


Abstract
Transfer of the human sodium iodide symporter (hNIS) has been proposed as a new principle of cancer gene therapy. This study evaluates the iodide kinetics and dosimetry of iodide in hNIS-expressing thyroid carcinoma cells under optimized conditions. Methods: Using a bicistronic retroviral vector for the transfer of the hNIS and the hygromycin resistance gene, hNIS-expressing rat thyroid carcinoma cell lines were generated. Afterward, Na125I uptake and efflux were determined in genetically modified and wild-type cells in the presence or absence of modulators of iodide transport. In addition, the 131I distribution in thyroid-ablated nude mice bearing wild-type and genetically modified thyroid carcinomas was monitored after intraperitoneal administration of 131I with and without coadministration of lithium carbonate. Results: hNIS-expressing cell lines accumulated up to 49 times more iodide than did noninfected cells, with a maximal iodide uptake after 30 min of incubation. However, a 90% efflux of the radioactivity occurred 20 min after replacement of the medium. In mice, the hNIS-expressing tumors accumulated up to 23 and 19.5 times more iodide than did the wild-type tumors in lithium-treated and control animals, respectively. However, efflux of the radioactivity was also observed in vivo: After 24 h, hNIS-expressing tumors lost 82.5% and 80.4% of the initial activity. Dosimetric calculations showed that 1,650 MBq of 131I per square meter resulted in 5.4 and 5.2 Gy in hNIS-expressing tumors and 0.24 and 0.26 in wild-type tumors. Conclusion: Transduction of the hNIS gene in rat thyroid carcinoma cells induces iodide transport, which is associated with rapid efflux. Application of 131I in clinically relevant amounts did not result in therapeutically useful absorbed doses in hNIS-expressing tumors in vivo, even under optimized conditions of thyroid ablation and treatment with lithium carbonate.
Currently used viral vectors for gene therapy of cancer have a low infection efficiency leading to moderate or low therapy effects. This problem could be solved using an approach that leads to accumulation of radioactive isotopes with β-emission. In this case, isotope-trapping centers in the tumor could create a cross firing of β-particles, thereby efficiently killing transduced and nontransduced tumor cells.
Since the cloning of the human and rat complementary DNA sequences, several experimental studies have investigated iodide accumulation after recombinant expression of the human sodium iodide symporter (hNIS) gene in malignant tumors (1–18). Although initial uptake is high in the genetically modified tumors, differing results have been reported for the efficiency of radioiodine treatment based on hNIS gene transfer. These differences may be due to the different experimental protocols, such as whether thyroid ablation is used, the administration route of the tracer, the doses given to the animals, or the tumor and animal models used. To optimize the efficiency of radioiodide therapy, modulation of iodide kinetics has been applied. A higher accumulation in genetically modified tumors in animals after thyroid ablation has been described by Smit et al. (16). Furthermore, lithium has been used in vitro as well as in clinical studies to prolong iodide retention in tumor cells or in benign thyroid disease. This study combines thyroid ablation, intraperitoneal administration, and lithium treatment with transfer of the hNIS gene to assess iodide kinetics and dosimetry during radioiodine therapy under these optimized conditions.
MATERIALS AND METHODS
Retroviral Transfer of the hNIS Gene and Generation of Recombinant Cell Lines
For transfer of the hNIS gene, a bicistronic retroviral vector was used consisting of the hNIS gene and the hygromycin resistance gene cloned downstream of the elongation factor 1α promoter. To ensure simultaneous expression of the genes coding for the hNIS and for the hygromycin resistance and stabilization of the messenger RNA, a synthetic intron and an internal ribosomal entry site from encephalomyocarditis virus was inserted between the genes (7,8). The rat papillary thyroid carcinoma cell line L2, which does not concentrate iodide ((19) obtained from Yoshio Hiasa, Nara, Japan), was maintained in Ham’s F12 medium modified by addition of insulin (10 μg/mL), hydrocortisone (10 nU), transferrin (5 μg/mL), somatostatin (10 ng/mL), glycyl-histidyl-lysin (10 ng/mL), and 5% fetal calf serum. The transient packaging cell line BOSC23 used for the production of ecotropic retroviral particles was cultured in RPMI1640 medium (Gibco BRL) supplemented with glutamine (292 mg/L), penicillin (100,000 units/L), streptomycin (100 mg/L), and 10% fetal calf serum (20). The cell lines were cultured at 37°C, in an atmosphere of 95% air and 5% CO2.
