


Abstract
The addition of ezetimibe, an intestinal cholesterol absorption inhibitor, to statin therapy has recently shown clinical benefits in the Improved Reduction of Outcomes: Vytorin Efficacy International Trial by reducing low-density-lipoprotein (LDL) cholesterol levels more than statin therapy alone. Here, we investigated the mechanisms by which inhibition of intestinal cholesterol absorption might contribute to the clinically observed reduction in cardiovascular events by evaluating its effect on inflammatory plaque development in apolipoprotein E−/− mice. Methods: Apolipoprotein E−/− mice were fed the Paigen diet (1.25% cholesterol, 0.5% cholic acid, and 15% fat) without or with ezetimibe (7 mg/kg/d) for 6 wk. In a first set of mice (n = 15), we intravenously injected 3H-cholesteryl oleate–labeled human LDL to test whether ezetimibe promotes LDL-derived cholesterol fecal excretion. In a second set (n = 20), we used the imaging agent 99mTc-cAbVCAM1–5 to evaluate expression of an inflammatory marker, vascular cell adhesion molecule 1 (VCAM-1), in atherosclerotic plaques. In a third set (n = 21), we compared VCAM-1 expression with 99mTc-cAbVCAM1–5 uptake in various tissues. Results: Mice treated with ezetimibe showed a 173% higher LDL–cholesteryl ester plasma disappearance rate (P < 0.001 vs. control) after 3H-cholesteryl oleate–labeled LDL injection. At 96 h after injection, the hepatic fraction of 3H-tracer was 61% lower in mice treated with ezetimibe (P < 0.001). Meanwhile, LDL-derived 3H-cholesterol excretion in the feces was 107% higher (P < 0.001). The antiatherogenic effect of ezetimibe monitored by 99mTc-cAbVCAM1–5 SPECT showed a 49% reduction in aortic tracer uptake (percentage injected dose per cubic centimeter, 0.95 ± 0.04 vs. 1.87 ± 0.11; P < 0.01). In addition to hypercholesterolemia, the proinflammatory Paigen diet significantly increased VCAM-1 expression with respect to the control group in various tissues, including the aorta, and this expression correlated strongly with 99mTc-cAbVCAM1–5 uptake (r = 0.75; P < 0.05). Conclusion: Inhibition of intestinal cholesterol absorption with ezetimibe promotes antiatherosclerotic effects through increased LDL cholesterol catabolism and LDL-derived cholesterol fecal excretion and reduces inflamed atherosclerotic plaques. These mechanisms may contribute to the benefits of adding ezetimibe to a statin therapy.
Cardiovascular diseases are caused mainly by coronary artery disease and, more specifically, by rupture of a vulnerable atherosclerotic plaque (1). To prevent cardiovascular events, lowering low-density-lipoprotein (LDL) cholesterol levels with statins is the most widely used therapy (2). Nevertheless, some patients treated with statins are not reaching low-enough levels of LDL cholesterol or experience adverse effects (3), calling for alternative LDL-lowering therapies.
Ezetimibe is a dietary and biliary intestinal cholesterol absorption inhibitor that selectively inhibits the Niemann-Pick C1-like 1 receptor without affecting intestinal absorption of triglycerides (4). Davis et al. demonstrated that ezetimibe decreased plasma cholesterol levels by 61% and reduced the development of atherosclerotic lesions in apolipoprotein E−/− (ApoE−/−) mice (5). Ezetimibe is prescribed as a second-line therapy (6), and its effectiveness for reducing LDL cholesterol levels has been demonstrated in several clinical studies (7,8). Moreover, a beneficial 22% reduction in LDL cholesterol levels has been reported when ezetimibe was given together with a statin, as compared with a statin alone (9). Results from a clinical trial, Improved Reduction of Outcomes: Vytorin Efficacy International Trial, that started in 2005 showed that reducing LDL cholesterol levels below the recommended target by adding ezetimibe to simvastatin treatment granted an additional benefit by significantly reducing cardiovascular events, as compared with treatment using simvastatin alone, in subjects with acute coronary syndrome (10). Interestingly, ezetimibe in combination with a statin significantly decreased the levels of C-reactive protein and might have metabolic effects other than simply the lowering of LDL cholesterol levels (11,12).
In the present study, we aimed to investigate the mechanisms by which ezetimibe might reduce cardiovascular events. We investigated whether blocking intestinal cholesterol absorption may promote fecal excretion of atherogenic cholesterol deriving from LDL particles. The effect of ezetimibe on LDL cholesterol metabolism and LDL-derived cholesterol fecal excretion was therefore evaluated with 3H-cholesteryl oleate–labeled LDL kinetics, and its effect on inflammatory plaque development was evaluated using 99mTc-cAbVCAM1–5 imaging in ApoE−/− mice fed the Paigen diet. 99mTc-cAbVCAM1–5, currently under clinical translation, is a SPECT radiotracer directed against the inflammatory marker vascular cell adhesion molecule 1 (VCAM-1) (13,14). This molecular imaging agent belongs to a new class of radiotracer derived from single-domain antibodies, or variable domain of the heavy chain of the heavy-chain antibody, which are composed of the single variable domain of camelidae immunoglobulins. Aortic atherosclerotic lesions have been successfully detected in ApoE−/− mice fed a western diet using 99mTc-cAbVCAM1–5 imaging, and the sensitivity of this tool has been demonstrated using statin therapy as a gold standard (13,14). This molecular imaging agent can therefore be used for the preclinical noninvasive evaluation of antiatherosclerotic therapies.
