


Abstract
Trans-1-amino-3-18F-fluorocyclobutanecarboxylic acid (anti–18F-FACBC) is an amino acid PET tracer that has shown promise for visualizing prostate cancer. Therefore, we aimed to clarify the anti–18F-FACBC transport mechanism in prostate cancer cells. We also studied the fate of anti–18F-FACBC after it is transported into cells. Methods: For convenience, because of their longer half-lives, 14C compounds were used instead of 18F-labeled tracers. Trans-1-amino-3-fluoro-1-14C-cyclobutanecarboxylic acid (14C-FACBC) uptake was examined in human prostate cancer DU145 cells with the following substrates of amino acid transporters: α-(methylamino) isobutyric acid (a system A–specific substrate) and 2-amino-2-norbornanecarboxylic acid (a system L–specific substrate). The messenger RNA expression of amino acid transporters in human prostate cancer specimens was analyzed by complementary DNA microarray and quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). Gene expression in DU145 cells was analyzed by qRT-PCR. We also examined the knockdown effect of the amino acid transporters system ASC transporter 2 (ASCT2) and sodium-coupled neutral amino acid transporter 2 (SNAT2) on 14C-FACBC uptake. In addition, the possibility of 14C-FACBC incorporation into proteins was examined. Results: 14C-FACBC uptake by DU145 cells was markedly decreased to approximately 20% in the absence of Na+, compared with that in its presence, indicating that Na+-dependent transporters are mainly responsible for the uptake of this tracer. Moreover, 2-amino-2-norbornanecarboxylic acid inhibited the transport of 14C-FACBC to the basal level in Na+-free buffer. In contrast, α-(methylamino) isobutyric acid did not inhibit 14C-FACBC accumulation in DU145 cells. Human prostate tumor specimens and DU145 cells had similar messenger RNA expression patterns of amino acid transporter genes. Although SNAT2 and ASCT2 are 2 major amino acid transporters expressed in prostate tumor tissues and DU145 cells, ASCT2 knockdown using small interfering RNA was more effective in lowering 14C-FACBC transport than SNAT2. Almost all intracellular 14C-FACBC was recovered from the nonprotein fraction. Conclusion: ASCT2, which is a Na+-dependent amino acid transporter, and to a lesser extent Na+-independent transporters play a role in the uptake of 14C-FACBC by DU145 cells. Among the Na+-independent transporters, system L transporters are also involved in the transport of 14C-FACBC. Moreover, 14C-FACBC is not incorporated into proteins in cells. These findings suggest a possible mechanism of anti–18F-FACBC PET for prostate cancer.
Prostate cancer is one of the most common malignant neoplasms among men, and its incidence is increasing worldwide (1). In Japan, because the age-standardized prostate cancer incidence rate has been rapidly increasing since 1998 (2), there is an urgent demand for accurate prostate cancer diagnosis leading to an appropriate treatment strategy. However, the current methodology does not allow precise identification of the site of the disease within the prostate.
18F-FDG is most commonly used for whole-body PET but does not work well for certain tumor tissues, such as brain tumors and tumors located in the pelvic region, because of its physiologic accumulation and elimination (3). In addition to 18F-FDG, radiolabeled amino acids have been studied as potential tumor-seeking PET agents for clinical use (4). The most frequently used amino acid PET tracer is 11C-methionine because of its easy and fast radiosynthesis (5). The availability and clinical efficacy of 11C-methionine in patients with brain tumors (4) and prostate cancers (6) have been investigated. Besides PET with radiolabeled natural amino acids, PET with synthetic amino acids labeled with 11C, such as 1-11C-aminocyclobutanecarboxylic acid (11C-ACBC) (7), 1-11C-aminocyclopentanecarboxylic acid (7), and α-11C-aminoisobutyric acid (8), exhibit high tumor–to–nontumor concentration ratios. Among these studies, Washburn et al. (7) show that 11C-ACBC is the most selective radioprobe for tumor imaging in an animal model. In a clinical study, 11C-ACBC is shown to be a good diagnostic probe specific for astrocytoma, suggesting that it can be taken up by such tumors, whereas it is negligibly taken up by normal brain tissue (9). However, because of the short half-life of 11C, the availability of 11C-labeled tracers is limited to facilities equipped with an in-house cyclotron, thus preventing the widespread use of these compounds. Therefore, 18F-labeled amino acids are more attractive as PET agents and are widely used in many facilities.
To improve the availability of 11C-ACBC, Shoup et al. (10) developed trans-1-amino-3-18F-fluorocyclobutanecarboxylic acid (anti–18F-FACBC). Anti–18F-FACBC is a synthetic l-leucine analog that exhibits high tumor-specific accumulation in patients with glioblastoma multiforme (10), renal papillary cell cancer (11), and prostate cancer (12). One of the interesting features of anti–18F-FACBC is that its renal excretion is much slower than that of 18F-FDG (13); this slower excretion could be helpful for prostate cancer imaging.