For transient packaging of the retroviral particles, a lipofection of BOSC23 cells was done. After 2 d, the medium was centrifuged to remove detached BOSC23 cells and used for the infection of L2 cells in the presence of 8 μg of polybrene per milliliter overnight. The cells were treated with hygromycin (425 μg/mL) for 4 wk until 13 resistant cell lines were established. These were used in iodide uptake experiments. The cell line that presented the maximal 125I− uptake (L2hNIShyg3) was used for the following experiments. The tumor doubling time in vitro was 15 ± 2.3 h for L2 and 14.5 ± 1.3 h for L2hNIShyg3.
Measurement and Modulation of 125I Uptake and Efflux
Iodide uptake was determined in triplicates as described previously (7,21), and the experiments were repeated twice. In the presence of 74 kBq of Na125I (Amersham Buchler Germany; specific activity, 625.3 MBq/μg; radioactive concentration, 3.7 GBq/mL; radiochemical purity, 99.3%), the recombinant cell line L2hNIShyg3 and wild-type L2 cells were cultured for 1, 2, 5, 10, and 30 min or 1, 2, and 4 h in 6 well plates. After being washed twice with ice-cold phosphate-buffered saline, the cells were lysed with sodium hydroxide on ice. With an automated NaI(Tl) well counter (Cobra II; Canberra Packard), the radioactivity was measured in cell lysates and in the medium. The viable cell number was determined in a Coulter counter (Coulter Electronics) and by trypan blue staining (more than 94% viable cells). The Coulter counter measured a median cell volume of 2.05 fL (range, 1.07–4.2 fL) for wild-type L2 cells and 1.96 fL (range, 1.11–4.05 fL).
For modulation of iodide uptake, hNIS-expressing and wild-type cells were incubated for 1 h in Na125I medium (74 kBq) or Na125I medium supplemented with sodium perchlorate (10 or 50 μmol/L; Sigma-Aldrich), ouabain (0.5 mmol/L), the anion channel blocker 4,4′-diisothiocyano-2,2′-disulfonic acid stilbene (DIDS, 100 or 300 μmol/L; Sigma-Aldrich), or the proton conductor carbonyl cyanide p-trifluoromethoxyphenylhydrazone (FCCP, 10 μmol/L; Sigma-Aldrich) according to Weiss et al. (21). Thereafter, the cells were washed, lysed, and counted as described.
To determine the 125I− efflux, recombinant and wild-type cells were incubated for 1 h in the presence or absence of 300 μmol of DIDS per liter, with medium containing 74 kBq of Na125I in 6 well plates. After the cells had been washed twice with phosphate-buffered saline, 3 wells were lysed immediately. To the other wells, fresh nonradioactive medium was added and the cells were again incubated for 2, 4, 6, 8, 10, 12, 16, or 20 min and washed and lysed as described.