MATERIALS AND METHODS
Animals and Diet
All experimental procedures were in accordance with institutional guidelines and approved by the animal care and use committees of Midi-Pyrénées and Grenoble University. Seven-week-old female mice were used. Fifty ApoE−/− mice were fed a proatherogenic Paigen diet (1.25% cholesterol, 0.5% cholic acid, and 15% fat) supplemented (Paigen+eze), or not (Paigen), with ezetimibe (0.005% in diet, equivalent to 7 mg/kg/d) for 6 wk. Six C57BL6/J mice were fed a chow diet to serve as a control. Body weight and food consumption were monitored weekly.
The effect of ezetimibe on LDL cholesterol metabolism and LDL-derived cholesterol fecal excretion in vivo was evaluated using 3H-cholesteryl oleate–labeled LDL kinetics (n = 15), and the effect of ezetimibe on atherosclerosis development and inflammation was investigated through in vivo SPECT and postmortem analysis (n = 41).
Effect on LDL Cholesterol Metabolism and Fecal Excretion
LDL particles were isolated from human plasma by ultracentrifugation (1.019 < density < 1.063), extensively dialyzed, and then labeled with 3H-cholesteryl oleate in the presence of lipoprotein-deficient serum isolated from human plasma, as described previously (15). After isolation of labeled LDL by ultracentrifugation and extensive dialysis, the mice (7 in the Paigen group and 8 in the Paigen+eze group) were weighed, individually caged for feces collection, and injected intravenously with 3H-cholesteryl oleate–labeled LDL (∼3–4 million disintegrations/min/mouse).
Blood was collected from the tail tip at 5 min and then 1, 3, 6, 24, and 48 h after intravenous injection to isolate plasma (5 μL) and count 3H-radioactivity. The plasma 3H-tracer decay curve was then used to calculate the LDL–cholesteryl ester fractional catabolic rate (i.e., the fraction of tracer irreversibly removed from the plasma per unit of time) using the SAAM II software program (The Epsilon Group), as described previously (16).
The mice were kept in individual cages for up to 96 h after tracer injection to collect feces and to measure fecal 3H-cholesterol and 3H-bile acids after chemical extraction, as described previously (15). The mice were then euthanized and exsanguinated. The liver was harvested and weighed. An approximately 50-mg liver sample was used to measure 3H-radioactivity in the whole liver as well as in the cholesterol and bile acid fractions. Hepatic and fecal total cholesterol and bile acid masses were also measured from the same samples using colorimetric kits, as described previously (17), and tracer recovery was expressed as percentage injected dose (%ID).
Effect on Atherosclerosis Development and Inflammation
In Vivo SPECT
In vivo SPECT of inflamed atherosclerotic lesions was performed using 99mTc-cAbVCAM1–5 in 10 Paigen mice and 10 Paigen+eze mice. cAbVCAM1–5 was radiolabeled as previously described using the tricarbonyl method (13). A SPECT/CT acquisition was performed 2 h after the intravenous injection of 47.2 ± 10.7 MBq of 99mTc-cAbVCAM1–5, and aortic uptake was expressed as %ID/cm3 (13).
Postmortem Analysis
Frozen sections of aortic root were obtained for histologic examination, VCAM-1 immunostaining, and 99mTc-cAbVCAM1–5 autoradiography. Plaque surface area was expressed as square millimeters. VCAM-1 intensity was graded as low (1), moderate (2), or strong (3) by 2 masked observers. 99mTc cAbVCAM1–5 uptake in autoradiographic images was expressed either as a concentration (%ID/g) or as the total uptake per slice (%ID/slice), as previously described (14).
VCAM-1 protein expression was further investigated by ELISA in 9 Paigen mice, 6 Paigen+eze mice, and 6 control mice. Three hours after injection of 99mTc-cAbVCAM1–5, the mice were euthanized by CO2 inhalation. Tissues of interest were rapidly harvested, rinsed in 0.9% NaCl, and weighed. 99mTc-cAbVCAM1–5 uptake was measured by γ-well counting (Wizard2; PerkinElmer). After the tissue had been processed with a Potter–Elvehjem tissue grinder in ice-cold radioimmunoprecipitation assay buffer, the homogenates were centrifuged at 10,000g for 10 min at 4°C. The supernatants were extracted and the VCAM-1 protein level was quantified using a commercial kit (sVCAM-1/CD-106; R&D Systems).
Western blot analysis was used to evaluate whether ezetimibe has a direct antiinflammatory effect on VCAM-1 expression in human THP1 monocytes. The human THP1 cell line was cultured in RPMI 1640 (Sigma-Aldrich) containing 10% heat-inactivated fetal bovine serum, supplemented with 2 mM glutamine and 1% penicillin/streptomycin at 37°C in an atmosphere of 5% CO2. A total of 1 × 106 cells per well were suspended in 6-well plates and differentiated into macrophages for 24 h with phorbol 12-miristate 13-acetate at 5 ng/mL in the presence or absence of ezetimibe (100 nM). Then, the cells were rinsed with RPMI 1640 and allowed to rest for 24 h. Differentiated THP1 macrophages were stimulated with TNF-α at 50 ng/mL for 24 h with or without ezetimibe treatment (100 nM). After treatment, the cells were rinsed with phosphate-buffered saline and lysed with ice-cold radioimmunoprecipitation assay buffer at −80°C. Cell lysates were centrifuged at 10,000g for 10 min at 4°C, and the supernatant was collected. Protein was quantified by the bicinchoninic acid method, and 30-μg protein samples were loaded into sodium dodecyl sulfate–polyacrylamide gel (7.5%) and then transferred on a nitrocellulose membrane. Rabbit anti–VCAM-1 antibody (ab13047; Abcam) was used as the primary antibody at 1/1,000 at 4°C overnight. Goat antirabbit (horseshoe peroxidase–labeled goat antirabbit IgG; Dako) was used as the secondary antibody at 1/2,000 at room temperature for 1 h. The protein signal, observed at 110 kDa, was quantified using ImageJ software and normalized to the expression of α-actin.
