


Abstract
Whether preclinical depression is one of the pathophysiologic features of Alzheimer disease (AD) has been under debate. In vivo molecular imaging helps clarify this kind of issue. Here, we examined in vivo changes in the brain serotoninergic system and glucose metabolism by scanning early- to moderate-stage AD patients with and without depression using PET with a radiotracer for the serotonin transporter, 11C-3-amino-4-(2-dimethylaminomethylphenylsulfanyl) benzonitrile (DASB), and a metabolic marker, 18F-FDG. Methods: Fifteen AD patients (8 nondepressed and 7 depressed) and 10 healthy subjects participated. All participants underwent 3-dimensional MRI and quantitative 11C-DASB PET measurements, followed by 18F-FDG PET scans in the AD group. Region-of-interest analysis was used to examine changes in 11C-DASB binding potential estimated quantitatively by the Logan plot method in the serotonergic projection region. In addition, statistical parametric mapping was used to examine whether glucose metabolism in any brain region correlated with levels of 11C-DASB binding in the dense serotonergic projection region (striatum) in AD. Results: Psychologic evaluation showed that general cognitive function (Mini-Mental State Examination) was similar between the 2 AD subgroups. Striatal 11C-DASB binding was significantly lower in AD patients, irrespective of depression, than in healthy controls (P < 0.05, corrected), and 11C-DASB binding in other dense projection areas decreased significantly in the depressive group, compared with the control group. The 11C-DASB binding potential levels in the subcortical serotonergic projection region correlated negatively with depression score (Spearman correlation, P < 0.01) but not with dementia score. Statistical parametric mapping correlation analysis showed that glucose metabolism in the right dorsolateral prefrontal cortex was positively associated with the level of striatal 11C-DASB binding in AD. Conclusion: The significant reduction in 11C-DASB binding in nondepressed AD patients suggests that presynaptic serotonergic function is altered before the development of psychiatric problems such as depression in AD. The depressive AD group showed greater and broader reductions in binding, suggesting that a greater loss of serotonergic function relates to more severe psychiatric symptoms in the disease. This serotonergic dysfunction may affect the activity of the right dorsolateral prefrontal cortex, a higher center of cognition and emotion in AD.
Alzheimer disease (AD) is a neurodegenerative disorder with progressive memory and cognitive deterioration followed by or concomitant with psychologic problems such as depression and hallucination, which have a strong negative impact on the course of the disease. The occurrence of these psychologic disturbances has been reported to closely relate to disruption of the serotonergic system (1). Loss of neurons in the serotonergic raphe nuclei (2) and dysfunction of its nerve terminals in the neocortex (3) have been reported in AD. Many lines of evidence support this serotonin (5HT) deficiency theory concerning the psychobehavioral symptomatology of AD, as examined in postmortem (4–6) and pharmacotherapeutic (7,8) studies. A caveat of the postmortem studies is that the findings do not always reflect the antemortem conditions of AD because of variations of clinical symptoms after a mixture of nootropic and psychiatric drugs administered during the patient's lifetime. In addition, the comorbidity of intrinsic depression in patients with AD may be a confounding factor in a study of in vivo serotonergic changes in AD. In vivo studies using PET so far have focused on 5HT receptors in the limbic brain region in association with mnemonic cognitive impairment in AD (9–11). Thus, the alterations in presynaptic 5HT function relative to psychologic and behavioral problems in AD with and without depression remain to be investigated.