Measurement of 131I Uptake in Tumor Tissue of Nude Mice
The experiments were performed in compliance with German laws relating to the conduct of animal experimentation. Swiss CD1 nu/nu mice (Charles River Laboratories, delivered by Iffa Credo) weighing 20–25 g were first treated with 11 MBq (300 μCi) of 131I given intraperitoneally to obtain thyroid ablation. Two weeks later, 2 × 106 tumor cells in 100 μL were transplanted subcutaneously into the right (L2hNIShyg3) or left (wild-type L2) thigh of the animals. At 4 wk after transplantation, 2 groups of animals were treated: group 1 (n = 16) received 0.5 mL of a physiologic sodium chloride solution intraperitoneally. One day later, 15 MBq (400 μCi) of 131I, corresponding to a dose of 1,650 MBq/m2 (calculated according to (8,22)), were given intraperitoneally together with a second dose of sodium chloride. Group 2 (n = 16) received lithium carbonate (370 mg/m2) in 0.5 mL of solution intraperitoneally on day 1. One day later, 15 MBq (400 μCi) of 131I were given intraperitoneally together with lithium carbonate (185 mg/m2) in 0.5 mL of solution. The animals were sacrificed at 2, 6, 24, and 48 h after injection to analyze the absolute amount of radioactivity (percentage injected dose per gram of wet tissue) in tumors and other tissues using an automated NaI(Tl) well counter (Cobra II; Canberra Packard). MIRDOSE3* (23) was used to calculate the cumulated activity and the absorbed dose in wild-type and hNIS-expressing tumors after administration of 1,650 MBq/m2. 131I was calculated for a unit density sphere with a mass of 1 g, considering only the value of the absorbed dose in the sphere. Therefore, the sum of 2 exponential decay functions was fitted to the data points. The MIRDDOSE3* calculation of S-value for self-irradiation was 29.5 μGy/MBq·s.
For imaging studies performed with the animal under general gaseous anesthesia (40% O2/60% N2O/1% halothane) using a 25.4-cm (10 in.) scintillation camera (Searle-Siemens), only animals bearing tumors of a minimum of 15-mm diameter were accepted. The time-dependent relative accumulation of radioactivity in different regions of interest, such as the heart, liver, tumor, bladder, and whole animal, was monitored in 6 animals at 1, 2, 4, 8, and 24 h after injection.