Statistical Analysis
All results are presented as mean ± SEM. Student t testing was used to compare datasets of equal variance, and Mann–Whitney U testing was used for unpaired datasets of unequal variance. The significance of linear correlations was assessed with Pearson testing. Multiple-comparison statistical testing was used to compare data among the 3 groups (Paigen, Paigen+eze, and control). The significance level was set at a P value of less than 0.05.
RESULTS
Effect on Plasma Cholesterol Levels and Body Weight
Treatment with ezetimibe significantly reduced total cholesterol levels in plasma (Paigen, 29.0 ± 3.1 mmol/L, and Paigen+eze, 14.6 ± 2.3 mmol/L; P < 0.01). Moreover, body weight was significantly higher in the ezetimibe-treated group than in the control group (21.0 ± 0.2 g vs. 18.4 ± 0.5 g, P < 0.01) (Supplemental Table 1; supplemental materials are available at http://jnm.snmjournals.org).
Effect on LDL Cholesterol Metabolism and Fecal Excretion
After injection of 3H-cholesteryl oleate–labeled human LDL, mice that were treated with ezetimibe showed a higher rate of 3H-tracer disappearance than mice that were not (Fig. 1A), resulting in a 173% higher (P < 0.001) LDL–cholesteryl ester catabolism (Fig. 1B). At 96 h after radiolabeled LDL injection, 3H-tracer recovery in the whole liver was reduced by 61% with ezetimibe (P < 0.001), and accordingly, the fractions of 3H-tracer recovery in hepatic cholesterol and hepatic bile acids were also significantly reduced, by 55% and 52%, respectively (Fig. 1C). Meanwhile, the fraction of 3H-tracer recovery in fecal cholesterol was increased by 107% (P < 0.001) (Fig. 1D). Similar trends were observed for hepatic cholesterol level (Fig. 1E) and fecal cholesterol mass excretion (Fig. 1F), with ezetimibe resulting in a 75% reduction and 99% increase, respectively (both P < 0.001).