Little is known about the transport mechanism of anti–18F-FACBC into cells, although anti–18F-FACBC is thought to cross the plasma membrane via amino acid transporters because of its structural similarity to natural amino acids. Because of the identification and kinetic studies of amino acid transporters, these transporters are categorized into at least 17 distinct classes (14). Neutral amino acids are considered to be mainly transported by 3 systems: A, ASC, and L (15). Systems A and ASC mainly serve to take up amino acids with short, polar, or linear side chains such as l-alanine and l-serine. In contrast, large, branched, and aromatic amino acids such as l-tyrosine mainly enter cells via system L (16). Although several transporters may be involved in anti–18F-FACBC uptake, previous studies show that the uptake of anti–18F-FACBC is mediated by the Na+-independent system L amino acid transporter in rat 9L gliosarcoma cells (17,18). However, the mechanism of anti–18F-FACBC uptake by prostate cancer cells is not well understood. Furthermore, the gene expression of amino acid transporters in prostate cancer cells has not been fully clarified.
In the present study, we characterized anti–18F-FACBC transport in human prostate cancer cells using a cell uptake assay to find candidate transporters. The gene expression profile of amino acid transporters was examined by complementary DNA (cDNA) microarray in human prostate biopsy specimens. Furthermore, we confirmed the gene expression of amino acid transporters in the human prostate cancer tissues and DU145 cells by quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). In addition, we examined the role of Na+-dependent amino acid transporters in anti–18F-FACBC uptake, using the small interfering RNA (siRNA) transfection method, and the intracellular fate of anti–18F-FACBC after its uptake.
MATERIALS AND METHODS
Materials
For convenience, because of their longer half-lives (5,700 y), 14C-labeled amino acids and analogs were used rather than 18F (half-life, 110 min) or 11C (half-life, 20 min). Trans-1-amino-3-fluoro-1-14C-cyclobutanecarboxylic acid (14C-FACBC; 2.08 GBq/mmol) was radiosynthesized by EaglePicher Pharmaceutical Services. l-1-14C-methionine (14C-Met, 2.04 GBq/mmol) was purchased from American Radiolabeled Chemicals. Both l-14C (U)-alanine (14C-Ala; 5.92 or 6.29 GBq/mmol) and l-14C (U)-leucine (14C-Leu; 11.99 GBq/mmol) were purchased from both Moravek Biochemicals and PerkinElmer.
The DU145 human prostate cancer cell line was obtained from the American Type Culture Collection.
Collection of Human Biopsy Specimens
Human prostate biopsy specimens (Table 1) for cDNA microarray and qRT-PCR were obtained by needle biopsy from 6 patients with elevated prostate-specific antigen levels at Kanazawa University Hospital from November 2007 to February 2008. All patients involved in this study were informed of a complete guarantee of confidentiality of their individual records, our ethically acceptable experiment, and the right to refuse or withdraw. The biopsy specimens used in this study were obtained from these patients with their informed consent. The specimens used for pathologic diagnosis were reviewed by pathologists; the specimens for gene expression analysis were immediately stored in RNAlater (Ambion) at 4°C and subsequently stored at −80°C until required. Patients were designated prostate cancer–free if cancer cells were not detected in their biopsy specimens.
List of Human Prostate Biopsy Specimens
Culture of Human Cell Lines
DU145 cells were cultured in Dulbecco's modified Eagle's medium (Life Technologies) supplemented with 10% fetal bovine serum (American Type Culture Collection), 100 units of penicillin per milliliter, and 100 μg of streptomycin per milliliter (Life Technologies). The cells were cultured at 37°C in 5% CO2/95% air; these cells were used for the messenger RNA (mRNA) quantitation and uptake assay.
RNA Extraction
Total RNA was extracted from human prostate biopsy specimens and DU145 cells using the RNeasy Micro and RNeasy Plus Mini Kits (Qiagen), respectively.
cDNA Microarray Analysis
All reagents, equipment, and software were purchased from Agilent Technologies. cDNA microarray analysis was performed according to the manufacturer's protocol. In brief, total RNA was subjected to linear amplification and Cy3 labeling using the Low RNA Input Linear Amplification Kit and a One Color RNA Spike-In Kit and was subsequently hybridized to a 44-K Whole Human Genome Microarray using a Gene Expression Hybridization Kit. Data were extracted using Feature Extraction software (version 9.5.3) and were analyzed using GeneSpring GX software (version 11.0).
qRT-PCR
cDNA was synthesized from 250 ng of total RNA of human biopsy samples using the WT-Ovation RNA Amplification System (NuGEN Technologies) according to the manufacturer's protocol. Total RNA (1 μg) from DU145 cells was reverse-transcribed using a Transcriptor First Strand cDNA Synthesis Kit (Roche Applied Science) by random hexamer priming at 50°C for 1 h according to the manufacturer's instructions. The primers (Supplemental Table 1; supplemental materials are available online only at http://jnm.snmjournals.org) were designed using the Universal ProbeLibrary Assay Design Center (Roche Applied Science) and were synthesized by Nihon Gene Research Laboratories. qt-PCR was performed using an Mx3005P or Mx3000P QPCR system (Agilent Technologies), FastStart Universal Probe Master (ROX), and the Universal ProbeLibrary (Roche Applied Science) with the following profile: 1 cycle of enzyme activation at 95°C for 10 min, 40 cycles of denaturing at 95°C for 15 s, and extension at 60°C for 1 min. All reactions were run in triplicate. The PCR products were analyzed by agarose gel electrophoresis, and no nonspecific PCR bands were detected. The PCR products for each gene were subsequently purified using a High Pure PCR Cleanup Micro Kit (Roche Applied Science) and quantified from the concentration and base pair numbers of amplicons. The mRNA copy number was calculated from standard curves generated by amplifying serial dilutions of a known quantity of purified amplicons. Expression data were normalized against the copy number of 18S ribosomal RNA.
