expressing tumors.
Integrins comprise a large family of cellular adhesion receptors that play an important role in development, immune response, and cancer (1–3). Unlike most epithelial integrins, αvβ6 integrin is upregulated in many carcinomas associated with a poor prognosis, including lung, ovarian, pancreatic, colorectal, and cervical (4–8). In contrast, healthy tissues show no or a rather low αvβ6 integrin expression (9). Therefore, αvβ6 integrin is considered a prognostic biomarker and an important target for imaging and anticancer therapies (10,11). Using a sunflower trypsin inhibitor 1 (SFTI1)–based phage display library for biopanning against head and neck squamous cell carcinoma (HNSCC) cells and fractionated high-pressure liquid chromatography–derived membrane proteins, we identified the novel αvβ6 integrin–specific peptide SFITGv6 (12). Because of the molecular scaffold of SFTI1, a 14-residue cyclic miniprotein isolated from sunflower seeds that is stabilized by a single disulfide bridge, the peptide exhibits excellent stability (13). SFITGv6 demonstrates high binding affinity to several αvβ6 integrin–positive tumor cells and to HNSCC xenografts. Moreover, it accumulates in tumor lesions of HNSCC and non–small cell lung carcinoma but not in inflammatory lesions of cancer patients (12).
αvβ6 integrin binds the tripeptide recognition sequence arginine-glycine-aspartic acid (RGD) of several extracellular matrix proteins, including fibronectin, vitronectin, tenascin C, and latency-associated peptides (LAP) 1 and 3, which participate in carcinogenesis by promoting cell migration and invasion of cancer cells (14,15). Particularly, αvβ6 integrin–mediated transforming growth factor β (TGF-β) activation plays an important role in epithelial-to-mesenchymal transition and thereby promotes tumor growth and metastasis (16). Noncovalently associated with the N terminus of TGF-β1 and TGF-β3, LAP prevents TGF-β from binding to TGF-β receptors. Binding of LAP1 or LAP3 to αvβ6 integrin induces dissociation of the complex followed by TGF-β activation (17,18). Considering both the requirement for additional tumor-specific peptides targeting epithelial tumors and the high stability of the SFTI1 scaffold (13), we designed further RGD sequence–containing SFTI1 derivates (SFTI1-based peptides containing RGD motif–spanning octamers of fibronectin [SFFN1], tenascin C [SFTNC], vitronectin [SFVTN], LAP1 [SFLAP1], and LAP3 [SFLAP3]) and compared their αvβ6 integrin–specific binding properties with those of SFITGv6. Thus, we discovered SFLAP3, a peptide that demonstrates high αvβ6 integrin affinity and specificity for several αvβ6 integrin–expressing tumor cells. Moreover, SFLAP3 displays an increased accumulation in HNSCC xenografts and patient tumors.
MATERIALS AND METHODS
Cell Lines, Tumor Material, and Ethical Approval
The HNSCC-derived cell lines HNO97, HNO199, HNO210, HNO223, and HNO399 (19) were cultured in RPMI 1640 medium supplemented with 10% fetal calf serum at 37°C and 5% CO2 and were negative for Mycoplasma contamination. Uniqueness was assessed by the German Collection of Microorganisms and Cell Culture. Patient-derived tumor tissues obtained intraoperatively were snap-frozen and stored at −80°C. Written informed consent was obtained from all patients according to the research proposals approved by the Institutional Review Board at the Medical Faculty of the University of Heidelberg.
In Vitro Binding Experiments
For assessment of peptide binding capacity in vitro, 2.5–4 × 105 cells were seeded into 6-well plates and cultivated for 48 h. The cells were exposed to 125I-labeled peptides resuspended in 1 mL of serum-free medium for 10 or 60 min either with or without different concentrations of unlabeled peptide (10−5–10−10 M). Unbound peptide was washed off (3 washes with phosphate-buffered saline), and finally, cells were harvested using 0.3 M NaOH. To evaluate kinetic properties, the cells were exposed to 177Lu-DOTA–labeled peptides for 10, 30, 60, 120, 360, and 480 min, respectively, and harvested using 0.3 M NaOH. Efflux was assessed by incubating the cells with 177Lu-DOTA–labeled peptides for 60 min. Next, radioactive peptide-containing medium was replaced, and the cells were incubated for another 10, 30, 60, 120, and 240 min and finally harvested using 0.3 M NaOH. To assess peptide internalization, the cells were exposed to 177Lu-DOTA–labeled peptides for 10, 30, 60, 120, and 240 min at 37°C. Surface-bound peptides were removed by 10 min of incubation with 1 M glycine-HCl solution (pH 2.2). The cells were rinsed with phosphate-buffered saline and lysed as described. Radioactivity (MBq) was determined using a γ-counter (LB951G; Berthold Technologies) and calculated as percentage of applied injected dose (%AD) per 106 cells.