Effect of ezetimibe on LDL cholesterol metabolism and excretion using in vivo 3H-cholesteryl: 3H-tracer plasma disappearance (A), LDL–cholesteryl ester fractional catabolic rate (B), hepatic (C) and fecal (D) 3H-tracer tracer recovery after 3H-cholesteryl oleate–labeled human LDL intravenous injection, and hepatic (E) and fecal (F) total cholesterol and bile acid mass in ApoE−/− mice. *P < 0.05 vs. Paigen group. **P < 0.01 vs. Paigen group. ***P < 0.001 vs. Paigen group.
Effect on Atherosclerosis Development and Inflammation
In Vivo SPECT
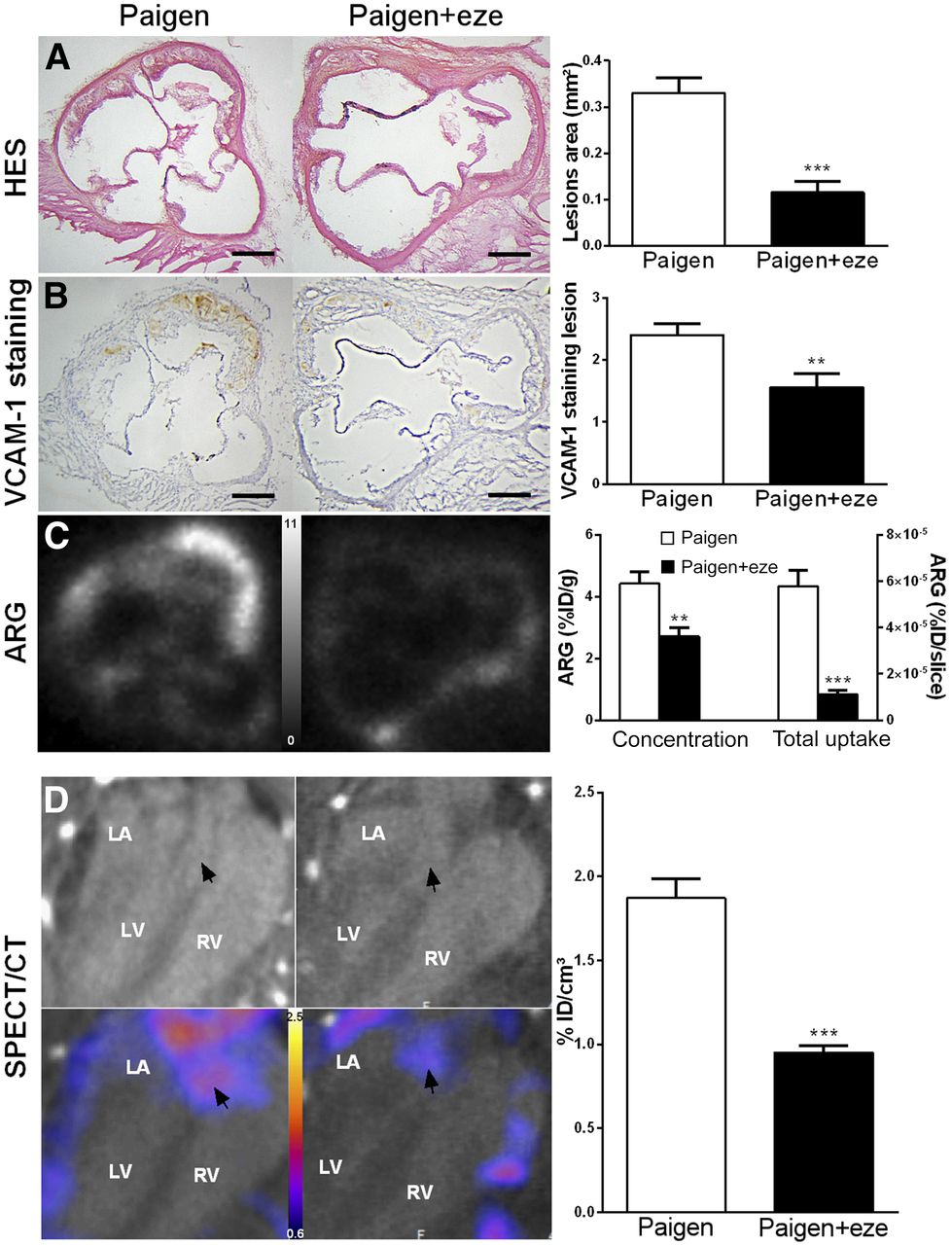
VCAM-1 expression was successfully evaluated noninvasively using in vivo 99mTc-cAbVCAM1–5 SPECT/CT to investigate the effect of ezetimibe on atherosclerosis-related inflammation. 99mTc-cAbVCAM1–5 uptake in the ascending aorta was visible in the Paigen group and was decreased by 49% when the Paigen diet was supplemented with ezetimibe (1.87 ± 0.11 %ID/cm3 vs. 0.95 ± 0.04 %ID/cm3, P < 0.001) (Fig. 2D; Supplemental Fig. 1).
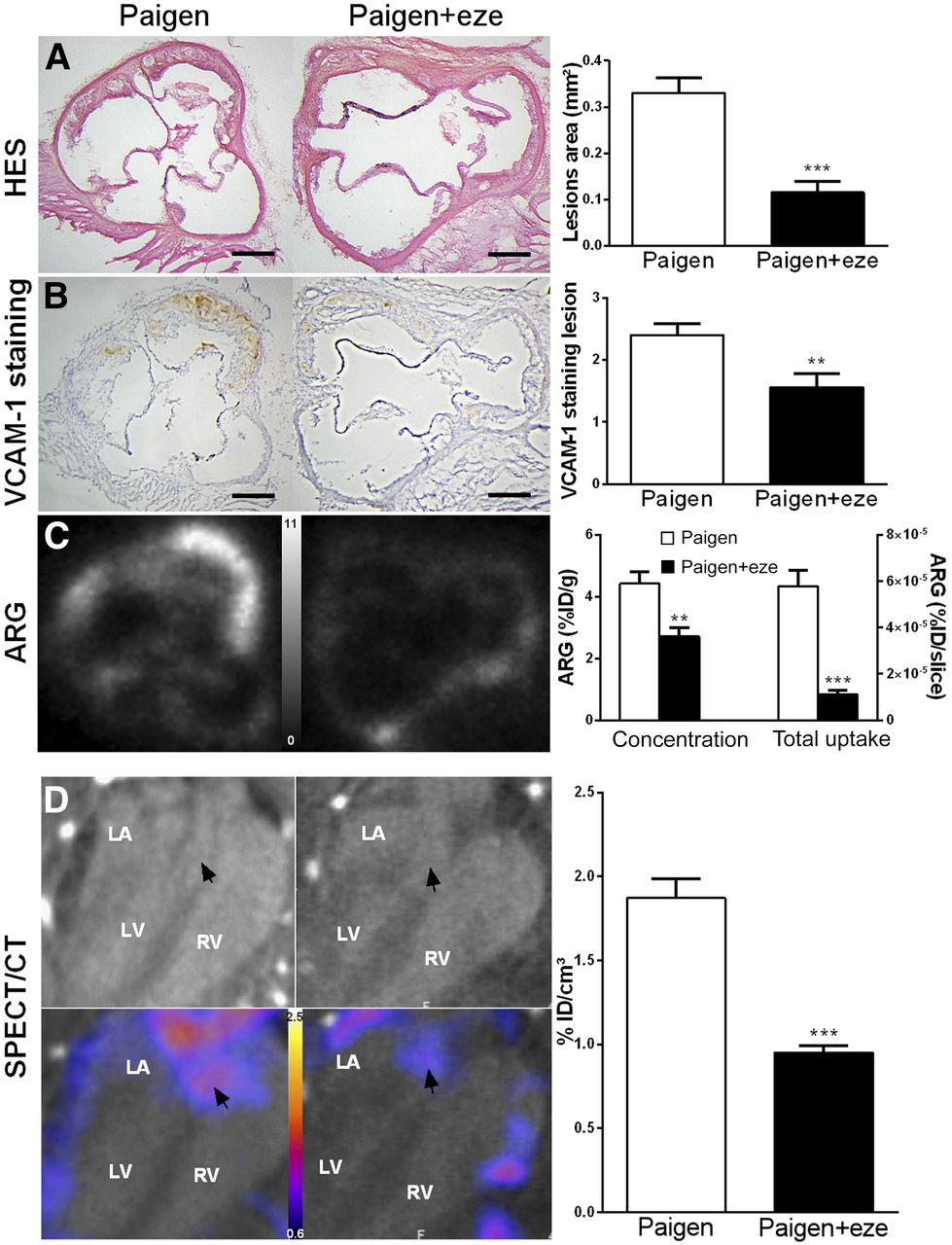
Effect of ezetimibe on plaque inflammation. (A and B) Paigen diet–induced aortic lesions and ezetimibe significantly reduced plaque volume and VCAM-1 expression as determined by histology (A) and immunohistochemistry (B). (C) As shown on autoradiographic images, 99mTc-cAbVCAM1–5 aortic uptake was significantly reduced in ezetimibe-treated mice when expressed either as concentration or as total uptake per slice. (D) In vivo quantification of 99mTc-cAbVCAM1–5 aortic uptake was in accordance with ex vivo data, and effect of ezetimibe was successfully monitored as shown on representative CT and SPECT/CT coronal views selected at level of aortic roots (arrows). **P < 0.01 vs. Paigen. ***P < 0.001 vs. Paigen. Scale bar = 200 μm; autoradiography scale = 0–11 %ID/g; SPECT scale = 0.8–2.6 %ID/cm3. ARG = autoradiography; HES = hematoxylin erythrosine saffron; LA = left atrium; LV = left ventricle; RV = right ventricle