The correlation coefficient of the log ratio between the results of the cDNA microarray and qRT-PCR was calculated as follows: log2(mRNA expression in cancer specimens/mRNA expression in normal specimens).
Measurement of 14C-Labeled Amino Acid Transport
Transport assays were based on the methods described by Shikano et al. (19,20). In brief, DU145 cells were seeded on 24-well tissue culture plates at 1 × 105 cells per well with 1 mL of culture medium. Transport assays were then conducted on the next day after inoculation (semiconfluent phase cells).
The sodium-containing incubation medium used was based on phosphate-buffered saline (pH 7.4–7.6), consisting of 137 mM NaCl, 2.7 mM KCl, 8 mM Na2HPO4, 1.5 mM KH2PO4, 5.6 mM d-glucose, 0.9 mM CaCl2, and 0.5 mM MgCl2. In the sodium-free incubation medium (Na+-free buffer), NaCl and Na2HPO4 were replaced with the equivalent concentrations of choline chloride and K2HPO4, respectively. After the culture medium was removed, each well was incubated with 1 mL of incubation medium for 10 min at 37°C. The cells were then incubated with 0.3 mL of incubation medium containing the respective 14C-labeled amino acid (10 μM) for 10 min at 37°C. At the end of uptake, each well was rapidly washed twice with 1 mL of ice-cold incubation medium. The cells were then solubilized in 0.5 mL of 0.1N NaOH, and the radioactivity of each aliquot was measured using a liquid scintillation counter (LS 6000SC; Beckman Coulter) after scintillation cocktail (Ultima Gold; PerkinElmer) was added. The protein content of the cell lysate was determined by assay with a BCA Protein Assay kit (Thermo Fisher Scientific).
For the competitive inhibition assay, the following compounds were used at a final concentration of 1 mM: α-(methylamino) isobutyric acid (MeAIB—a substrate specific to system A transporters), 2-amino-2-norbornanecarboxylic acid (BCH—a substrate specific to system L transporters), p-aminohippurate (PAH—a substrate specific to organic anion transporters), and tetraethylammonium chloride (TEA—a substrate specific to organic cation transporters). Basal transport activity was defined by the results of these assays performed on ice.
All experimental conditions were examined in triplicate. Reproducibility was confirmed by repeating the same experiment.
siRNA Transfection
All siRNAs used in this study were purchased from Ambion. We used 2 different kinds of siRNAs for each target gene; each transfection was performed using only one of them. The identification numbers of siRNAs used are as follows: s12916 and s12918 for system ASC transporter 2 (ASCT2) and s633 and s634 for sodium-coupled neutral amino acid transporter 2 (SNAT2). Silencer Select Negative Control 1 and 2 siRNA (catalog nos. 4390843 and 4390846) were used as controls. siRNA transfection was performed according to the manufacturer's instructions. DU145 cells were transfected with 5 nM of each siRNA in antibiotic-free growth medium using Lipofectamine 2000 (Life Technologies). After transfection (48 h), the cells were used for amino acid uptake experiments. The efficiency and specificity of each siRNA were confirmed by qRT-PCR. We rated the result as the target gene–specific effect when both siRNAs for the same target gene significantly altered the transport of 14C-labeled amino acids.
Measurement of 14C-Labeled Amino Acid Incorporation into Proteins
DU145 cells were incubated with 50 μM 14C-FACBC at 37°C for 3 or 24 h. The cells were then washed twice with phosphate-buffered saline and lysed in CelLytic M cell lysis reagent (Sigma-Aldrich) containing protease inhibitor cocktail (Sigma-Aldrich). Cell lysates were centrifuged at 18,000g for 15 min at 4°C, and the supernatants were subjected to trichloroacetic acid (TCA) precipitation. The precipitation reaction was performed for 10 min on ice by adding ice-cold TCA (final concentration, 10%) to the supernatants, and the TCA precipitates were washed 3 times with ice-cold 5% TCA. The TCA precipitates and cell debris were lysed in 0.1N NaOH and neutralized with 0.1N HCl. The radioactivity of the TCA precipitates, TCA-soluble fraction, and cell debris was measured using a liquid scintillation counter after scintillation cocktail was added.
Statistical Analysis
Data are presented as means and SDs. P values were calculated using a 2-tailed paired Student t test for comparison between 2 groups. A P value less than 0.05 was considered significant.