Surface Plasmon Resonance Assay
Binding affinity was assessed via a BiaCore X100 system (GE Healthcare) as described in detail previously (12). Briefly, to prevent integrin heterodimer denaturation during immobilization, αvβ6 integrin (1, 2, 3, and 4 μg/mL) and αvβ3 integrin (15, 30, 35, and 40 μg/mL) were used as analytes. SFLAP3 was dissolved in HBS-EP (HEPES-buffered saline, containing EDTA and Surfactant P20) running buffer and immobilized (loading level, 12 response units) on the C1 sensor chip (both GE Healthcare) using a manual amine-coupling protocol. Next, the binding affinity of αvβ6 and αvβ3 integrin, dissolved in HBS-EP running buffer (1–50 μg/mL), to SFLAP3 was measured (flow rate, 30 μL/min). The dissociation constant (KD) was determined by a 1:1 Langmuir model fit of the surface plasmon resonance sensograms.
Animal Experiments
Animal experiments were conducted in compliance with the German animal protection laws. For in vivo small-animal PET imaging and organ distribution studies, 8-wk-old BALB/c nude mice (Charles River Laboratories) were inoculated subcutaneously at the right shoulder with 5 × 106 tumor cells in Matrigel (BD Bioscience).
Xenografts were grown to a 10- to 15-mm diameter. The mice were anesthetized using isoflurane inhalation, and a 100-μL phosphate-buffered saline solution containing 68Ga-DOTA-SFLAP3 (HNO97, 37.9 MBq; HNO399, 26 MBq; HNO223, 27 MBq) or 68Ga-DOTA-SFITGv6 (HNO97, 30 MBq; HNO399, 34 MBq; HNO223, 34 MBq) was injected into the tail vein. Three-dimensional PET images were captured (Siemens Inveon PET scanner) as previously described (12). To assess the biodistribution, 100 μL of a 20 nM 177Lu-DOTA-SFLAP3 or 177Lu-DOTA-SFITGv6 solution (1 MBq) were administered as a bolus injection into the tail vein. At each time point (30, 60, 120, 240, 360 min), we sacrificed 3 animals, collected peripheral blood and the respective organs, weighed the tissues, and measured the radioactivity using a γ-counter. Radioactivity (MBq) was expressed as percentage of injected dose (%ID) per gram of tissue.
Peptide Staining
Histochemical staining was performed using biotinylated polyethylene glycol 12–labeled peptides on acetone-fixed cryosections (5 μm) derived from snap-frozen HNSCC tissues as described previously (12). Briefly, to prevent background staining, HNSCC cryosections were preincubated with an SP-2001 avidin/biotin blocking kit (Vector Laboratories), and 10−5 M peptide diluted in Dako Antibody Diluent (Agilent) was subsequently applied at 4°C overnight. Detection of bound peptide was assessed using the Vectastain Elite ABC Kit (PK-6100; Vector Laboratories) according to the manufacturer’s instructions. Peptide-binding specificity was ensured by using a peptide containing a scrambled (GRD) sequence and by incubation without peptide. Because of the instability of the peptide binding, staining was assessed on the same day as the binding experiment.
PET/CT Scanning of HNSCC Tumor Patient
An unenhanced PET/CT scan of a patient was obtained 60 and 180 min after intravenous injection of the 68Ga-DOTA–labeled peptide using a Biograph mCt Flow PET/CT scanner (Siemens Medical Solution) and the following parameters: slice thickness of 5 mm, increment of 3–4 mm, soft-tissue reconstruction kernel, and CARE Dose4D (Siemens). Immediately after CT scanning, a whole-body PET scan was acquired in 3 dimensions (matrix, 200 × 200) using FlowMotion continuous-bed-motion technology (Siemens) at 0.7 cm/min. The emission data were corrected for randoms, scatter, and decay. Images were reconstructed using ordered-subsets expectation maximization with 2 iterations and 21 subsets and were Gauss-filtered to a transaxial resolution of 5 mm in full width at half maximum. Attenuation correction was performed using the low-dose unenhanced CT data. Uptake was quantitatively assessed using a region-of-interest technique and SUVs.
