


Abstract
This study compared the possibilities and limitations of 99mTc-labeled synthetic peptides derived from two human antimicrobial peptides, namely, ubiquicidin (UBI) and lactoferrin (hLF), for the scintigraphic detection of bacterial and fungal infections in mice and rabbits. The rationale of our approach was that selected peptides accumulate in infected areas but not in sterile inflammatory lesions, because they bind preferentially to microorganisms. 99mTc-labeled human neutrophil peptides (defensins), ciprofloxacin, and human polyclonal IgG were included as control agents. Methods: 99mTc-labeled peptides and control agents were injected intravenously into animals that had been injected intramuscularly 18 h earlier with multidrug-resistant Staphylococcus aureus, Klebsiella pneumoniae, or fluconazole-resistant Candida albicans. Sterile inflammatory sites were induced by the injection of heat-killed microorganisms or lipopolysaccharide (LPS) into the thigh muscle. Up to 4 h after injection, the accumulation of 99mTc-labeled compounds in the infected/inflamed thigh muscles was determined using scintigraphic techniques and radioactivity counts in dissected tissues. Results: Scintigraphy revealed that 99mTc-labeled peptides UBI 29–41, UBI 18–35, UBI 31–38, hLF 1–11, and defensins, which showed preferential in vitro binding to microorganisms in a former study, accumulated at a significantly higher rate (P < 0.01) in bacterial and C. albicans infections in mice and rabbits than in inflamed tissues induced by heat-killed microorganisms or by LPS. No significant difference in the accumulation of 99mTc-labeled ciprofloxacin was observed between infected and sterile inflamed thigh muscles in mice. Conclusion: 99mTc-labeled antimicrobial peptides UBI 29–41, UBI 18–35, UBI 31–38, hLF 1–11, and defensins accumulate significantly in tissues infected with gram-positive and gram-negative bacteria and C. albicans. Significantly lower (P < 0.01) accumulation of these peptides occurs in sterile inflamed tissues. These data indicate that the peptides preferentially tag microorganisms at the site of infection, which is in agreement with their preferential binding to the microorganisms in vitro and in vivo. 99mTc-labeled ciprofloxacin does not distinguish between infections and sterile inflammatory lesions, which implies that its specificity for the detection of bacterial infections is not warranted.
Various 99mTc-labeled compounds have been developed for the scintigraphic detection of infection and sterile inflammation in humans (1). Unfortunately, these radiopharmaceuticals do not discriminate between infections and sterile inflammatory processes, which is often of clinical importance. In an attempt to develop such a tracer, our research focused initially on radiolabeled human immunoglobulin fractions. Unfortunately, the interaction of these radiolabeled compounds with host cells makes them less optimal for infection detection (2). However, 99mTc-labeled antimicrobial compounds, such as ciprofloxacin (3) and antimicrobial peptides (4,5), may be promising radiopharmaceuticals that distinguish between bacterial infections and sterile inflammatory processes in humans (6). The antibiotic ciprofloxacin and the human neutrophil peptides HNP-1–3 (defensins) have been introduced in nuclear medicine as tracers that preferentially detect bacterial infections. A major drawback of ciprofloxacin is its interaction with both bacterial and mammalian DNA (7). Furthermore, the accumulation of radiolabeled ciprofloxacin at sterile inflammatory sites in laboratory animals was reported recently (8–10). The effects on cells of the immune system, such as chemotactic effects on leukocyte populations and their microbicidal activities (11), may hamper the application of defensins for infection detection (4).
The rationale of our approach was that radiolabeled synthetic peptides derived from antimicrobial domains of human natural peptides that bind preferentially to microorganisms (5) can serve as infection-specific tracers. This article is a sequel to previous studies in this field (4,5). It is a comparative study of the possibilities and limitations of various 99mTc-labeled antimicrobial peptides, ciprofloxacin, and human IgG to detect bacterial or fungal infections in mice and rabbits and their accumulation in sterile inflamed tissues.