Postmortem Analysis
The Paigen diet induced rapid atherosclerotic plaque formation in the aortic root of ApoE−/− mice (0.33 ± 0.03 mm2), which was significantly reduced by 73% by ezetimibe (0.11 ± 0.03 mm2, P < 0.0001). Histologically, as demonstrated by hematoxylin erythrosine saffron quantification, this was attributed to a significant decrease in core thickness and, therefore, intima-to-media ratio, whereas cap thickness remained unchanged (Fig. 2A and Supplemental Fig. 2). Moreover, as determined by VCAM-1 immunostaining, VCAM-1 score in atherosclerotic lesions was significantly reduced in ezetimibe-treated mice (1.56 ± 0.23 vs. 2.4 ± 0.18, P < 0.01) (Fig. 2B). Accordingly, on ex vivo 99mTc-cAbVCAM1–5 autoradiography, aortic uptake was significantly reduced in ezetimibe-treated mice (Fig. 2C). Indeed, 99mTc-cAbVCAM1–5 density was 39% lower in the ezetimibe-treated group than in the untreated group (2.7 ± 0.29 %ID/g vs. 4.4 ± 0.38 %ID/g, P < 0.01). Overall, the ezetimibe-induced decreases in VCAM-1 expression and burden of atherosclerotic lesions resulted in a profound, 81%, reduction in total 99mTc-cAbVCAM1–5 aortic uptake.
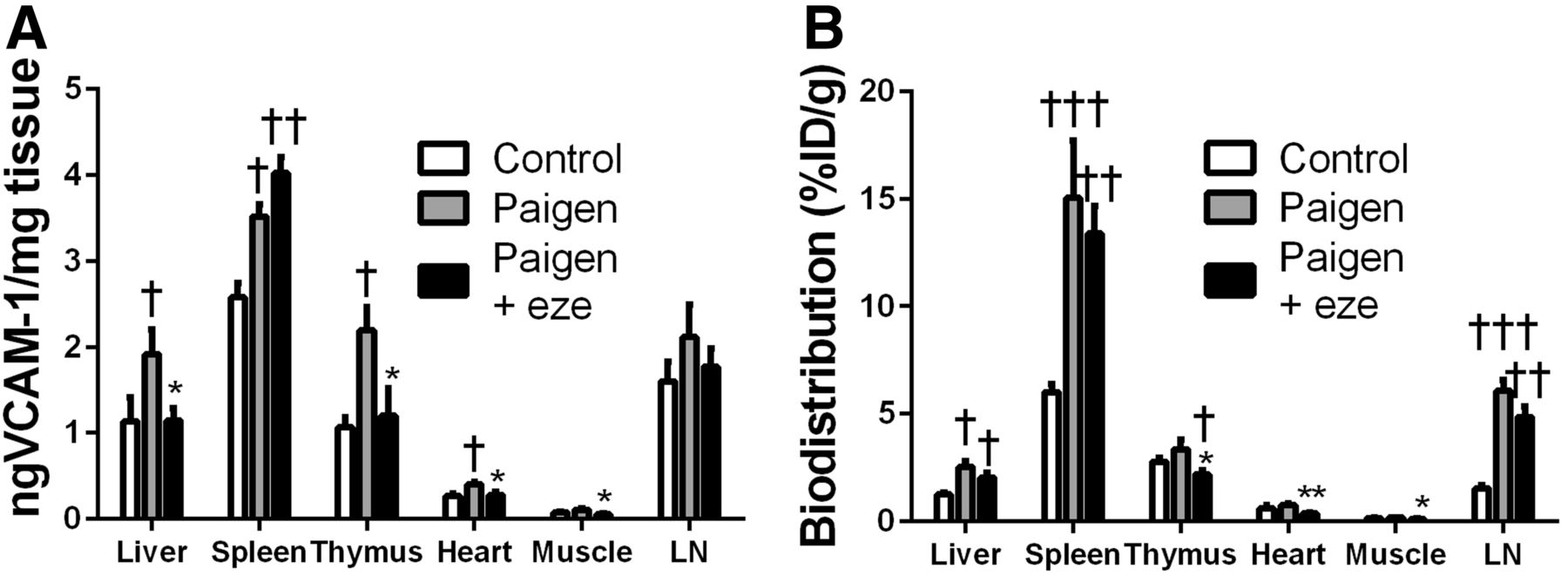
VCAM-1 protein expression was further investigated in various tissues by ELISA (Fig. 3). Low levels of VCAM-1 were found in the aortas of control mice, and no difference in the VCAM-1 expression of lesion-free abdominal aortas was found among the experimental groups. The Paigen diet induced a significant, 11-fold, increase in the ascending aorta and aortic arch of ApoE−/− mice. This effect was significantly inhibited by ezetimibe (Fig. 3A). Indeed, VCAM-1 expression in the ascending aorta was reduced by 47% in the ezetimibe-treated Paigen group in comparison to the untreated Paigen group (2.40 ± 0.60 ng/mg vs. 4.54 ± 0.37 ng/mg, P < 0.05) (Fig. 3A). 