The serotonergic projections from the dorsal raphe nucleus are dense in the basal ganglia and thalamus (12,13). The 5HT transporter is a component of 5HT presynaptic neurons and a therapeutic target of selective 5HT reuptake inhibitors, which are presently the first-choice treatment for AD patients with depression (1). Because the 5HT transporter located on the presynaptic 5HT terminal regulates 5HT signaling, levels of 11C-3-amino-4-(2-dimethylaminomethylphenylsulfanyl) benzonitrile (DASB) binding in these regions reflect the activity of the presynaptic 5HT neurons chiefly in the dorsal raphe nuclei. A recent postmortem study showed a marked reduction in the binding of 5HT transporter tracer in the prefrontal cortex regardless of the presence of depression in AD (14). This finding suggests that the loss of raphe nuclei neurons and cortical 5HT transporter is one of the pathologic features of AD at the time of death. In addition, a recent SPECT study on AD with depression showed significantly lower perfusion in the dorsolateral prefrontal cortex (DLPFC) in patients with depressive symptoms than in patients without such symptoms (15). The DLPFC is known as a neural substrate of executive function that is consistently compromised in depression (16). Furthermore, patients with frontal glucose hypometabolism exhibit a faster deterioration of cognitive abilities (17) than do patients with glucose hypometabolism only in the posterior brain. Thus, a combination study of the presynaptic 5HT system and cerebral cortical metabolism is important in elucidating the underlying mechanism for AD with psychotic deterioration.
For this purpose, we measured 2 biomarkers on the same day in mild- to moderate-stage AD patients with and without depression to investigate the levels of presynaptic serotonergic function and cortical neuronal activity using PET with 11C-DASB, a specific 5HT transporter marker, and 18F-FDG.
MATERIALS AND METHODS
Participants
A total of 15 nootropic- and antipsychotic-naïve patients with AD (8 men, 7 women; mean age ± SD, 61.3 ± 5.9 y) and 10 age-, sex-, and education-matched healthy control subjects (5 men, 5 women; mean age, 55.8 ± 8.8 y) participated in the study. The diagnosis of AD was based on the criteria of the National Institute of Neurologic and Communicative Disorders and Stroke and the Alzheimer Disease and Related Disorders Association. The clinical dementia rating in all patients ranged from 1.0 to 1.5, and no patient had psychiatric disorders before the onset of dementia. The participants were evaluated with the Mini-Mental State Examination, the affect test for emotional cognition (subjects evaluate facial expressions on different cards by choosing appropriate answers from the basic emotions: happiness, sadness, surprise, disgust, anger, fear; the full score = 20) (18), and the geriatric depression scale (GDS, maximum = 15) for AD patients (19). AD patients were classified into 2 subgroups (nondepressed and depressed) according to the GDS scores as shown in Table 1. The present study was approved by the local Ethics Committee of the Hamamatsu University School of Medicine, and written informed consent was obtained from all participants themselves. We obtained written approval from the spouses or family members of AD patients as well.
Clinical Demographics
MRI and PET Procedure
All participants first underwent 3-dimensional MRI just before the PET measurement. We used a static magnet (0.3 T MRP7000AD; Hitachi) with 3-dimensional mode sampling to determine the brain areas for setting the regions of interest (ROIs). The MRI measurements and a mobile PET gantry allowed us to reconstruct PET images parallel to the intercommisural line without reslicing. Using this approach, we were able to allocate ROIs to the target regions of the original PET images (20).
PET was performed as described previously (20) on a high-resolution brain SHR12000 tomograph (Hamamatsu Photonics K.K.) having an intrinsic resolution of 2.9 × 2.9 × 3.4 mm in full width at half maximum, 47 slices, and a 163-mm axial field of view. After head fixation using a thermoplastic face mask and a 10-min transmission scan for attenuation correction, serial scanning (4 × 30 s, 20 × 60 s, and 14 × 300 s) with periodic arterial blood sampling was performed for 92 min after a slow bolus injection (taking 1 min) of a 300-MBq dose of 11C-DASB with a specific activity of more than 90 GBq/μmol. The method was the same as described previously for another 5HT transporter tracer, 11C-McN5652 (21). After completion of the 11C-DASB measurement (after 5 times the half-life of 11C had elapsed), 10 min of emission data were acquired under resting conditions 50 min after injection of 90 MBq of 18F-FDG, according to a previously described autoradiographic technique (22).