RESULTS
In Vitro Uptake and Competitive Inhibition Assay
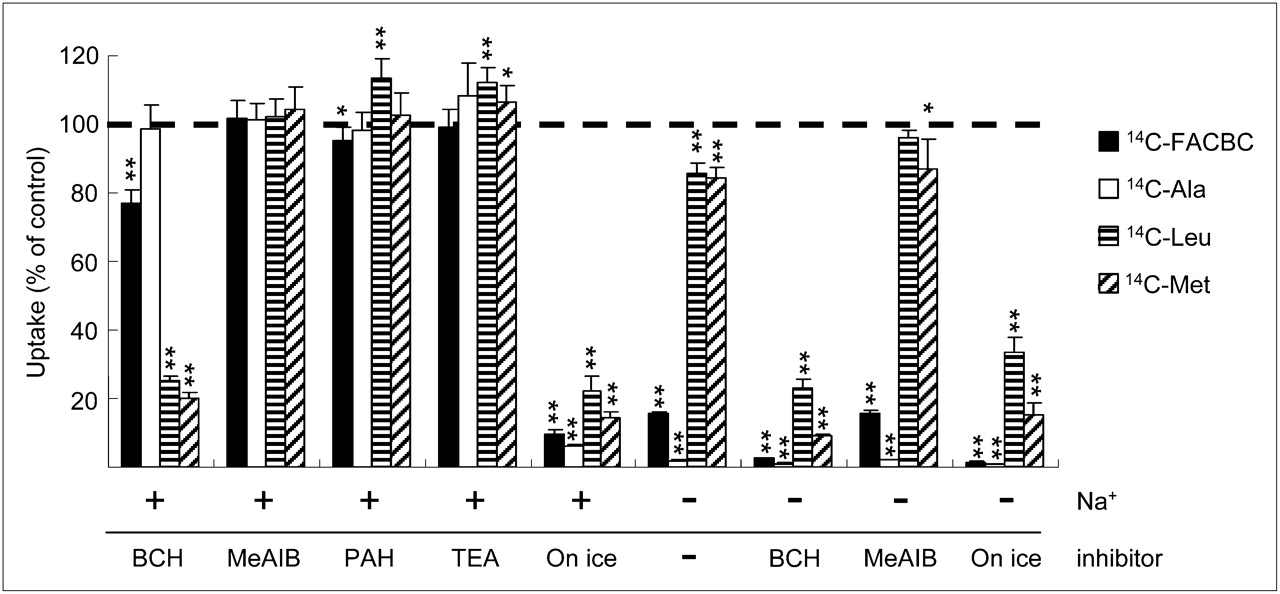
We examined the contribution of individual transport systems to anti-FACBC uptake by measuring the intracellular accumulation of 14C-FACBC in DU145 cells in the absence or presence of Na+ or various types of inhibitors (Fig. 1). In Na+-free buffer, 14C-FACBC uptake markedly decreased to approximately 20% of the control. BCH, a substrate specific to Na+-dependent systems B0 and B0,+ and Na+-independent system L, decreased the transport of 14C-FACBC to approximately 75% in Na+-containing buffer and inhibited its uptake to the basal level in Na+-free buffer. In contrast, 14C-FACBC uptake was unchanged in the presence of MeAIB (a substrate specific to Na+-dependent system A and Na+-independent H+-coupled amino acid transport system [system PAT]) and TEA. PAH slightly decreased 14C-FACBC uptake to approximately 95% of the control.

Comparison of 14C-FACBC, 14C-Ala, 14C-Leu, and 14C-Met uptake by DU145 cells. Data are expressed as percentage of uptake in cells that were incubated without inhibitors. Uninhibited uptake of 14C-FACBC, 14C-Ala, 14C-Leu, and 14C-Met was 6,510.71, 3,086.29, 3,193.04, and 4,036.06 pmol/mg of protein, respectively. Each bar represents mean and SD (n = 3–6). *P < 0.05. **P < 0.01.
Regarding the comparison of uptake mechanisms, other 14C-labeled natural amino acids (i.e., Ala, Leu, and Met) were also examined (Fig. 1). 14C-FACBC uptake in Na+-containing buffer without inhibitors was higher than the uptake of other 14C-labeled natural amino acids (Fig. 1), and we found a difference in the Na+ dependency of the uptake of these amino acids by DU145 cells. That is, the Na+ dependency of 14C-FACBC and 14C-Ala uptake was relatively larger than that of 14C-Leu and 14C-Met. 14C-Ala transport was completely abolished in Na+-free buffer, and BCH, MeAIB, PAH, and TEA did not inhibit its uptake in Na+-containing buffer.
By contrast, both 14C-Leu transport and 14C-Met transport were decreased to approximately 85% in Na+-free buffer, and MeAIB, PAH, and TEA did not inhibit their accumulation in Na+-containing buffer. Furthermore, BCH inhibited the uptake of both 14C-Leu and 14C-Met to the basal level in both Na+-containing and Na+-free buffer, because for 14C-Leu and 14C-Met, results for assays performed on ice were almost the same as those for assays performed in the presence of BCH.