RESULTS
Binding of SFTI1 Derivates to αvβ6 Integrin–Expressing HNSCC Cell Lines
RGD motif–containing octamers of the natural αvβ6 integrin ligands fibronectin, tenascin C, vitronectin, LAP1, and LAP3 (Supplemental Table 1; supplemental materials are available at http://jnm.snmjournals.org) were grafted between Thr4 and Cys11 into the binding loop of the SFTI1 scaffold. First, the binding properties of 125I-labeled SFFN1, SFTNC, SFVTN, SFLAP1 SFLAP3, and SFITGv6 were compared using 5 HNSCC cell lines differing in their αvβ6 integrin expression as assessed by flow cytometry (Supplemental Fig. 1). As shown in Figure 1A, 125I-SFLAP3 displayed the highest binding capacity to all tested HNSCC cell lines (HNO97, 24.7%; HNO399, 10.1%; HNO199, 19.7%; HNO223, 17.6%; HNO210, 10.6%)—higher even than that of 125I-SFITGv6 (HNO97, 21.8%; HNO399, 9.8%; HNO199, 16.8%; HNO223, 13.5%; HNO210, 10.5%). In contrast, only a moderate binding capacity was detected for SFLAP1 (4.1%–12.1%) and almost none (<1%) for SFFN1, SFTNC, and SFVTN (Fig. 1A). In summary, SFLAP3 and SFITGv6 demonstrated the highest binding capacity and, thus, were further investigated comparatively.

(A) Binding of 125I-labeled SFLAP3, SFITGv6, SFLAP1, SFTNC, SFFN1, and SFVTN to different HNSCC cells after exposure to peptides for 60 min. Bars represent %AD/106 cells (HNO97, HNO399, HNO223, HNO210, and HNO199). Increasing concentrations (10−5–10−10 M) of respective unlabeled peptide were used to compete for binding of 125I-labeled SFLAP (B) and SFITGv6 (C) to HNSCC cell lines HNO97, HNO399, and HNO223 (%AD/106 cells). Data are mean and SD of triplicate measurements from representative experiment. (D) Affinity of αvβ6 integrin (applied concentrations: 1, 2, 3, and 4 μg/mL) for immobilized SFLAP3 (loading level: 12 response units) was measured by surface plasmon resonance assay (n = 3) with flow rate of 30 mL/min. Surface plasmon resonance sensograms were evaluated with BiaCore evaluation software (black curves) and correspond to experimentally fitted red curves. KD was determined by 1:1 Langmuir model fit of surface plasmon resonance sensograms. IC50 = half-maximal inhibitory concentration.
SFLAP3 Affinity for αvβ6 Integrin and Serum Stability
To assess the specificity of the peptide, binding competition experiments were performed. In all HNSCC cell lines, binding of 125I-SFLAP3 and 125I-SFITGv6 could almost be completely inhibited by adding 10−6 M unlabeled counterparts as competitor (Figs. 1B and 1C). However, SFLAP3 exhibited higher affinity than SFITGv6 (mean half-maximal inhibitory concentrations, 3.5 and 14.11 nM, respectively). Concordantly, the surface plasmon resonance assay revealed that SFLAP3 had a higher affinity than SFITGv6 for αvβ6 integrin (KD = 7.4 and 14.8 nM, respectively) (Fig. 1D) (12). In contrast, SFLAP3 had a 10-fold lower affinity to αvβ3 integrin (KD = 167 nM) (Supplemental Fig. 2A), further ensuring the αvβ6 integrin specificity of SFLAP3. Additionally, SFLAP3 showed high proteolytic stability, with no degradation over 24 h (Supplemental Fig. 2B), demonstrating suitability for in vivo applications.