Image Data Analysis
To determine brain ROIs within the 5HT projection system, we used original morphologic MRI data instead of using spatially normalized MRI data. Most of the cell bodies are in the raphe nuclei in the midsagittal brain stem, with the largest collection of serotonergic neurons residing in the dorsal and median raphe nuclei of the caudal midbrain (23). The use of original MR images allowed us to demarcate ROIs on the region covering the substantia nigra (one of the 5HT projection area) on the MR image. In addition, multiple semicircular ROIs (∼36–120 mm2) were drawn bilaterally over the nucleus accumbens, thalamus, putamen, amygdala, prefrontal (Brodmann area 9) and temporal (Brodmann area 21) cortices, and cerebellum on the MR images in reference to an MRI atlas (24). These ROIs were then transferred onto the corresponding dynamic 11C-DASB images with 6.8-mm slice-thickness data, generated after adding 2 consecutive slices using image-processing software (Dr View; Asahi Kasei Co.) on a workstation (Ultraspark 300; SUN Microsystems) (20).
In the ROI analysis, 11C-DASB binding was quantified using a 2-tissue-compartment model, in which the cerebellar hemisphere was chosen as a reference region. The binding potential (BP) in each target region was calculated by the formula (target tissue Vd)/(cerebellum Vd) − 1, where each Vd (the volume of distribution) was obtained by the Logan graphical method (25). Because the pontine and medullary raphe nuclei send a proportion of 5HT axons into the cerebellum, this region is not an ideal candidate for a 5HT-free region. However, the use of the cerebellum as the reference region is acceptable in the present groups because smaller changes in the 5HT system were reported in the cerebellum (26).
In voxelwise analysis of 18F-FDG data, all original PET data were converted into semiquantitative parametric images using a unit of standardized uptake denoting the tracer activity per injected dose normalized to body weight. Correlation analysis by statistical parametric mapping (SPM) was performed to examine neural correlates with the level of 11C-DASB binding in the dense 5HT projection area—putamen in the AD group.
Statistics
First, right and left values for the ROIs in each region were averaged because no significant difference was observed between the interhemispheric values with the Student t test. Then, the regional 11C-DASB BP values were compared among the 3 groups with 1-way ANOVA using a post hoc Student–Newman–Keuls test. Statistical significance was set at P < 0.05 because post hoc multiple comparisons were performed in the analyses. In addition, the Kendall τ test was performed to compare the regional 11C-DASB BP values with the clinical variables in the AD group. The level of significance for the Kendall τ test was set at P < 0.01.
For the voxelwise mapping analysis, SPM software was used (SPM2; Wellcome Department of Cognitive Neurology). The standardized uptake value–based 18F-FDG images were then smoothed with an isotropic gaussian kernel of an 8 × 8 × 8 mm filter, as described elsewhere (27). Voxel-based correlation between cerebral 18F-FDG uptake and the 11C-DASB BP in the dense 5HT projection area was computed by covariance analysis, using age as a confounding covariate. The statistical threshold was set at P < 0.001, and P values were left uncorrected.
RESULTS
Levels of 11C-DASB Binding in the 3 Groups
One-way ANOVA showed that the levels of 11C-DASB BP in the dense 5HT projection regions (midbrain, nucleus accumbens, putamen, and thalamus) of the depressed AD group and the level in the putamen of the nondepressed AD group were significantly lower than those in the control group (Table 2; Fig. 1). 11C-DASB BP tended to be reduced in other dense projection areas of the 5HT system (nucleus accumbens, thalamus, and midbrain) in the nondepressed AD group. No significant reduction was found in the cortical areas of the AD group. No significant difference in 11C-DASB binding was found between the nondepressed and depressed AD groups, although the depressed AD group showed a clear tendency toward lower levels. Representative 11C-DASB PET images of subjects in each group are shown in Figure 1.

Levels of 11C-DASB binding within dense 5HT projection areas in midbrain (A), putamen (B), and thalamus (C). Representative PET images of 11C-DASB binding relative to cerebellar uptake are shown. Color scale denotes magnitude of tissue-to-cerebellum ratio, ranging from 0 to 3. NC = healthy control; AD-n = nondepressive AD; AD-d = depressive AD. *P < 0.05, ANOVA with post hoc Student–Newman–Keuls test.