Amino Acid Transporter mRNA Expression
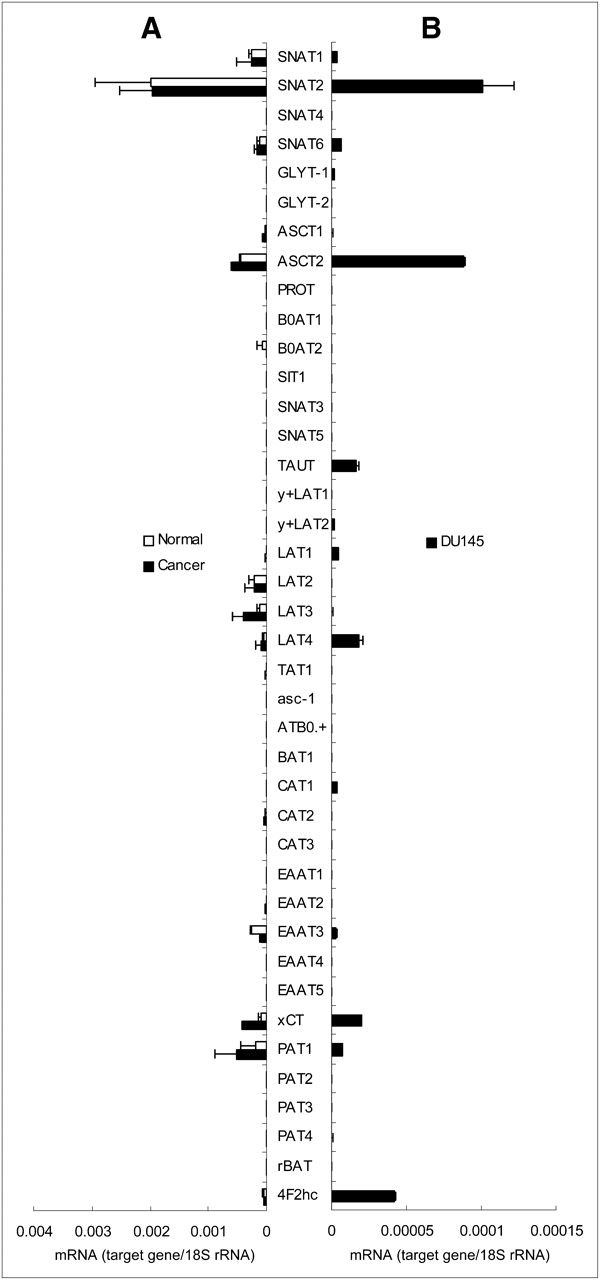
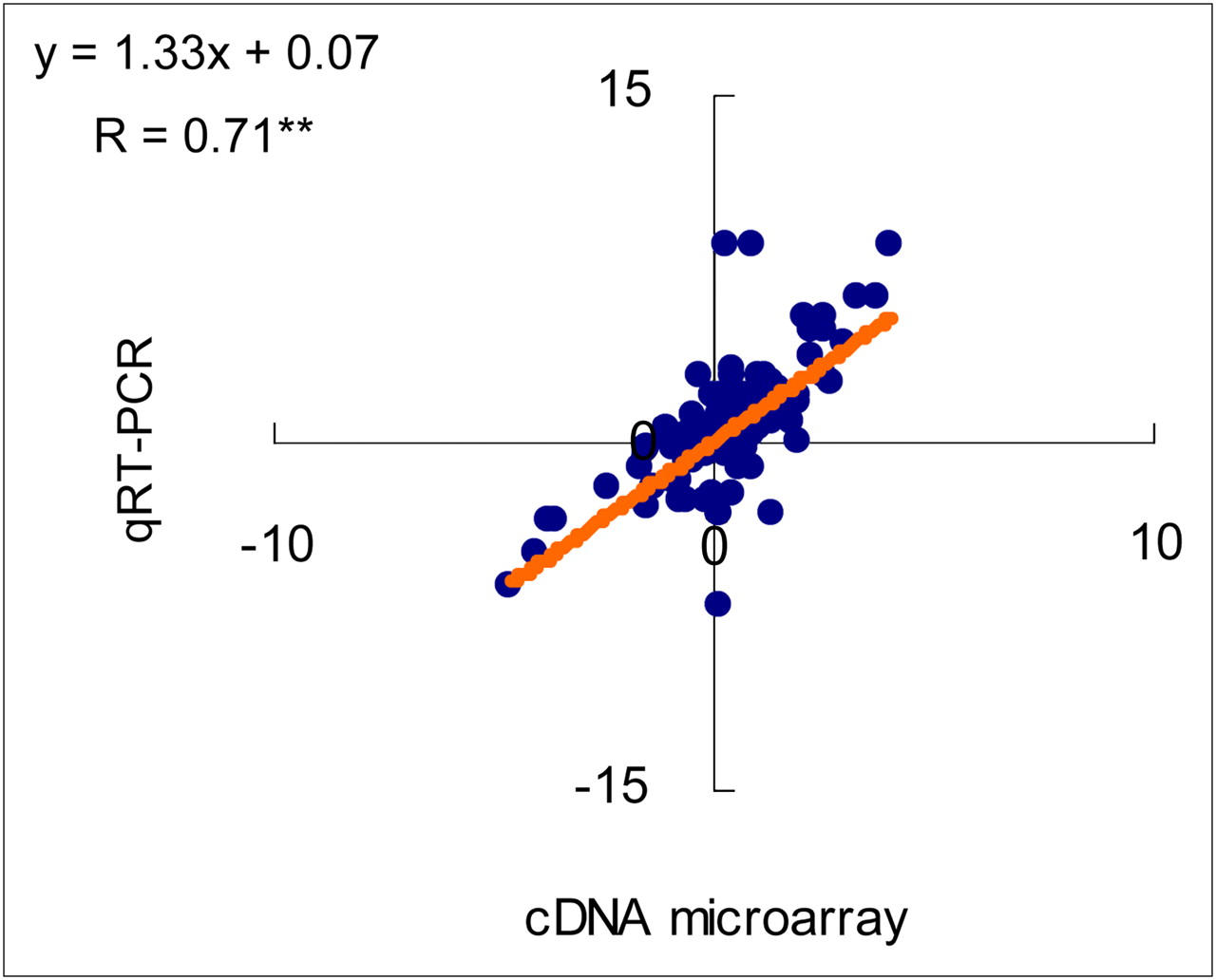
To evaluate transporter gene expression in biopsy specimens from patients with prostate tumor tissues, gene expression was profiled by cDNA microarray (data not shown). On the basis of the cDNA microarray results, 40 gene expressions of amino acid transporter mRNAs were quantified by qRT-PCR in the clinical specimens and DU145 cells (Fig. 2). There was a strong significant correlation between the log ratio determined by the cDNA microarray and qRT-PCR for the gene expression of amino acid transporters analyzed in the tissue specimens (r = 0.71, P < 0.01) (Fig. 3). qRT-PCR revealed that the mRNA expression of both ASCT2 and SNAT2, which are Na+-dependent transporters, was remarkably high in both the human prostate biopsy specimens and DU145 cells (Fig. 2). In addition, several amino acid transporter mRNAs (i.e., ASCT2, LAT3, xCT, and PAT1) tended to be expressed more in human prostate cancer samples than in noncancerous samples (Fig. 2). The mRNA expression pattern of amino acid transporters in DU145 cells was similar to that in biopsy specimens.

mRNA expression levels of amino acid transporters in human prostate biopsy specimens (A) and DU145 cells (B) determined by qRT-PCR. Each bar represents mean and SD (n = 3). rRNA = ribosomal RNA.

Correlation between log ratio of amino acid transporters in human prostate biopsy specimens determined by cDNA microarray and qRT-PCR. **P < 0.01.
ASCT2 Involvement in 14C-FACBC Uptake
To evaluate the involvement of Na+-dependent amino acid transporters in anti-FACBC uptake by prostate cancer cells, we examined the transport of 14C-FACBC in ASCT2 and SNAT2 knockdown DU145 cells. The expression of ASCT2 and SNAT2 were silenced with a gene-specific siRNA in DU145 cells. qRT-PCR showed that ASCT2 and SNAT2 mRNA expression significantly decreased in the ASCT2 and SNAT2 siRNA-transfected cells, compared with those of the controls (Fig. 4A). 14C-FACBC uptake was decreased to 69% and 58% of the control in DU145 cells with ASCT2 knocked down by s12916 and s12918 siRNAs, respectively; uptake decreased to 83% of the control in DU145 cells with SNAT2 knocked down by both s633 and s634 (Fig. 4B).