The animal data were statistically analyzed by the t test, using SigmaPlot (version 8.0; Jandel Scientific).
RESULTS
Na125I Uptake and Efflux in Wild-Type and Recombinant L2 Cell Lines
With respect to the wild-type counterparts, up to 49-fold more iodide was transported into the recombinant thyroid carcinoma cells. The initial uptake of iodide was dependent on the incubation time, and cells accumulated 125I− maximally after 10–30 min of incubation. The radioactivity measured after 1 and 4 h of 125I− incubation was at a plateau level, implicating steady-state uptake (Fig. 1).
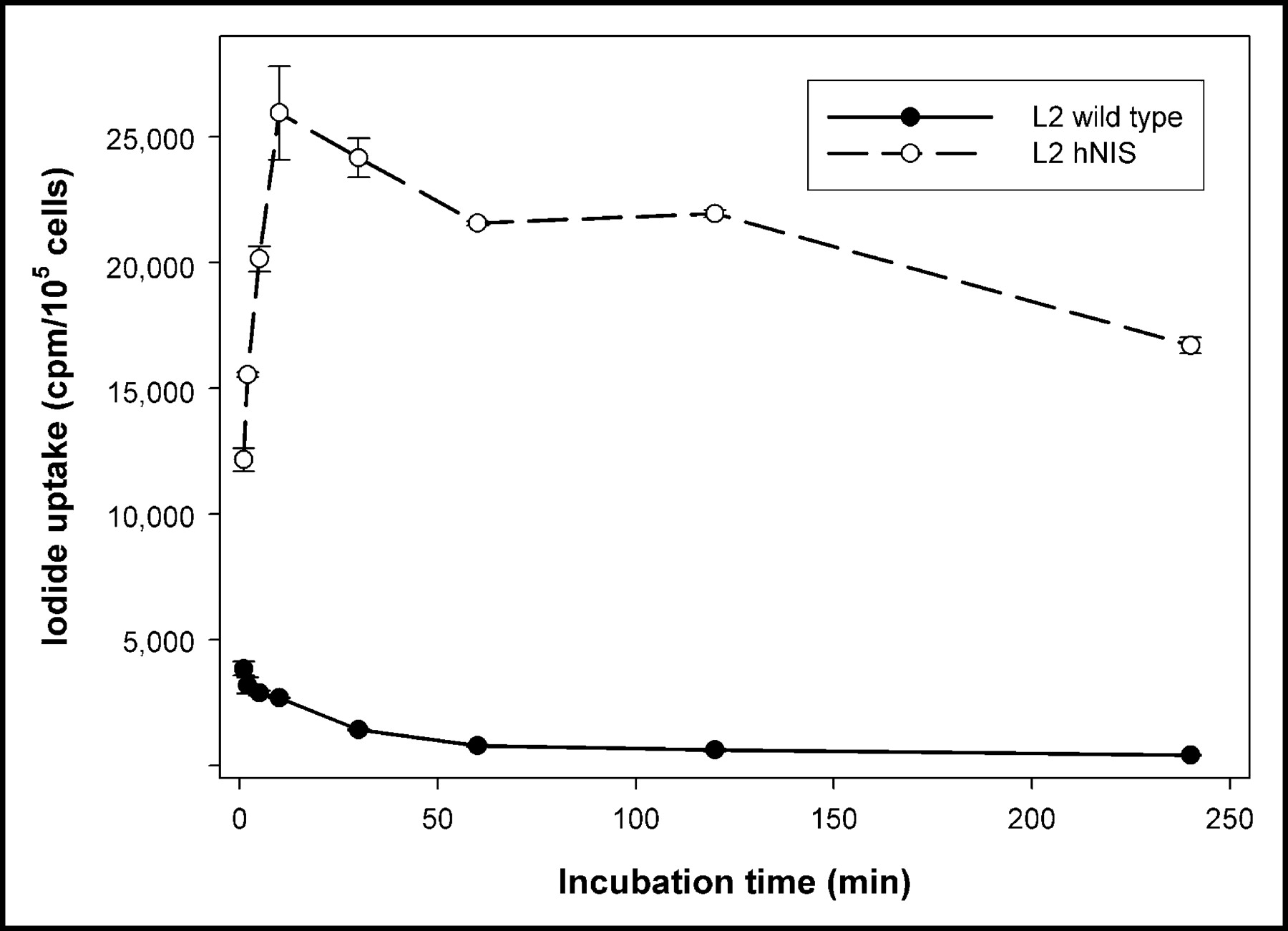
Time dependence of 125I− uptake in wild-type rat thyroid carcinoma cells and in the hNIS-expressing cell line L2hNIShyg3. Values are mean and SD (n = 3).
Cell function was characterized by competition and efflux experiments. Figure 2 presents the effect of DIDS, FCCP, sodium perchlorate, and ouabain on Na125I uptake in the wild-type and the hNIS-expressing cells. In the presence of 10 or 50 μmol of sodium perchlorate per liter, we observed a dose-dependent inhibition of iodide accumulation—of 87% and 92%, respectively—in the L2hNIShyg3 cells. Ouabain caused a 75% inhibition, whereas addition of the proton conductor FCCP led to a 75% loss of the accumulated I−. The anion channel blocker DIDS had an opposite effect, with a 60% increase at a concentration of 300 μmol/L and a 18% decrease at a concentration of 100 μmol/L. In wild-type cells, 300 μmol of DIDS per liter caused an 86% increase at a low uptake level, whereas no changes occurred for the other modulators (Fig. 2).

Effect of DIDS, FCCP, sodium perchlorate (SP), and ouabain (ouab) on 125I− uptake in wild-type rat thyroid carcinoma cells and in the hNIS-expressing cell line L2hNIShyg3 after 1 h of incubation. Values are mean and SD (n = 3). con = control.