Levels of 11C-DASB BP in the 3 Groups
Correlation Between 11C-DASB Binding and Clinical Parameters in AD Group
As shown in Figure 2A, the levels of 11C-DASB BP in all dense 5HT projection regions correlated negatively with GDS scores (midbrain: r = 0.692, P < 0.01, f(x) = −0.057x + 1.834; putamen: r = 0.72, P < 0.005, f(x) = −0.046x + 1.683; thalamus: r = 0.839, P < 0.005, f(x) = −0.046x + 1.638), indicating that lower binding of 11C-DASB in the 5HT system may be related to greater deterioration of the emotional state in AD. Although no significant correlation was found with the scores for dementia (Mini-Mental State Examination, Fig. 2B) or emotional cognition (affect test, Fig. 2C), there was a tendency toward positive correlations between the putaminal 11C-DASB BP and the Mini-Mental State Examination scores and between subcortical 11C-DASB BPs and the affect scores.

Correlations between levels of 11C-DASB BP in midbrain (▵), putamen (○), and thalamus (•) for GDS score (A), Mini-Mental State Examination (MMSE) score (B), and affect score (C) in all AD patients. Dotted (putamen), straight (thalamus), and dashed (midbrain) lines denote significant correlations (Kendall rank correlation, P < 0.01).
Neural Correlates of Cerebral Glucose Metabolism with Striatal 11C-DASB Binding in AD Group
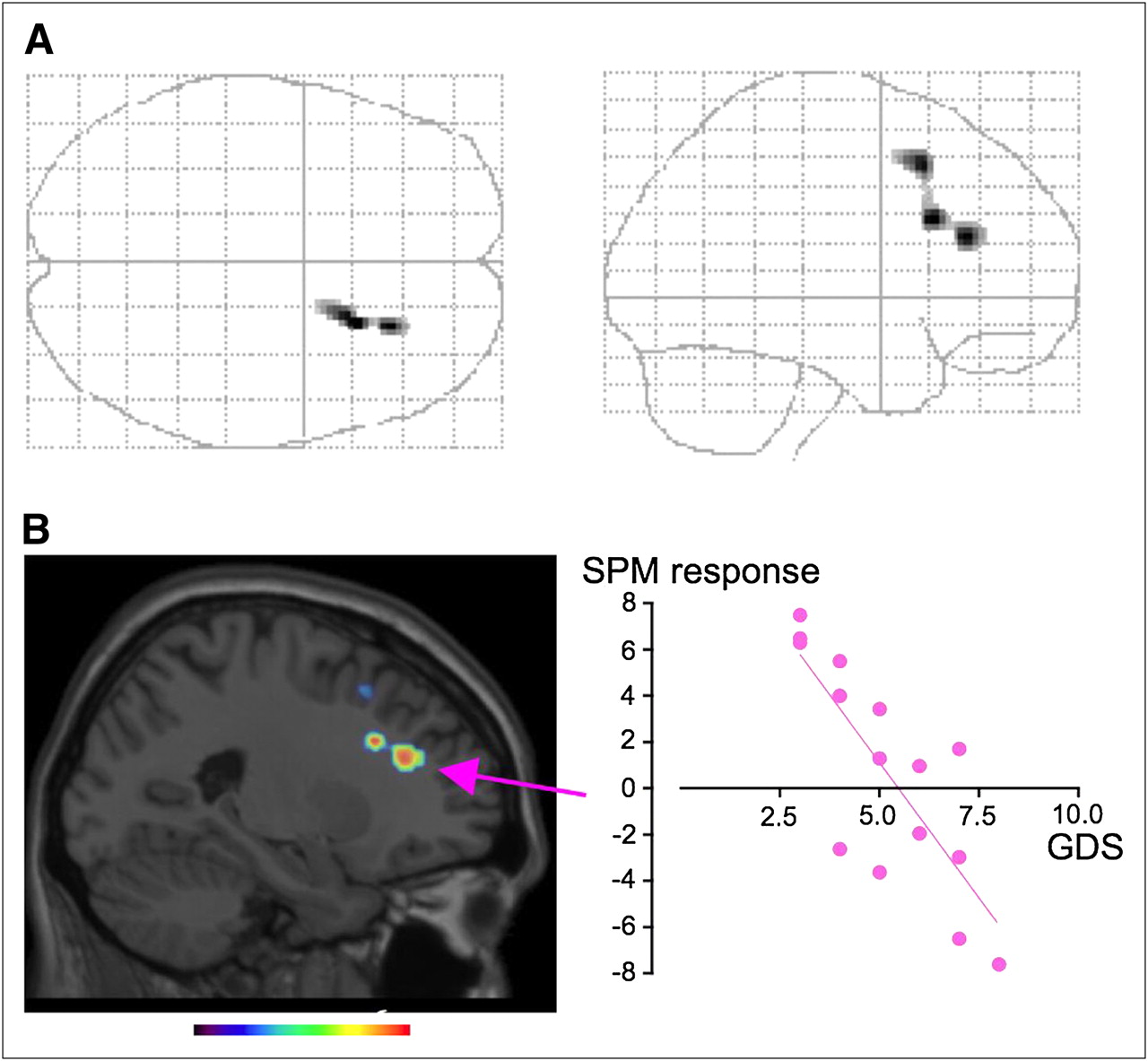
SPM correlation analysis showed that glucose metabolism in the right DLPFC and superior frontal gyrus correlated positively with putaminal 11C-DASB binding in AD patients (glass brain: Fig. 3A; Table 3). No other significant correlation was found, but there was a tendency toward a correlation when either 11C-DASB BP in the