Effects of ASCT2 or SNAT2 knockdown, compared with control siRNA-transfected cells. (A) mRNA expression levels of amino acid transporters in siRNA-treated DU145 cells. (B) Uptake of 14C-FACBC, 14C-Ala, 14C-Leu, and 14C-Met by each siRNA-transfected DU145 cell. Each bar represents mean and SD (n = 6). **P < 0.01.
For comparison, 14C-labeled Ala, Leu, and Met were examined in the same way (Fig. 4B). The alteration in 14C-Ala uptake was similar to that of 14C-FACBC; 14C-Ala uptake decreased to 59% and 54% in DU145 cells with ASCT2 knocked down by s12916 and s12918 siRNAs, respectively, and to 79% and 84% in cells with SNAT2 knocked down by s633 and s634, respectively. 14C-Leu uptake was slightly reduced to 93% in ASCT2 knockdown cells and decreased to 93% (s633) and 86% (s634) of the control in SNAT2 knockdown cells. 14C-Met uptake did not change significantly in either ASCT2 or SNAT2 knockdown cells, compared with the controls.
Intracellular Fate of 14C-FACBC
Because little is known about the intracellular fate of 14C-FACBC, we measured the radioactivity of TCA precipitate (protein fraction), TCA-soluble fraction (intracellular suspended fraction), and cell debris (nucleic acid fraction), which were prepared from DU145 cells incubated with 14C-FACBC or 14C-Met. Figure 5A shows that almost all intracellular 14C-FACBC was detected in the TCA-soluble fraction after a 24-h incubation. In contrast, 67% and 81% of intracellular 14C-Met was detected in the TCA precipitate after 3- and 24-h incubations, respectively (Fig. 5B). To further clarify whether 14C-FACBC is incorporated into proteins, we analyzed the cell lysates obtained from DU145 cells incubated for 24 h with 14C-FACBC or 14C-Met by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After SDS-PAGE, the distribution of radioactivity of the radiolabeled proteins was analyzed. However, no radiolabeled bands corresponding to cellular proteins were detected on the SDS-PAGE gel with lysates from the cells incubated with 14C-FACBC, whereas significant radiolabeled protein bands were evident with the lysates from the cells incubated with 14C-Met (data not shown).