99mTc-cAbVCAM1–5 aortic uptake correlated with this VCAM-1 expression pattern. Indeed, 99mTc-cAbVCAM1–5 uptake in atherosclerotic lesions increased significantly in the untreated Paigen group, and ezetimibe treatment significantly reduced this effect by approximately 50% (3.97 ± 0.31 %ID/g vs. 1.98 ± 0.5 %ID/g, P < 0.01) (Fig. 3B). Consequently, 99mTc-cAbVCAM1–5 uptake strongly correlated with VCAM-1 expression in the aorta (R = 0.76, P < 0.0001) (Supplemental Fig. 3). Interestingly, the Paigen diet also induced an increase in VCAM-1 expression in all other investigated tissues, which reached statistical significance in the liver, spleen, thymus, and heart. This effect was significantly reduced by ezetimibe in the liver, thymus, heart, and muscle (Fig. 4A). As observed in the aorta, the biodistribution profile of 99mTc cAbVCAM1–5 correlated with that of VCAM-1 expression. Indeed, the Paigen diet induced an increase in 99mTc-cAbVCAM1–5 uptake in all investigated tissues, reaching statistical significance in the liver, spleen, and lymph nodes. This effect was significantly inhibited by ezetimibe in the thymus, heart, and muscle (Fig. 4B).
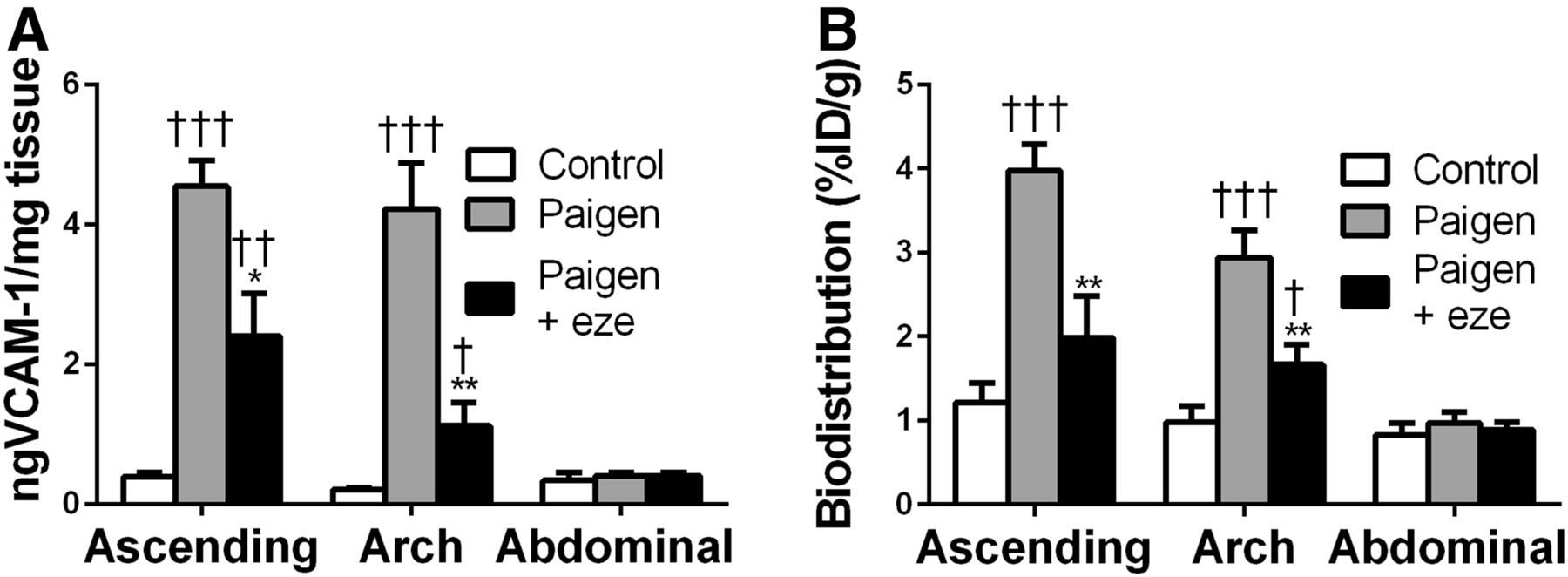
Antiinflammatory effect of ezetimibe on aortic atherosclerotic lesion. 99mTc-cAbVCAM1–5 uptake was reduced in Paigen+eze group and correlated well with VCAM-1 expression in aorta. (A) VCAM-1 expression was evaluated on ascending aorta, aortic arch, and abdominal aorta by ELISA and expressed as nanograms of VCAM-1 per milligram of tissue. (B) 99mTc-cAbVCAM1–5 uptake was determined by γ-well counting and expressed as %ID/g. Significant P levels: **<0.01 vs. Paigen, †<0.05 vs. control, ††<0.01 vs. control, and †††<0.001 vs. control.