thalamus or BP in the midbrain was chosen as a covariate (data not shown). The glucose metabolic level in the statistically highest peak region (right DLPFC) statistically correlated negatively with GDS scores in AD patients (Fig. 3B, scattergram, r = 0.798, P < 0.0004, f(x) = −2.34x + 12.8). Between-group comparison of SPM failed to show any brain region with statistically significant differences in glucose metabolism between the nondepressed and depressed groups (data not shown).
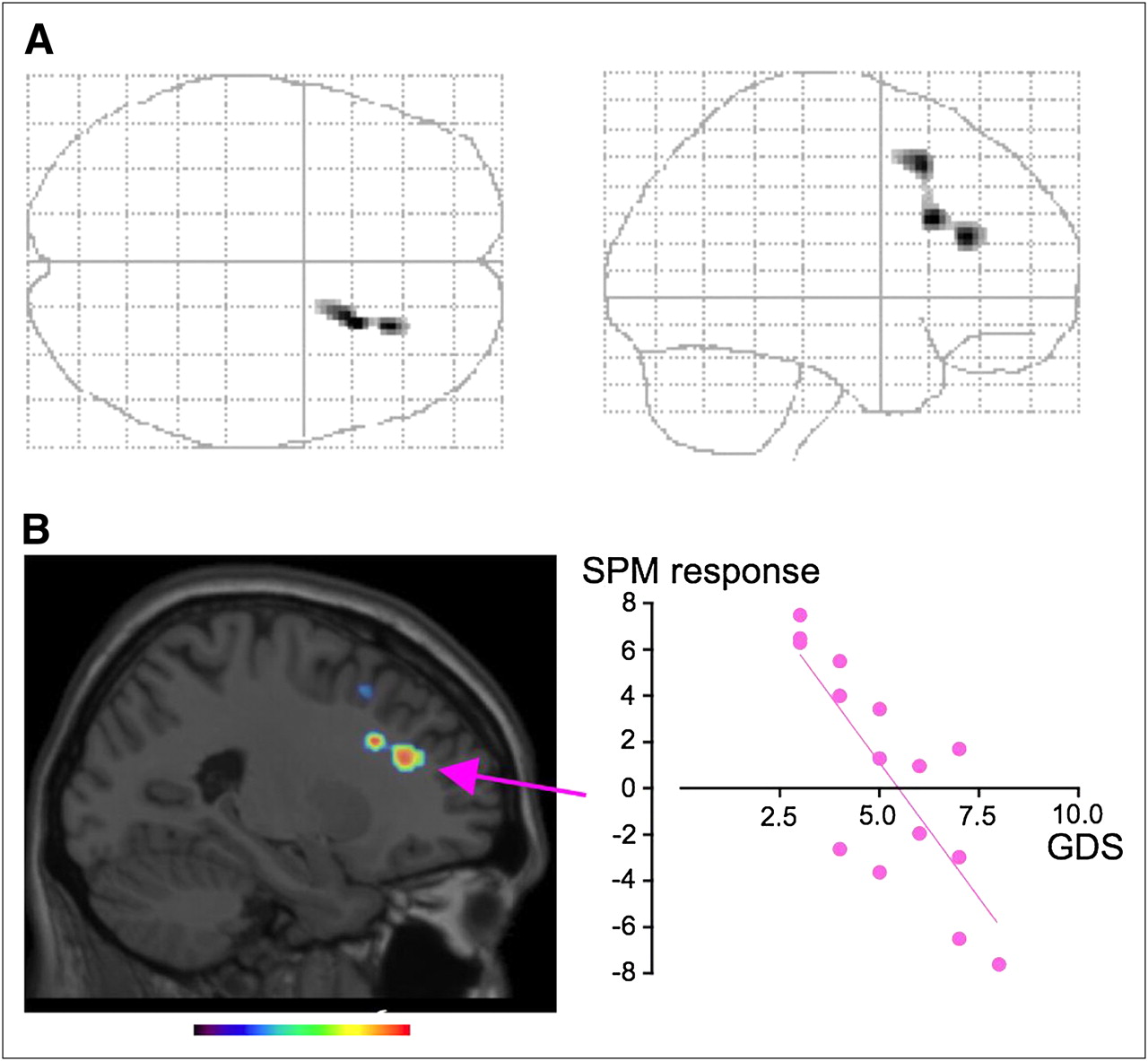
SPM images in AD group. (A) SPM correlation analysis showed positive correlation between striatal 11C-DASB BP and glucose use in right DLPFC (glass brain, P < 0.001, uncorrected). (B) SPM responses in area with highest peak (arrow) correlated negatively with GDS scores. Color scale bar denotes t value.
Brain Regions Showing Significant Correlation Between Glucose Metabolism and Striatal 11C-DASB Binding in AD
DISCUSSION
The present results showed a significant reduction in binding of the 5HT transporter marker 11C-DASB in subcortical 5HT projection areas (especially the striatum) in patients with early- to moderate-stage AD regardless of depression and that the reduction was associated with the severity of their depressive states. The brain mapping analysis depicted the right DLPFC as a neural correlate of raphe-striatal 5HT activity in the disease. Furthermore, DLPFC metabolism was found to correlate with depressive scores, suggesting that 5HT dysfunction might affect right DLPFC activity, possibly leading to the generation of a clinical phenotype depression in AD. Thus, as suggested ex vivo (3,28), a progressive 5HT dysfunction may be present in the living brain of patients even in the absence of depression and may worsen the state of emotion.