Comparative Evaluation of Kinetics, Internalization, and Efflux
Considering time-dependent deionization of 125I, the determination of kinetics, internalization, and efflux was performed using 177Lu-DOTA–labeled peptides. Kinetic experiments revealed a continuous increase in peptide uptake over the whole period of measurement for both peptides. Yet, 177Lu-DOTA-SFLAP3 exhibited a lower maximum binding capacity (HNO97, 18.5%; HNO223, 9.66%; HNO399, 4.31%) (Supplemental Fig. 3A) than 177Lu-DOTA-SFITGv6 (HNO97, 22.15%; HNO223, 14.45%; HNO399, 5.85%) (Supplemental Fig. 3B). Accordingly, within 60 min after exposure to HNO97, we measured an increase in 177Lu-DOTA-SFITGv6 binding of up to 16.85% and a fast internalization of up to 70% of the total bound peptide (Fig. 2B), but up to 11.14% binding and 60% internalization of 177Lu-DOTA-SFLAP3 (Fig. 2A). For both peptides, similar trends regarding a time-dependent increase in peptide binding and internalization were observed after exposure to HNO399 (Supplemental Figs. 4A and 4B) and HNO223 (Supplemental Figs. 4C and 4D), although the amount of internalized peptide slightly decreased (>45%). Concordantly, an efflux of more than 50% of the originally accumulated SFLAP3 and SFITGv6 within 120 min after termination of the uptake was measured (Figs. 3A and 3B). Notably, for 177Lu-DOTA-SFLAP3, we observed a moderate renewed increase in binding and internalization after 240 min (Fig. 3A), possibly indicating peptide reuptake from the culture medium.

Uptake (gray bars) and internalization of 177Lu-DOTA–labeled SFLAP3 (A) and SFITGv6 (B) in HNO97 cells. After incubation for 10, 30, 60, 120, and 240 min, radioactivity in lysates was determined and calculated as %AD/106 cells. Data are mean and SD of triplicate measurements from representative experiment.
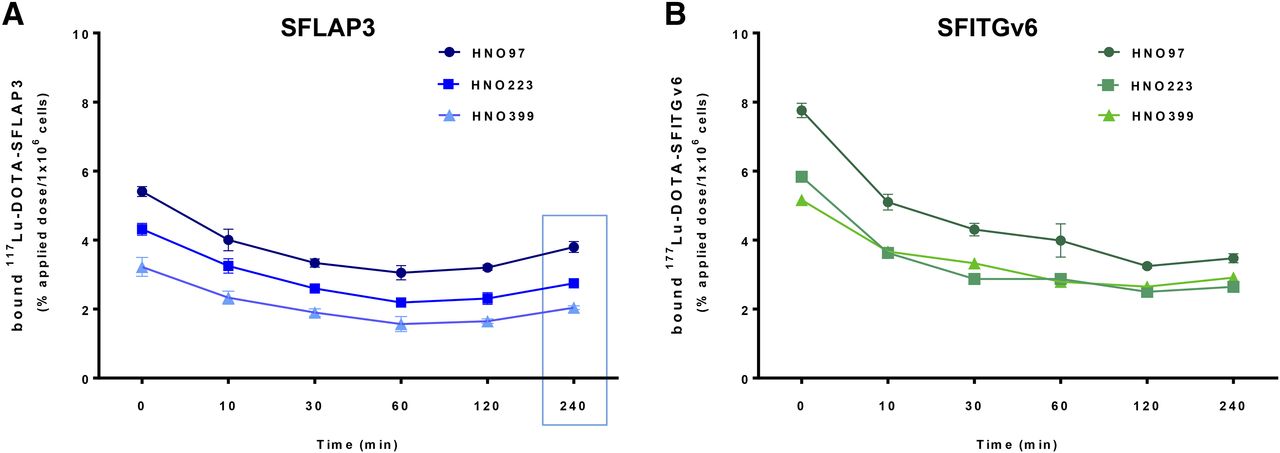
Time-dependent efflux of SFLAP3 (A) and SFITGv6 (B) from HNSCC cell lines. HNO97, HNO399, and HNO223 cells were exposed to 177Lu-DOTA–labeled peptides for 60 min. After replacement of culture medium, radioactivity in cell lysates was measured immediately and after 10, 30, 60, 120, and 240 min and calculated as %AD/106 cells. Data are mean and SD of triplicate measurements from representative experiment.