MATERIALS AND METHODS
Proteins, Peptides, and Ciprofloxacin
The three ubiquicidin (UBI) peptides described in this study (UBI 29–41, UBI 18–35, and UBI 31–38) were chosen because of their preferential in vitro and in vivo binding to microorganisms over host cells (5). This characteristic was also the rationale for the selection of the three lactoferrin (hLF) peptides (hLF 1–11, hLF 21–31, and hLF 4–11). The peptide hLF 4–11 was chosen as a negative control agent (5). The defensins were chosen as positive control agents for infection detection (5) and were purified from human neutrophils (12). The amino acid sequences of the various peptides are given in Table 1. Human polyclonal IgG was obtained from the Central Laboratory of the Red Cross Blood Transfusion Service (Amsterdam, The Netherlands) and served as a positive control agent for infection and inflammation (13). Stocks of the peptides were stored in 0.01% (vol/vol) acetic acid (pH 4; −20°C). Ciprofloxacin was included in this study because of its suggested specificity for bacteria (3) and was obtained from Bayer AG (Leverkusen, Germany).
Antimicrobial Agents Used in This Study
Labeling Procedure and Quality Control
Antimicrobial peptides and IgG were labeled with 99mTc as described in an earlier article (5), and ciprofloxacin was labeled with 99mTc as described elsewhere (3). The percentage free 99mTc activity in the labeling solutions was determined by instant thin-layer chromatography (ITLC) using saline or methyl ethyl ketone as the eluent and by high-performance liquid chromatography as described earlier (5). Because free 99mTc activity in the solutions containing radiolabeled antimicrobial peptides and IgG (referred to as 99mTc-labeled peptides hereafter) did not exceed 5%, no further purification was performed. Before the application of 99mTc-labeled ciprofloxacin in the experiments, this preparation was subjected to ion-exchange Sephadex-DEAE A-25 chromatography columns (Sigma Chemical Co., St. Louis, MO) to remove free 99mTc activity, which ranged from 20% to 40% of the total activity of the mixture. The percentage free 99mTc activity in the labeling solutions was determined by ITLC using saline or methyl ethyl ketone as the eluent. Finally, all solutions containing 99mTc labeling agents were diluted in phosphate-buffered saline to a concentration of 10 nmol/mL peptide or ciprofloxacin.
Microorganisms
Staphylococcus aureus 25923 and Klebsiella pneumoniae 43816 were obtained from the American Type Culture Collection (Rockville, MD), and the fluconazole-resistant Candida albicans Y01–19 was a gift from Pfizer Inc. (New York, NY). The multidrug-resistant S. aureus type 2141 (MRSA) was a clinical isolate. Overnight cultures of bacteria were prepared in brain–heart infusion broth (Oxoid, Basingstoke, U.K.) in a shaking water bath at 37°C. Overnight cultures of C. albicans were prepared in sabouraud broth (Oxoid), and they were subcultured for 2.5 h on a rotary wheel at 37°C. Virulent bacteria and C. albicans were maintained in mice. Briefly, about 1 × 107 colony-forming units (CFUs) of the microorganisms were injected into the tail vein of each mouse, and 24 h thereafter the mice were killed. The spleen of each mouse was removed aseptically and homogenized, and appropriate dilutions of the homogenate were plated onto diagnostic sensitivity test agar (Oxoid), trypsone soy agar (Oxoid), or sabouraud agar (Oxoid). A single CFU was transferred into 25 mL of the appropriate broth and incubated for 24 h at 37°C. Aliquots of these suspensions containing about 5 × 108 virulent microorganisms per milliliter of broth were stored at −20°C. Additionally, stocks of heat-killed bacteria or C. albicans, boiled for 2 h at 100°C, were prepared and stored at −20°C.
Animals
Specific-pathogen–free male Swiss mice (weight range, 20–25 g) and male New Zealand White rabbits (weight range, 2.5–4 kg) were used in this study. The animals were housed in the animal housing facilities of the Leiden University Medical Center for at least 1 wk before the onset of the experiments. Food and water were given ad libitum. All animal studies were performed in compliance with the local Experimental Animal Ethical Committee and the Dutch laws related to the conduct of animal experiments.