In the present study, 11C-DASB binding in the basal ganglia and thalamus in AD patients was about 25% that in healthy subjects—a smaller reduction than the loss of 5HT cell density observed in the raphe nucleus of postmortem AD brains (45%) (29). Although this difference might be due to differences between antemortem and postmortem specimens, a previous finding that marked 5HT cell attrition occurs regardless of the presence or absence of depression (29) and our finding that striatal 11C-DASB binding is reduced in parallel with GDS scores suggest that 5HT neuronal reduction and psychologic deterioration may covary up to the last stage of 5HT neuronal loss in AD patients. Indeed, the 5HT neurons may be affected early in the course of the disease because AD patients with GDS scores of around zero showed a marked reduction in 11C-DASB binding specifically in the putamen, compared with that in healthy subjects (Fig. 1B). This finding is in contrast to the increase in binding of 5HT transporter marker 11C-McN5652 in major depression, (30) suggesting that the underlying etiologies are clearly different. The mechanism of 5HT neuronal degeneration in AD remains unclear, but previous reports about the extent of 5HT neuron loss in the dorsomedial raphe nuclei (28) without association between 5HT transporter polymorphism and depression in depressed AD patients (31) suggest that the 5HT neuronal degeneration seems acquired. This may be true, because aggregation of Aβ protein exerts an early, focal neurotoxic effect on 5-HT and ACh axons (32).