In Vivo Targeting Properties of SFLAP3 and SFITGv6
To compare the in vivo targeting properties of the peptides, small-animal PET imaging of HNO97, HNO399, and HNO223 xenografts was performed using 68Ga-DOTA–labeled SFLAP3 and SFITGv6 as radiotracer. In HNO97 tumor lesions, a fast and continuous accumulation of 68Ga-DOTA-SFLAP3 (SUVmean, 0.63) (Fig. 4A; Supplemental Fig. 5A) and 68Ga-DOTA-SFITGv6 (SUVmean, 0.68) (Supplemental Figs. 5B and 5C) was measured within 60 min after injection and stayed for at least 140 min (Supplemental Figs. 5D and 5E). Nonspecific peptide activity cleared quickly from the blood, resulting in images with excellent tumor-to-background ratios. Accordingly, the biodistribution of 177Lu-DOTA-SFLAP3 in tumor lesions and individual organs of HNO97 xenografts revealed maximal peptide accumulation in the tumors after 60 min (9.1 ± 1.2 %ID/g), followed by a decrease to 5.80 ± 1.44 %ID/g at 360 min after injection (Fig. 4B). In contrast, the maximal uptake of 177Lu-DOTA-SFITGv6 in HNO97 tumors (6.73 ± 1.00 %ID/g) was already present at 30 min after injection, and uptake decreased to 2.43 ± 0.51 %ID/g after 360 min (Supplemental Fig. 5F). Small-animal PET imaging of HNO399 (Fig. 4C; Supplemental Fig. 6B) and HNO223 xenografts (Supplemental Figs. 7A and 7B) revealed less accumulation of 68Ga-DOTA-SFLAP3 (SUVmean, 0.44 for HNO399 and 0.32 for HNO223) (Supplemental Figs. 6A and 7C) and of 68Ga-DOTA-SFITGv6 (SUVmean, 0.33 for HNO399 and 0.23 for HNO223) (Supplemental Figs. 6C and 7D). Accordingly, we measured a maximal 177Lu-DOTA-SFLAP3 accumulation of 3.6 %ID/g (Fig. 4D) and 3.4 %ID/g (Supplemental Fig. 7E) in HNO399 and HNO233 xenografts, respectively, at 60 min after injection, followed by a decrease to almost 2% at 360 min. Less than 1 %ID/g was detected in the blood and most tissues of xenograft mice 120 min after injection of either peptide, accounting for tumor-to-tissue ratios predominantly above 1 (Supplemental Tables 2A–2D). However, in the kidneys we observed a constant high SFLAP3 retention (8–14 %ID/g), which was indeed clearly reduced in comparison to SFITGv6 (Supplemental Fig. 5F).

Small-animal PET imaging of mice bearing HNO97 (A) and HNO399 (C) xenografts 60 min after injection of 68Ga-DOTA-SFLAP3. Biodistribution of 177LU-DOTA-SFLAP3 in tumor and organs of HNO97 (B) and HNO399 (D) tumor–bearing mice was measured 30, 60, 120, 240, and 360 min after injection and calculated as %ID/g. Data are mean of 3 per time point and SD. MIP = maximum-intensity projection.
In Situ Binding Properties of SFLAP3
In analogy to the previous experiments (12), we compared the binding specificity of the peptides to sections of HNO97 xenografts and patient-derived HNO399 and HNO210 tumors by peptide histochemical staining using biotin-labeled SFLAP3 and SFITGv6. Although SFLAP3 showed a slightly fainter staining, both peptides revealed tumor cell–specific binding, whereas lower peptide binding was observed at the stromal compartments (Fig. 5). In contrast, no binding of the GRD sequence containing peptide was observed, further corroborating the specificity of the SFLAP3 peptide.

Histochemical peptide staining of HNSCC cyrosections of HNO97 xenografts and of patient-derived HNO399 and HNO210 tumor sections performed with 10−5 M biotinylated polyethylene glycol 12–labeled SFLAP3 and SFITGv6. As control, scrambled SFLAP3 derivate containing GRD was applied. Scale bar = 50 μm.