Intracellular distribution of 14C-FACBC (A) and 14C-Met (B) when DU145 cells were incubated with each tracer for 3 or 24 h. Each bar represents mean and SD (n = 3), except 14C-FACBC for 3 h, which represents mean (n = 2).
DISCUSSION
This is the first report, to our knowledge, that demonstrates the transport mechanism of anti-FACBC that refers to amino acid transporter subtype. Our competitive inhibition assays clearly demonstrate a major role of Na+-dependent amino acid transporters for 14C-FACBC uptake by DU145 cells. The mRNA expression analysis results show that ASCT2 and SNAT2 are 2 major amino acid transporters in prostate tumor tissues and DU145 cells. The RNA interference studies revealed that ASCT2 knockdown in DU145 cells leads to a corresponding reduction of 14C-FACBC accumulation. Given the possibility of compensatory uptake, the effect of ASCT2 knockdown on 14C-FACBC uptake is thought to be more pronounced. Thus, ASCT2 is critically involved in 14C-FACBC uptake. In contrast, organic anion and cation transport systems involved in the uptake of many kinds of drugs do not apparently take part in 14C-FACBC uptake by DU145 cells, also showing that the amino acid transporters are mainly involved in the uptake of anti-FACBC.
While considering the transport mechanism of anti-FACBC, we concluded that anti-FACBC is transported into prostate cancer cells in a fashion similar to the transport of l-Ala. Among the amino acids examined in this study, the characteristics of 14C-FACBC uptake were similar to those of 14C-Ala but not those of 14C-Met and 14C-Leu. l-Ala is a typical substrate of ASCT2 (14), which is a major transporter involved in 14C-FACBC uptake by DU145 cells. Therefore, it is thought that anti-FACBC shares the binding sites of ASCT2 with other substrates.
SNAT2 is one of the isoforms of system A amino acid transporters, and its typical substrate is l-Ala. It is reported that the expression of this transporter is more widespread than those of other system A subtypes (21). However, the contribution of SNAT2 to anti-FACBC uptake by prostate cancer cells would be lower than that of ASCT2, although its gene was the most expressed among amino acid transporters in prostate cancer tissues and DU145 cells. 14C-FACBC uptake slightly decreased in SNAT2 knockdown cells. However, MeAIB did not affect 14C-FACBC uptake in the inhibition assay. Because 14C-Ala uptake was also affected by the knockdown of SNAT2 but not MeAIB, the inhibitory effect of the knockdown on SNAT2 transport function might be stronger than that of MeAIB. In addition, it is known that the affinity of substrate amino acids—particularly small neutral amino acids—to SNAT2 is lower than that to ASCT2 (14). Their affinity (Km value) to ASCT2 is less than 20 μM (22), whereas the affinity of typical substrates, such as L-Ala and MeAIB, to SNAT2 is 200–400 μM (23,24). Hence, the affinity of 14C-FACBC to SNAT2 may be lower than that to ASCT2; this difference in the Km values of anti-FACBC would reflect the contribution to its transport. However, the extent of the affinity between 14C-FACBC and transporters is still a matter of speculation at this time; we are currently planning to confirm this conjecture.
System L transporters are also possibly involved in the Na+-independent transport of 14C-FACBC in DU145 cells, although our data suggest that Na+-dependent transporters play a major role in 14C-FACBC uptake by DU145 cells. The LAT1 subtype of system L functions as a transporter for neutral amino acids with large, branched, or aromatic side chains (e.g., leucine and phenylalanine) when LAT1 heterodimerizes with the 4F2 heavy chain (25). It is reported that the expression of LAT1 is increased in many types of cancer tissues (25) and that LAT1 transports substrate amino acids with a high affinity, with Km values of 15–50 μM (25). Hence, although the mRNA expression of LAT1 was lower than that of ASCT2 in human biopsy samples and DU145 cells, LAT1 may be also involved in anti-FACBC uptake.
Among the Na+-dependent transporters, the system B0, + transporter (ATB0, +) may be involved in the transport of anti-FACBC. The involvement of other Na+-dependent transporters in anti-FACBC uptake is also suggested because 14C-FACBC uptake was inhibited to approximately 80% in Na+-free buffer, and the transport activity of 14C-FACBC remained at more than 50% of the control in ASCT2 knockdown cells. The inhibition assay using BCH implies the participation of systems B0 and B0, +. Because the affinity of substrates to system B0, + is higher than that to system B0 (26–28), the contribution of system B0, + to anti-FACBC uptake would be higher than that of system B0.