It was reported that glucose metabolism in the DLPFC was lower in depressed AD patients than in their nondepressed counterparts (33). In the present study, we found a significant correlation between striatal 11C-DASB binding and DLPFC glucose metabolism (Fig. 3A), the magnitude of which was associated with the extent of clinical deterioration and depression severity. Although striatal 11C-DASB binding was lower in depressed AD patients in the present study, SPM analysis performed separately in each subgroup of AD (nondepressed and depressed) failed to show statistical significance for any brain region (data not shown). This result may be ascribed to the limited number of patients in each group or the narrow variation in 11C-DASB BP in each subgroup. Previous studies showing that the DLPFC was involved in cognitive processing in negative emotion (34) and that successful 5HT augmentation therapy activated prefrontal glucose metabolism (35) would support our finding that striatal 5HT activity is associated with right DLPFC metabolism in AD patients. This right-sided dominancy may reflect a depression-related susceptibility because lesions in the right frontal cortex lead to anxiety and depression (36) and because recent memories preferentially involve the right prefrontal cortex in AD (37). Thus, there might be a link between this important clinical phenomenon and a 5HT-related alteration in right prefrontal activity.
In the present study, no significant correlation was found between presynaptic 5HT dysfunction and dementia, whereas the scattergrams of Figures 2B and 2C showing a weak tendency toward a correlation may hint that a greater number of patients would exhibit significant correlations in presynaptic 5HT activity and these cognitive abilities. In addition, the limited number of clinical features in AD in the current study may be responsible for the failure in clinicobiochemical correlation. Previous reports showing that a reduction in 5HT signaling causes synaptic dysfunction and neuronal death in AD (38) and that enhanced 5HT signaling reduces levels of Aβ protein in the brain of transgenic AD mice (39) would support this speculation. At the moment, however, it is unlikely that the 5HT system is directly involved in the generation of dementia, as opposed to the implications of cholinergic failure in the AD brain. Because a history of depression is a risk factor of AD, alteration in the 5HT system even at an early stage of the disease should be borne in mind in the management of AD patients. Indeed, in the clinical setting, AD patients with neuropsychiatric symptoms tend to be institutionalized sooner than those without (40). Thus, on top of anticholinergic and antiamyloid deposition therapy, enhancement of 5HT signaling by, for example, selective 5HT reuptake inhibitors may be important in the treatment and prophylaxis of AD.
CONCLUSION
This study suggests that a degree of presynaptic 5HT function in the subcortical 5HT projection region is compromised in AD patients even before the development of depression. Right DLPFC dysfunction in parallel with 5HT inactivation is also implicated in the progression of emotional and cognitive deterioration in AD.
Acknowledgments
We thank Dr. Mitsuo Kaneko (Kaneko Clinic), Dr. Masanobu Sakamoto (Hamamatsu Medical Center), Toshihiko Kanno (Hamamatsu Medical Center), and Yutaka Naito (Japan Environment Research Corporation) for their clinical and technical support. This work was supported by a Research Grant for Longevity Science from the Ministry of Health, Labor, and Welfare, Japan.
Footnotes
-
COPYRIGHT © 2009 by the Society of Nuclear Medicine, Inc.
References
- Received for publication February 9, 2009.
- Accepted for publication April 2, 2009.





{kind=link}
{kind=link}
{kind=link}