To evaluate iodide efflux, we allowed 125I uptake to proceed for 1 h, at which time a steady-state level of iodide accumulation was achieved. After the medium had been replaced by nonradioactive medium, the amount of 125I− present in the L2hNIShyg3 cell lysates was determined as a function of time. Up to 66% of the cellular radioactivity was released into the medium after 10 min, and 90% efflux was observed after 20 min (t1/2 value of 5.8 min), indicating that the radiotracer was not trapped in the recombinant thyroid carcinoma cells (Fig. 3). The addition of a 300 μmol/L concentration of DIDS to the culture led to an increased initial 125I− uptake; however, intracellular retention of the tracer was not prolonged, with 64% and 93% efflux at 10 and 20 min (t1/2 value of 6.7 min), respectively.

Iodide efflux from L2hNIShyg3 cells after 1 h of incubation with Na125I with and without 300 μmol of DIDS per liter. Values are mean and SD (n = 3).
131I− Uptake in Tumor-Bearing Mice
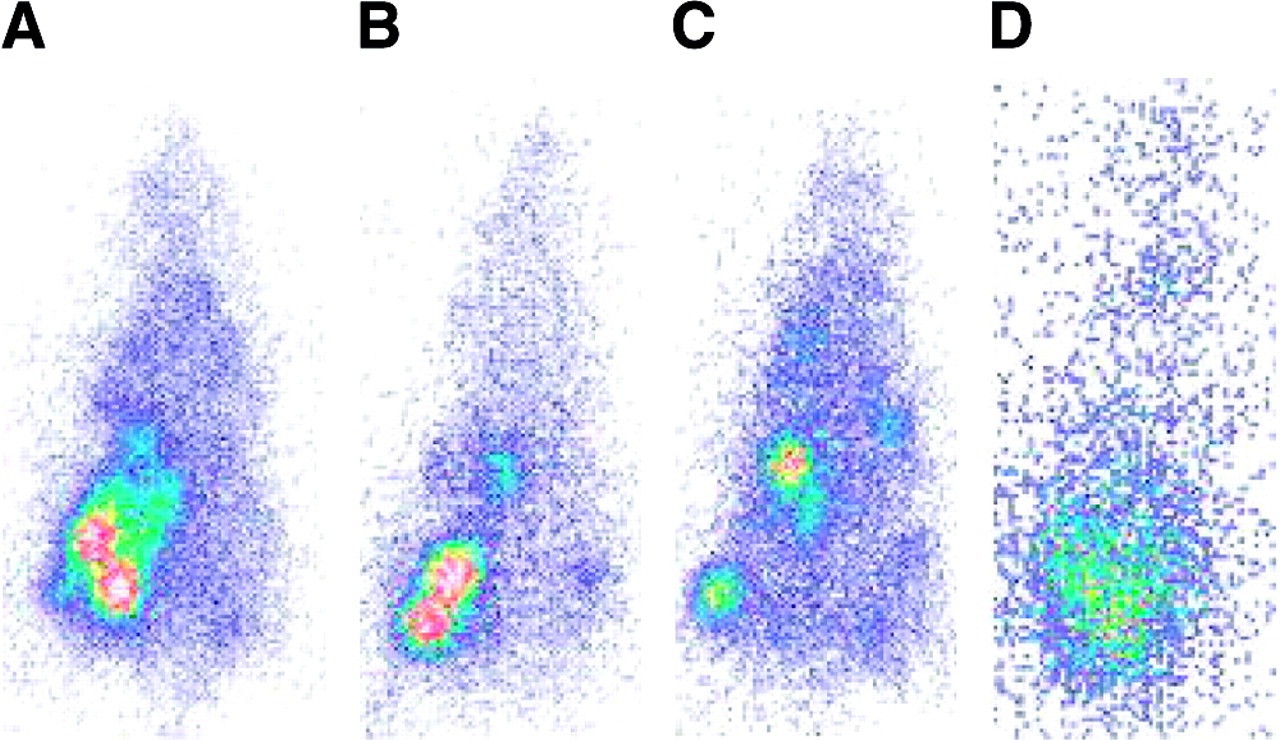
To investigate iodide uptake in hNIS-expressing or -nonexpressing thyroid carcinoma in vivo, L2hNIShyg3 cells or wild-type L2 cells were transplanted into the right and left thighs of Swiss CD1 nu/nu mice. Consistent with the data obtained from the in vitro studies, hNIS-expressing tumor tissue significantly accumulated 131I−, leading to scintigraphic visualization up to 24 h after tracer administration, whereas only low iodide uptake was observed in the wild-type thyroid carcinoma (Fig. 4). Because the animals received a thyroid ablative dose of 131I− before tumor cell transplantation, the thyroid was scintigraphically not visible. Tables 1 and 2 present the ex vivo quantitation of 131I− uptake per gram of tumor tissue, which was evaluated at different times after tracer administration. Except for the stomach (at 2 and 6 h after injection) iodide uptake was not statistically different in the organs and tumors after lithium carbonate or sodium chloride solution. Iodide accumulation was 23-fold (lithium treated) and 19.5-fold (sodium chloride) higher in hNIS-expressing tumor tissues than in wild-type tumor tissues. However, at 24 and 48 h after 131I administration, only 4.09% and 0.52% of the injected dose per gram