The anti-FACBC transport mechanism clarified in this study has important implications for the feasibility of anti–18F-FACBC PET. This mechanism illustrates the clinical application of anti–18F-FACBC for imaging cancers in a wide variety of sites, because the expression of ASCT2 and LAT1 are elevated in many kinds of primary human cancer tissues relative to other neutral amino acid transporters (25). These amino acid transporters have important roles in anti–18F-FACBC transport. Furthermore, it has been reported that ASCT2 expression is associated with aggressive biologic behavior in colorectal adenocarcinomas (29). Witte et al. (29) showed that the survival of colorectal adenocarcinoma patients decreases with an increased percentage of ASCT2-positive cancer cells. These findings imply that anti–18F-FACBC PET is effective for the diagnosis of other malignancies in addition to prostate cancer and that anti–18F-FACBC PET might be able to predict patient prognosis. Further investigations on the expression profiling of amino acid transporters in cancers and the relationship between prognosis of cancer patients and expression level of amino acid transporters would clarify the target sites that can be effectively diagnosed using anti–18F-FACBC in addition to the potential ability of this radiotracer.
Considering how anti-FACBC is used or metabolized once it is taken up by cells, how it is incorporated into proteins because of its structural similarity to natural amino acids should be determined. However, this study indicates that 14C-FACBC is not incorporated into proteins. The intracellular distribution pattern of 14C-FACBC was quite different from that of 14C-Met. The intracellular distribution ratio of TCA precipitate containing 14C-FACBC was 2.08%, even when the cells were incubated with 14C-FACBC for 24 h. The radioactivity of TCA precipitate including 14C-FACBC was not considered significant because no protein bands were observed in the lysates from the cells incubated with 14C-FACBC in SDS-PAGE. The radioactivity of TCA precipitate might be derived from the nonspecific binding of 14C-FACBC to cellular proteins. A previous report shows that 1-11C-aminocyclopentanecarboxylic acid, which is a structural analog of anti-FACBC, is not metabolized and remains intact inside cells (30). Therefore, anti-FACBC may remain intact without being incorporated into cellular proteins—an important feature of PET tracers for clinical use, because the incorporation of anti-FACBC into enzymes, peptides, and proteins that are essential for biologic phenomena might cause significant adverse effects. However, this study suggests that almost all 14C-FACBC in cells was recovered from the nonprotein fraction. Furthermore, it is reported that “cold” anti-FACBC does not exhibit any acute toxic effects on Sprague–Dawley rats, even when the amount of injected “cold” anti-FACBC was approximately 1 × 106 times in excess of the amount that a patient would receive (10). Thus, these findings indicate that this radiotracer would not affect the biologic function of proteins; these features would enable the safe use of anti–18F-FACBC in PET examination. In addition, these features indicate that the images derived from anti–18F-FACBC PET would simply represent the amount of amino acid transporters involved in anti-FACBC uptake and their activity and not protein synthesis or amino acid metabolism.
Our findings are currently limited to 2 amino acid transporter systems extracted from an inhibition assay, mRNA expression analysis, and RNA interference study. To further interpret the transportation mechanism of anti-FACBC, future studies are necessary to clarify the affinity of anti-FACBC for the amino acid transporters that are expressed, to some extent, in cancer cells and the expression of amino acid transporter proteins in malignancies.
CONCLUSION
We demonstrated that Na+-dependent amino acid transporters predominantly function in 14C-FACBC uptake by DU145 cells. Na+-independent system L transporters are also involved in 14C-FACBC transport. Gene expression analysis suggests that the mRNA expression of both ASCT2 and SNAT2 is remarkably high in human prostate specimens and DU145 cells. ASCT2, which is a Na+-dependent amino acid transporter, at least in part plays an important role in the uptake of 14C-FACBC by DU145 cells. Moreover, 14C-FACBC is not incorporated into cellular protein. These findings suggest a possible mechanism of anti–18F-FACBC PET for prostate cancer.
DISCLOSURE STATEMENT
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
We thank Hiroyo Araki, Kodai Nishi, Masato Ogura, and Fumiya Takahashi for their assistance with the gene expression analysis. We also acknowledge the valuable discussion with Dr. Shuntaro Oka and the helpful advice of Drs. Yasunori Yoshida, Yoshifumi Shirakami, and Kazuhiro Shiba.
- © 2011 by Society of Nuclear Medicine
REFERENCES
- Received for publication November 30, 2010.
- Accepted for publication January 31, 2011.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Detection of Additional Primary Neoplasms on 18F-Fluciclovine PET/CT in Patients with Primary Prostate Cancer
- Update on 18F-Fluciclovine PET for Prostate Cancer Imaging
- The Utility of PET/CT in the Planning of External Radiation Therapy for Prostate Cancer
- Glutamine Transporters Are Targets of Multiple Oncogenic Signaling Pathways in Prostate Cancer
- Evaluation of Prostate Cancer with Radiolabeled Amino Acid Analogs
- Anti-1-Amino-3-18F-Fluorocyclobutane-1-Carboxylic Acid: Physiologic Uptake Patterns, Incidental Findings, and Variants That May Simulate Disease
- Comparative Evaluation of 18F-Labeled Glutamic Acid and Glutamine as Tumor Metabolic Imaging Agents
- Preparation and Characterization of L-[5-11C]-Glutamine for Metabolic Imaging of Tumors
- PET Imaging of Glutaminolysis in Tumors by 18F-(2S,4R)4-Fluoroglutamine