SFLAP3 Accumulation in HNSCC Lesions and Lymph Node Metastases
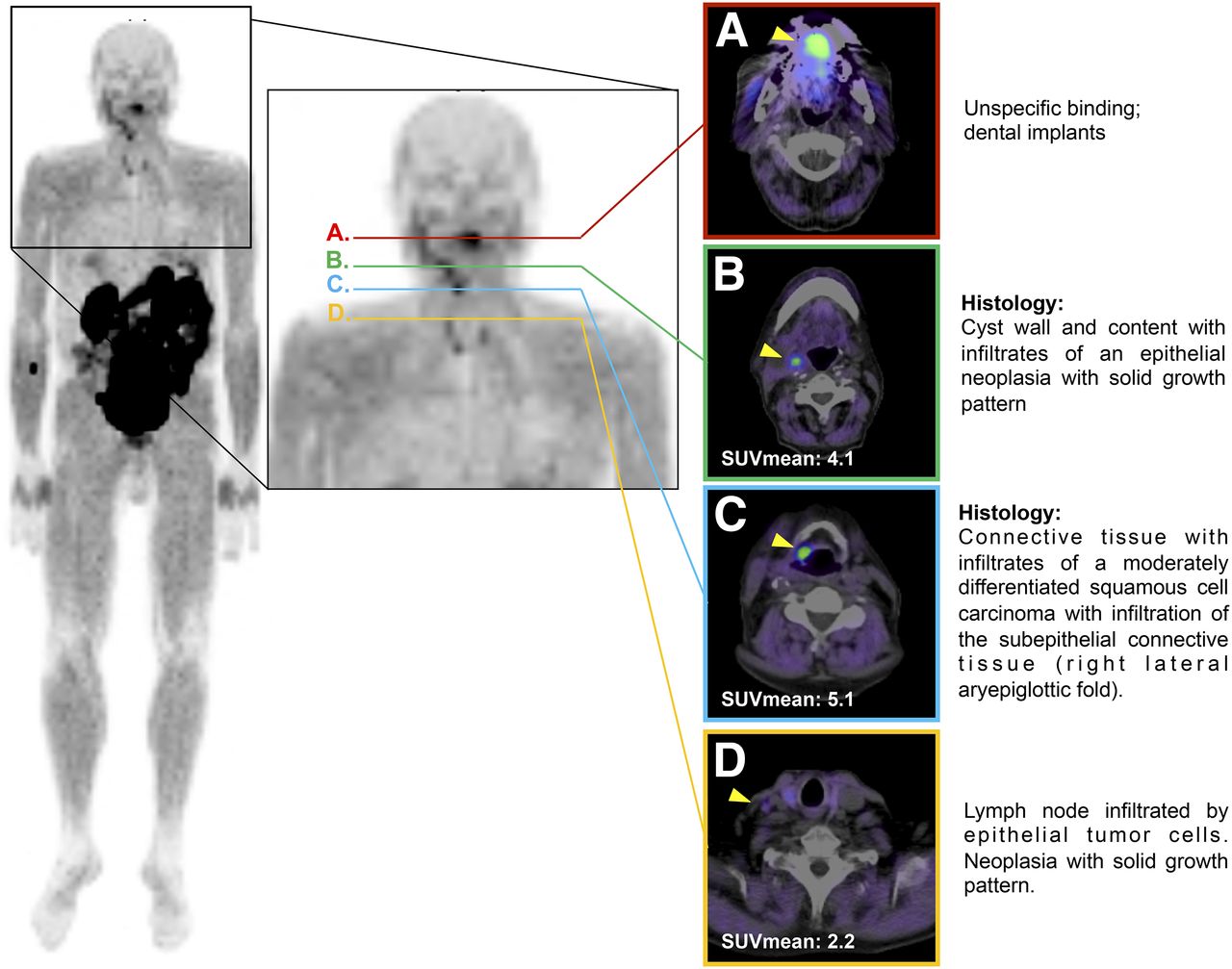
Finally, we investigated the therapeutic or diagnostic suitability of SFLAP3, using the example of a 79-y-old male carcinoma patient who presented with lymph node swelling in the right cervical area, assuming the diagnosis cancer of unknown primary. To clarify the diagnosis, 322 MBq of 68Ga-DOTA–labeled SFLAP3 were administered and PET/CT performed. After 60 min, we observed a tumor-specific accumulation of SFLAP3 at the primary tumor lesion (right site of aryepiglottic folds; median SUVmax, 5.1) (Fig. 6) and at the right cervical area (SUVmax, 4.1) (Supplemental Table 3). The histopathologic examination of the surgically resected respective tissues proved the presence of level II/III squamous cell carcinoma and corresponding lymph node metastases at the respective locations. The correlation of PET findings and histopathologic findings revealed the primary tumor lesion to be subepithelial connective tissue infiltrated by moderately differentiated squamous cell carcinoma (Fig. 6C). The tumor lesion at the right cervical area was classified as cyst wall and content; histopathologic analysis showed infiltrates of an epithelial neoplasia with a solid growth pattern (Fig. 6B). Moderate tracer accumulation was also seen in a lymph node in the right cervical area (SUVmax, 2.2), which was histopathologically classified as a lymph node infiltrated by epithelial tumor cells and neoplasia with a solid growth pattern (Fig. 6D).