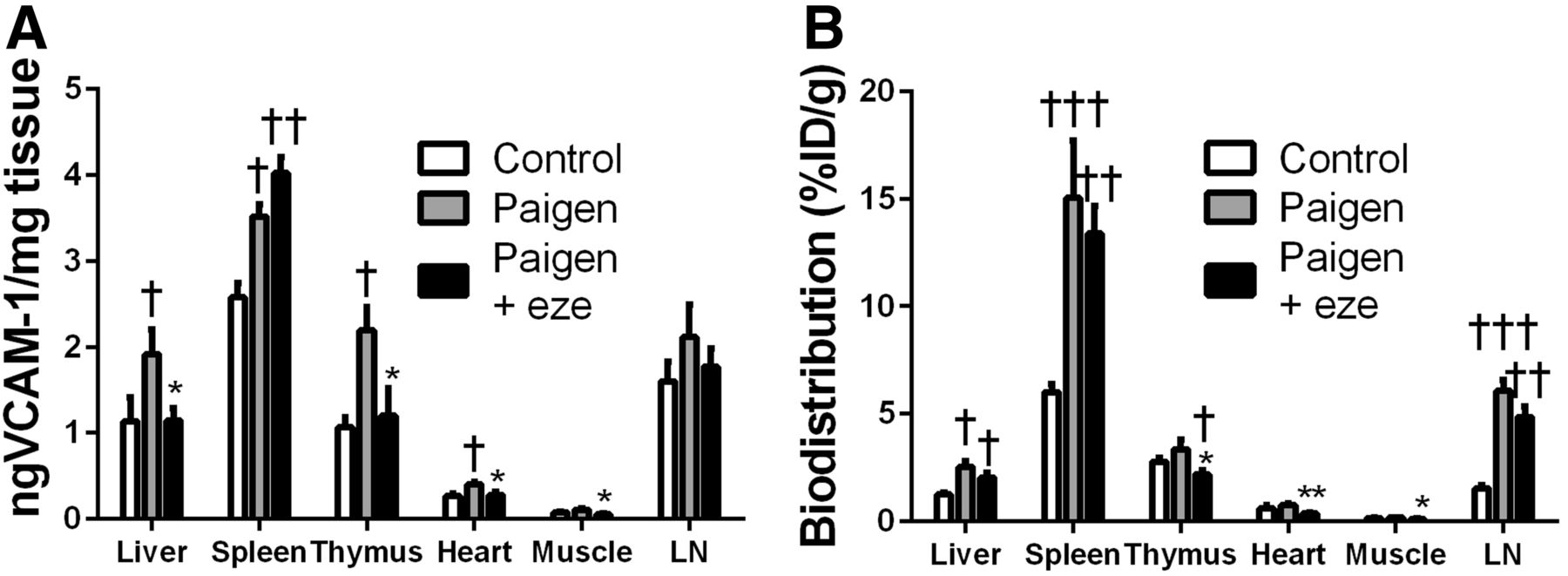
Effect of ezetimibe on VCAM-1 expression in liver, spleen, thymus, heart, muscle, and lymph node. (A) VCAM-1 expression was evaluated by ELISA and expressed as nanograms of VCAM-1 per milligram of tissue. (B) 99mTc-cAbVCAM1–5 uptake was determined by γ-well counting and expressed as %ID/g. Significant P levels: *<0.05 vs. Paigen, **<0.01 vs. Paigen, †<0.05 vs. control, ††<0.01 vs. control, and †††<0.001 vs. control. LN = lymph node.
As determined in vitro on human THP1 cells by Western blot analysis, VCAM-1 expression was observed on differentiated and stimulated macrophages but not on monocytes. Interestingly, ezetimibe reduced VCAM-1 expression by 18% (Supplemental Fig. 4).
DISCUSSION
Ezetimibe is a potent lipid-lowering drug that targets the Niemann-Pick C1-like 1 receptor in enterocytes, which in turn results in inhibition of cholesterol absorption by the intestine. Consequently, ezetimibe inhibits not only absorption of exogenous dietary cholesterol but also reabsorption of biliary cholesterol. Therefore, whereas statins act on endogenous cholesterol synthesis, ezetimibe lowers circulating cholesterol by inhibiting exogenous cholesterol absorption and endogenous cholesterol reabsorption.