of tissue were found in the hNIS-expressing tumors after lithium administration and 4.22% and 0.35% after sodium chloride (0.18% and 0.02%, and 0.16% and 0.02%, respectively, for wild-type tumors; Tables 1 and 2). Therefore, an efflux of radioactivity from hNIS-expressing tumors was observed at 24 h after injection, with a loss of 82.5% and 80.4% of the initial activity in lithium-treated animals and control animals, respectively (Fig. 5). At 48 h after injection, 97.8% and 98.4% of the radioactivity were lost in these tumors. The intratumoral half-life of 131I in hNIS-expressing tumors was 8.75 h in lithium-treated animals and 9.32 h in control animals. According to dosimetric calculations, 1,650 MBq/m2 resulted in 5.2 and 5.4 Gy in animals after sodium chloride and lithium carbonate administration, respectively (wild-type tumor, 0.24 and 0.26 Gy, respectively).

Scintigraphic images of tumor-bearing nude mice with subcutaneously transplanted hNIS-expressing (right thigh) or wild-type (left thigh) thyroid carcinoma cells at 2 h (A), 6 h (B), 24 h (C), and 48 h (D) after injection of 131I−.

131I− uptake (kBq/g) in hNIS-expressing and wild-type thyroid carcinomas after administration of 1,650 MBq/m2 at different times after radiotracer application in rats. Values are mean and SDs (n = 4).
Iodide Activity Concentration in Animals Treated with Lithium Carbonate at Different Times After Injection of 131I
Iodide Activity Concentration in Animals Treated with Sodium Chloride at Different Times After Injection of 131I
DISCUSSION
Transfer of the hNIS gene has been suggested as a new therapeutic principle based on 131I accumulation in malignant tumors. The hNIS gene has been transferred in a variety of tumor models using different vector systems and promoters and has resulted in a high initial uptake (3–18). Selective killing of hNIS-expressing cells has been observed in vitro using clonogenic assays, and a bystander effect has been suggested in 3-dimensional spheroid cultures (3–5,9,11,13,17). Furthermore, stably transfected human prostate carcinoma cells revealed a long biologic half-life of 45 h in vivo, resulting in 84% tumor reduction after a single intraperitoneal application of a very high, 111-MBq, dose of 131I (17,18).