Transaxial PET/CT scans of HNSCC tumor patient after application of 322 MBq of 68Ga-DOTA-SFLAP3, with arrowheads indicating peptide accumulation. (A) Level showing unspecific binding. (B) Level showing cyst wall and content. (C) Level showing primary tumor. (D) Level showing lymph node infiltration.
DISCUSSION
SFTI1 has already been proven to be a suitable scaffold for the development of extremely stable peptides targeting tumor-associated receptors, including the αvβ6 integrin–specific peptide SFITGv6 (12,20). αvβ6 integrin has been shown to be expressed in up to 99.9% of HNSCC (9,14), as well as in lung (4), colon (7), breast (21), and pancreas carcinoma (6), and is often associated with a poor prognosis (9). Thus, αvβ6 integrin represents an important target for imaging and therapy for a variety of epithelial malignancies (22–25). To design novel target-specific peptides, we introduced the RGD-containing octamer of natural αvβ6 integrin ligands resembling the RGDLXXL sequence into the SFTI1 scaffold and compared the specificity and affinity of the novel peptides with that of SFITGv6.
In comparing the binding properties of these SFTI1 derivates, we identified the superiority of the αvβ6 integrin–specific peptide SFLAP3, containing the amino acid sequence GRGDLGRL. Comparative analysis of SFLAP3 and SFITGv6 revealed improved binding (>20%) of the novel peptide to several HNSCC cell lines corresponding to their αvβ6 integrin expression profile. Considering the importance of the RGDLXXL motif—in particular, the DLXXL sequence—for αvβ6 integrin binding affinity and specificity (26–28), it is not surprising that SFLAP3 provides significant binding properties. In contrast, for SFLAP1, SFFN1, SFVTN, and SFTNC—partially or completely lacking the LXXL motif—we observed less or even almost no binding. Although being physiologically involved in αvβ6 integrin–dependent promotion of migration, invasion, and signal transduction pathways (14), the RGD motif–containing sequences of LAP1, fibronectin, vitronectin, and tenascin C incorporated into the SFTI1 scaffold might not allow for αvβ6 integrin affinity and specificity in vitro.
As expected from stability analysis of the naturally occurring SFTI1 (13) and the SFTI1 derivates DLL4 (20) and SFITGv6 (12), SFLAP3 also demonstrated marked stability in human serum over 24 h. Additionally, an improved affinity of SFLAP3 (KD = 7.4 nM) for αvβ6 integrin when compared with SFITGv6 was measured. Subsequent investigation of 177Lu-DOTA–labeled SFLAP3 and SFITGv6 regarding kinetics, internalization, and efflux revealed a continuous increase in SFLAP3 binding (≤18.5% within 240 min) and fast internalization (>60%) followed by a decrease (<50%). Correspondingly, we observed greater than 50% efflux and almost 50% retention of the originally accumulated peptide over 120 min, justifying further in vivo evaluation. These experiments also demonstrated a consistently higher binding (≤5.7%) of 177Lu-DOTA–labeled SFITGv6 than of SFLAP3. It is thought that the chemical properties of the SFLAP3 and SFITGv6 amino acid composition influence the binding capacity of DOTA–labeled peptides differently.