To our knowledge, the present study was the first to evaluate the effect of ezetimibe in vivo on ApoE−/−mice fed the Paigen diet, a hyperlipidemic diet containing a combination of cholesterol and cholic acid. Cholic acid is one of the two major bile acids that potentiate cholesterol influx (18). Therefore, ApoE−/− mice fed the Paigen diet exhibit pronounced hypercholesterolemia resulting in the rapid development of atherosclerotic lesions within only 6 wk. Our finding of a significant increase in LDL cholesterol catabolism and LDL-derived cholesterol fecal excretion in ezetimibe-treated mice demonstrated that use of ezetimibe to inhibit cholesterol absorption by the intestine promotes elimination of proatherogenic LDL cholesterol, thereby potentially preventing it from forming atherosclerotic plaques. The benefits of the 50% reduction in Paigen diet–induced hypercholesterolemia were quantified in vivo using SPECT performed with 99mTc-cAbVCAM1–5. This radiolabeled single-domain antibody, which is currently undergoing clinical translation, targets the inflammatory marker VCAM-1 and has been validated as a sensitive and reproducible tool for imaging inflammation in mouse atherosclerotic lesions (14). In the present study, atherosclerotic lesions were readily observable in the aortic root and ascending aorta on SPECT images. As demonstrated by quantification based on contrast-enhanced CT images, ezetimibe induced a 2-fold reduction in 99mTc-cAbVCAM1–5 uptake. Such a result is in accordance with the profound and significant 73% decrease in plaque area observed on histologic sections of the aortic root. Meanwhile, it was previously reported that 99mTc-cAbVCAM1–5 SPECT quantification in mouse atherosclerotic lesions reflects not only plaque volume but also level of VCAM-1 expression within the lesion (14). An interesting finding of the present study was the reduced 99mTc-cAbVCAM1–5 uptake in the atherosclerotic lesions of ezetimibe-treated mice when tracer uptake was corrected for plaque volume as demonstrated by ex vivo autoradiography. This result suggests that, independently of the reduction in plaque size, ezetimibe induced a decrease in VCAM-1 expression as well. The reduction in VCAM-1 expression might be attributed either to a decrease in cholesterolemia and the associated lipotoxicity or to a direct antiinflammatory effect of the ezetimibe as suggested by recent studies. Indeed, Muñoz-Pacheco et al. showed that ezetimibe decreased the expression of adhesion molecules (ICAM-1, CD11a, and CD11b) on THP1 monocytes and thus inhibited their differentiation into macrophagelike cells and modified the expression of micro-RNA-155, which is involved in the development of atherosclerosis (19). Moreover, Cerda et al. demonstrated that ezetimibe reduced the expression of micro-RNA-221, a proatherogenic micro-RNA, on tumor necrosis factor α–stimulated endothelial cells of the human umbilical vein (20). These results are in accordance with those obtained by Western blot analysis of THP1 macrophages in the present study and may partly explain the reduction in VCAM-1 expression in the aortic sinus and aortic arch of ApoE−/− mice. Furthermore, the fact that ezetimibe acts on monocytes and macrophages may partly explain the reduction in VCAM-1 expression in lymphoid organs such as the spleen.
The ELISA results of the present study confirmed the 2- to 3-fold decrease in VCAM-1 protein density induced by ezetimibe in lesions in the ascending aorta and aortic arch. Moreover, the significant correlation found between the amount of VCAM-1 expressed by atherosclerotic lesions and the degree of 99mTc-cAbVCAM1–5 uptake as quantified ex vivo by γ-well counting confirmed the sensitivity of this imaging agent. Another interesting finding in our study was the significant increase in VCAM-1 expression induced by the Paigen diet in most other investigated organs—an effect that reached statistical significance in the liver, spleen, thymus, and heart and was inhibited by ezetimibe in most of them. This result is in accordance with previous studies demonstrating that the Paigen diet has not only a proatherosclerotic effect but a proinflammatory effect as well. Indeed, Vergnes et al. found that the Paigen diet induced hepatic inflammation and that this effect was attributable to the combination of cholesterol and cholic acid since removal of one of these two compounds significantly changed the pattern of proinflammatory gene expression (21). Our study further demonstrated that the Paigen diet induced inflammation in other organs and tissues, including the aorta, as shown by the level of VCAM-1 expression and that this effect was significantly reduced by ezetimibe. Moreover, the pattern of 99mTc-cAbVCAM1–5 biodistribution was in accordance with VCAM-1 expression in all investigated organs, suggesting that this imaging agent might be a suitable tool for noninvasive imaging of chronic inflammation in other relevant pathophysiologic settings.
Our study had some limitations. Endothelial cells and intraplaque cells expressing VCAM-1 contribute to 99mTc-cAbVCAM1–5 uptake within the lesion, and this relative contribution still needs to be determined. Image quantification and thereby inflammatory score depend on targeted cells, and an imaging agent targeting both endothelial and intraplaque cells will not provide the same information as one targeting just endothelial cells, such as anti-VCAM-1 microbubbles. Moreover, identifying the targeted cells would help us better understand the effect of a therapy on a specific cell type.
CONCLUSION
We demonstrated the antiatherogenic effect of ezetimibe on atherosclerotic plaque progression in an in vivo experimental model of ApoE−/− mice fed the Paigen diet. Contributing to this effect was increased LDL cholesterol catabolism and LDL-derived cholesterol fecal excretion, as well as reduced inflammation in atherosclerotic plaques as demonstrated using 99mTc-cAbVCAM1–5 imaging.
DISCLOSURE
This work was partly funded by grant ANR-11-INBS-0006 from France Life Imaging and by grant ANR-13-PRTS-0019 from the Agence Nationale pour la Recherche (ANR) and the Direction Générale de l’Offre de Soins (DGOS). Tony Lahoutte is a consultant for Camel-IDS NV and a member of the “Comité Stratégique” Institute for RadioElements (IRE). Nick Devoogdt is a consultant for Camel-IDS NV. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Dimitri Tissot, Laurie Arnaud, and Hadjer Guetarni—students from Université Grenoble Alpes—for their help with data collection, and we thank Stéphane Tanguy for assistance with ELISA.
Footnotes
↵* Contributed equally to this work.
Published online Mar. 9, 2017.
- © 2017 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication April 25, 2016.
- Accepted for publication December 26, 2016.




{kind=link}
{kind=link}
{kind=link}
{kind=link}