However, no organification occurred, and despite initially high uptake, rapid efflux has been observed in different tumor models (7,8,12,13,15,16,24). Compared with uptake in the wild-type cell line, uptake in rat thyroid carcinomas was increased up to 49-fold, which is within the range of values reported for other models (3,5,7,10). Furthermore, a 90% efflux could be observed in vitro. The potent erythrocyte anion channel blocker DIDS increased initial uptake of iodide in the rat thyroid cell line FRTL5 but had no effect on the first component of iodide efflux, leading to the interpretation that the primary effect of DIDS is a stimulatory effect on iodide influx (21). Consequently, DIDS caused an increased initial 125I− uptake in the hNIS-expressing rat thyroid carcinoma cells; however, prolonged intracellular retention of the tracer was not achieved. Similar results have been obtained in other studies—in vitro as well as in vivo (7,8,12,15,25). Because the effectiveness of radioiodine therapy depends on the amount and biologic half-life of the isotope in the tumor, a therapeutically useful absorbed dose in vivo seems unlikely in these studies.
To optimize radioiodine therapy after hNIS gene transfer, we combined 3 different factors with a potential benefit concerning isotope retention in the tumor: thyroid ablation, intraperitoneal administration of 131I, and treatment with lithium carbonate. Smit et al. have used thyroid ablation together with a low-iodine diet to increase iodide uptake in genetically modified tumors (16). Intraperitoneal or subcutaneous administration may result in prolonged retention in the animal because of prolonged resorption, which may partly explain the long half-life of iodide observed in former studies (16–18). In contrast to previously published data on Morris hepatoma or prostate carcinoma, the hNIS-expressing thyroid carcinomas in our study were clearly visible at 24 h after intraperitoneal tracer administration. Because the hNIS gene was under the control of a promoter independent of thyroid-stimulating hormone (TSH) (elongation factor 1α), the increased TSH levels caused by thyroid ablation were not likely to be influential.
Lithium has been reported to reduce iodine release from the thyroid and was therefore used to enhance radioiodine treatment of differentiated thyroid cancer (26). In FRTL-5 rat thyroid cells and in primary cultures of porcine thyroid follicles, lithium suppressed TSH-induced iodide uptake, iodide uptake stimulated by 8-bromo-cyclic adenosine monophosphate (cAMP), iodine organification, and de novo thyroid hormone formation (27,28). A variety of actions have been described for lithium, such as inhibition of adenosine triphosphatase activity, cAMP, intracellular enzymes, and inositol phospholipid metabolism, with an effect on signal transduction, and alteration of the in vitro response of cultured cells to thyrotropin-releasing hormone (27–29). Lithium is concentrated by the thyroid and inhibits thyroidal iodine uptake, iodotyrosine coupling, and thyroid hormone secretion and alters thyroglobulin structure (27–31). Therefore, if enhancement of iodide trapping in the thyroid by lithium relies on interaction with iodine coupling to tyrosine residues or inhibition of thyroid hormone secretion, an organification process is still needed for sufficient iodine to accumulate in the tumor.
Sieger et al. performed efflux experiments on hNIS-expressing hepatoma cells in the presence of different concentrations of lithium. Initial iodide uptake was higher in lithium-treated cells than in untreated cells when lithium concentrations of 2 and 10 mmol/L were used, but still, a 62% efflux was observed after 20 min. When lithium was used at a concentration of 50 mmol/L, the initial iodide uptake in treated cells decreased by 50% in comparison with untreated cells, with a 70% efflux after 20 min (25). We observed significant efflux in rat thyroid carcinoma in vivo: Only 4.09% ± 0.38% (lithium-treated animals) and 4.22% ± 2.2% (control animals) of the injected dose per gram in the hNIS-expressing tumors were observed at 24 h after tracer administration. At 48 h, these values were reduced to 0.52% ± 0.3% and 0.35% ± 0.006%, respectively. Similarly, Nakamoto et al. found that less than 1% of the injected radioactivity remained at 24 h after 131I administration in modified MCF7 mammary carcinomas (12).