On the basis of the excellent tumor-to-background ratios, we were able to selectively image HNSCC xenografts using small-animal PET at 60–140 min after the administration of 68Ga-DOTA–labeled SFLAP3 and SFITGv6. Unbound and nonspecifically bound peptide rapidly cleared from the blood and the surrounding tissues, except for the kidneys. As expected from the αvβ6 integrin expression profile of HNSCC cell lines, we detected a rather low but clearly visible accumulation of both 68Ga-DOTA-SFLAP3 (SUVmean, 0.63) and 68Ga-DOTA-SFITGv6 (SUVmean, 0.68) in HNO97 xenografts. In contrast, HNO399 and HNO223 xenografts showed a lower uptake of the 68Ga-DOTA–labeled peptides but a generally higher affinity of 68Ga-DOTA-SFLAP3 (SUVmean, 0.44; SUV, 0.32) than did 68Ga-DOTA-SFITGv6 (SUVmean, 0.32; SUVmean, 0.23). The biodistribution of 177Lu-DOTA–labeled peptides in HNSCC xenograft mice confirmed the improved affinity of SFLAP3 for αvβ6 integrin–expressing tumors. In HNO97 tumors, we found a marked accumulation of SFLAP3 (≤9.10 %ID/g) and SFITGv6 (6.73 %ID/g), compared with that in healthy tissues, even after 240 and 360 min. In contrast, less than 4 %ID/g of SFLAP3 was measured for HNO399 and HNO223, emphasizing the importance of pronounced target expression for selective in vivo tumor imaging and targeting. As expected from a previous biodistribution analysis of αvβ6 integrin binding peptides in HNO97 (12) and BxPC-3 xenografts (11), a remarkable but slightly decreasing accumulation of 177Lu-DOTA-SFLAP3 was noticed in the kidneys of HNSCC xenografts. Although the renal retention of SFLAP3 was clearly lower than that of SFITGv6, we can recommend that the pharmacokinetic properties of the peptide be improved.
Even though peptide-based histochemical staining revealed a moderate but tumor cell–specific binding of SFLAP3 to HNO97 xenografts and patient-derived HNSCC tumor cyrosections, the 68Ga-DOTA–labeled peptide was successfully used as a radiotracer for PET/CT imaging of a patient with a carcinoma of unknown primary, leading to the discovery of a histologically proven HNSCC and a corresponding lymph node metastasis. In line with the small-animal PET imaging results for HNSCC xenografts, the SUVs in the patient persisted from 60 min (SUVmax, 5.1) to 180 min (SUVmax, 4.6) after injection, demonstrating the diagnostic suitability of the peptide.
In analogy to the 68Ga-DOTA-SFITGv6 PET/CT findings for patients with HNSCC and non–small cell lung carcinoma (12), 68Ga-DOTA-SFLAP3 PET/CT revealed peptide accumulation in the kidneys, secretion into the gastrointestinal tract, and intraluminal transport to the terminal ileum and the cecum. Thus, when SFLAP3 is being considered for imaging of αvβ6 integrin–positive carcinomas occurring below the diaphragm, administration of laxatives (reducing intrabowel activity and avoiding a high background signal) and improvement of the pharmacokinetic properties of the peptide are required.
CONCLUSION
We found that the αvβ6 integrin–binding peptide SFLAP3 provides improved affinity and excellent binding properties for αvβ6 integrin–expressing HNSCC tumors in vitro and in vivo. Thus, SFLAP3 represents a promising tracer for both diagnostic imaging and targeted therapy of αvβ6 integrin–expressing tumors.
DISCLOSURE
This project was supported by the Deutsche Forschungsgemeinschaft (DFG) (HA 2901/12-1). No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Vanessa Kohl and Marlene Tesch for providing technical assistance, Ursula Schierbaum and Karin Leotta for performing animal experiments, Dr. Esther Herpel (Tissue Bank, National Center for Tumor Diseases (NCT), Heidelberg, Germany) for examining the tumor tissues histopathologically, and Dr. Carmen Rapp for proofreading the manuscript.
Footnotes
Published online Apr. 19, 2018.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication February 15, 2018.
- Accepted for publication March 28, 2018.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Clinical Translation of a 68Ga-Labeled Integrin {alpha}v{beta}6-Targeting Cyclic Radiotracer for PET Imaging of Pancreatic Cancer
- A Conversation Between Uwe Haberkorn and Johannes Czernin
- Preclinical Development and First-in-Human Imaging of the Integrin {alpha}v{beta}6 with [18F]{alpha}v{beta}6-Binding Peptide in Metastatic Carcinoma
- Using backbone-cyclized Cys-rich polypeptides as molecular scaffolds to target protein-protein interactions