Our data corresponded to a very short half-life of 131I (8.75 and 9.32 h in lithium-treated and control animals, respectively) in rat thyroid carcinomas. A short biologic half-life—3.6 h—has also been calculated by Nakamoto et al. for human mammary carcinomas (12). In contrast, differentiated thyroid carcinoma showed a biologic half-life of less than 10 d, and normal thyroid showed a biologic half-life of approximately 60 d (32). Therefore, whether radioiodine treatment is successful after transfer of the hNIS gene alone remains questionable. Consequently, dosimetric calculations showed that 1,650 MBq/m2 resulted in 5.2 and 5.4 Gy in the hNIS-expressing rat thyroid carcinomas after sodium chloride and lithium carbonate administration, respectively (in wild-type tumor, 0.24 and 0.26 Gy, respectively). Because approximately 80 Gy have been found necessary to eliminate metastases in patients with thyroid cancer, these levels are not likely to induce a significant therapeutic effect on tumors (33). Furthermore, the experiments were performed under ideal conditions, with 100% hNIS-expressing cells in the tumors. Given the low infection efficiency of current viral vectors in vivo, the absorbed dose in a clinical study would be considerably lower.
To prolong iodide retention, the thyroperoxidase gene was transferred either alone or in combination with the hNIS gene. However, a rapid efflux of iodide was observed, indicating no or very low thyroperoxidase activity (4,24,34). A further option to improve the outcome of therapy is the use of biologically more effective isotopes, as has been done using 188Re-perrhenate or the high-linear-energy-transfer emitter 211At (12,33,35). Although all radioisotopes showed rapid efflux, higher absorbed doses in the tumor were found for 211At and 188Re (33,35).
Compared with the experiments of Spitzweg et al. (16) and Smit et al. (17,18), this study differed in the administered dose and the animal models used. The 74- and 111-MBq doses used by Smit and Spitzweg correspond to administered doses of 11,100 and 16,650 MBq/m2, respectively, in mice. These doses are far greater than doses used in patients. In our study, we used a dose that is in the range of doses administered to patients with differentiated thyroid carcinoma. Furthermore, Smit et al. injected 131I at 3 wk after transplantation, when tumors were not visible. Small tumors may have a better chance to respond because they have no hypoperfused regions with limited tracer availability. However, this is not a realistic simulation of the tumor patient. There are also differences in the animal and tumor models and their biochemical properties that may lead to differences in iodide retention. The possibility also exists that a low-iodide diet benefits isotope accumulation in the tumor (16).
CONCLUSION
In conclusion, transfer of the sodium iodide transporter gene into rat thyroid adenocarcinoma cells, together with application of 131I in clinically relevant amounts, did not result in therapeutically useful absorbed doses in hNIS-expressing tumors in vivo even under optimized conditions of thyroid ablation and treatment with lithium carbonate. Further studies have to examine pharmacologic modulation of iodide efflux or the use of the hNIS gene as an in vivo reporter gene (8,37–39).
Acknowledgments
The authors thank Ursula Schierbaum and Karin Leotta for their help in performing the animal experiments. This study was supported by a grant from the Tumorzentrum Heidelberg/Mannheim.
Footnotes
Received Oct. 21, 2003; revision accepted Dec. 2, 2003.
For correspondence or reprints contact: Uwe Haberkorn, MD, Department of Nuclear Medicine, University of Heidelberg, Im Neuenheimer Feld 400, Germany-69120 Heidelberg.
E-mail: Uwe_Haberkorn{at}med.uni-heidelberg.de
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Novel Approaches in Anaplastic Thyroid Cancer Therapy
- Bone metastases from differentiated thyroid carcinoma
- Enhancement of sodium/iodide symporter expression in thyroid and breast cancer.
- Radioiodine Therapy of Hepatoma Using Targeted Transfer of the Human Sodium/Iodide Symporter Gene
- The Human Norepinephrine Transporter in Combination with 11C-m-Hydroxyephedrine as a Reporter Gene/Reporter Probe for PET of Gene Therapy