Bacterial and C albicans Infections in Mice and Rabbits
Mice were anesthetized with a single intraperitoneal injection of 0.1 mL saline containing 1 mg fluanisone and 0.03 mg fentanyl citrate (Hypnorm; Janssen Pharmaceutics, Tilburg, The Netherlands). Next, approximately 1 × 107 CFUs of bacteria (S. aureus, MRSA, and K. pneumoniae) or C. albicans in 0.1 mL saline were injected into the right thigh muscle of each mouse. After 24 h, the mice were killed. The infected thigh muscles were dissected and homogenized, and the number of bacteria or C. albicans was determined microbiologically. The number of K. pneumoniae (2 × 106 CFUs/g tissue) after 4 h of infection was significantly less (P < 0.05) than the number of S. aureus (2 × 107 CFUs/g tissue), MRSA (8 × 108 CFUs/g tissue), and C. albicans (5 × 107 CFUs/g tissue). In additional studies, mice were made leukocytopenic by injecting 0.2 mL cyclophosphamide (200 mg/kg) intraperitoneally 3 d before induction of infections with 2 × 105 CFUs of bacteria (S. aureus, MRSA, and K. pneumoniae). After 24 h, the number of microorganisms in the thigh muscles was comparable with the number in immunocompetent mice (0.2–8 × 108 CFUs/g tissue). Blood counts were performed on randomly selected mice to confirm leukocytopenia.
Selected peptides were also applied in rabbits with an experimental infection. In short, the rabbits were injected with 0.4 mL saline containing 1 × 107 CFUs of bacteria (S. aureus, MRSA, and K. pneumoniae) into the right thigh muscle or into the right front leg muscle. The rabbits were anaesthetized 18 h thereafter by a single injection of 0.4 mL saline containing 4 mg fluanisone and 0.13 mg fentanyl citrate into the left thigh muscle.
Inflammatory Processes in Mice and Rabbits
The accumulation of radiolabeled tracers was also studied in animals with a sterile inflammatory process to select compounds with a low accumulation in inflamed tissues that may discriminate between infection and sterile inflammation. Mice and rabbits were anaesthetized as described previously. Sterile inflammation was induced 18 h before administration of the tracer by an intramuscular injection of 0.1 mL saline containing either 1 mg lipopolysaccharide (LPS) or approximately 2 × 108 heat-killed bacteria or C. albicans. Eighteen hours after injection with both stimuli, we observed a significant increase (>60%; P < 0.001) in the weight of the injected thigh muscle compared with the contralateral thigh muscle.
Scintigraphy
Animals were anaesthetized 18 h after infection as described previously, and 0.1 mL labeling solution containing 1 nmol peptide or anti-infective was injected into a tail vein. The accumulation of the radiolabeled peptides, ciprofloxacin, and IgG in the bacterial or C. albicans infected muscles in mice and rabbits was assessed by planar scintigraphy. Before scintigraphy, a subcutaneous injection of 0.05 mg diazepam (Valium; Hoffmann-Roche, Mijdrecht, The Netherlands) in 0.1 mL saline was administered to the mice to induce muscle relaxation. Next, the animals were placed in a supine position on a collimator of a planar gamma camera with both hind legs spread out and fixed with surgical tape. Continuous anterior whole-body acquisitions of the animals every 60 s during the first 2 h after injection and a single static acquisition at 4 h after injection of the tracer were made. On the scintigrams, anatomically adjusted regions of interest were drawn over the entire infected or inflamed muscle (target) and the contralateral muscle (nontarget). The accumulation of 99mTc-labeled tracers at sites of infection or inflammation is expressed as the ratio of the counts in the target muscle to the counts in the nontarget muscle (T/NT). Scintigraphic data were interpreted by two observers, and the results were always similar to the data obtained after obduction, as described later. When scintigraphic data from the two sessions differed, we used T/NT obtained after obduction. For comparison of the accumulation of the various 99mTc-labeled compounds, we calculated the mean ± SEM of T/NT at 30, 60, and 120 min after injection of the tracers into the mice with infected or inflamed thigh muscles. The animals were killed 4 h after tracer injection by an intraperitoneal injection of 0.5 mL (mice) or 5 mL (rabbits) sodium barbiturate (60 mg/mL saline, Nembutal; Sanofi BV, Division Algin, Maassluis, The Netherlands). Next, the entire infected and noninfected thigh muscles were removed, weighed, and counted for radioactivity, and T/NT was calculated.
Statistical Analysis
Differences in the data were evaluated with the Student t test. Results for the probability value using the 2-tailed test are reported, and all results are given as mean ± SEM.
RESULTS
Detection of Bacterial Infections in Mice
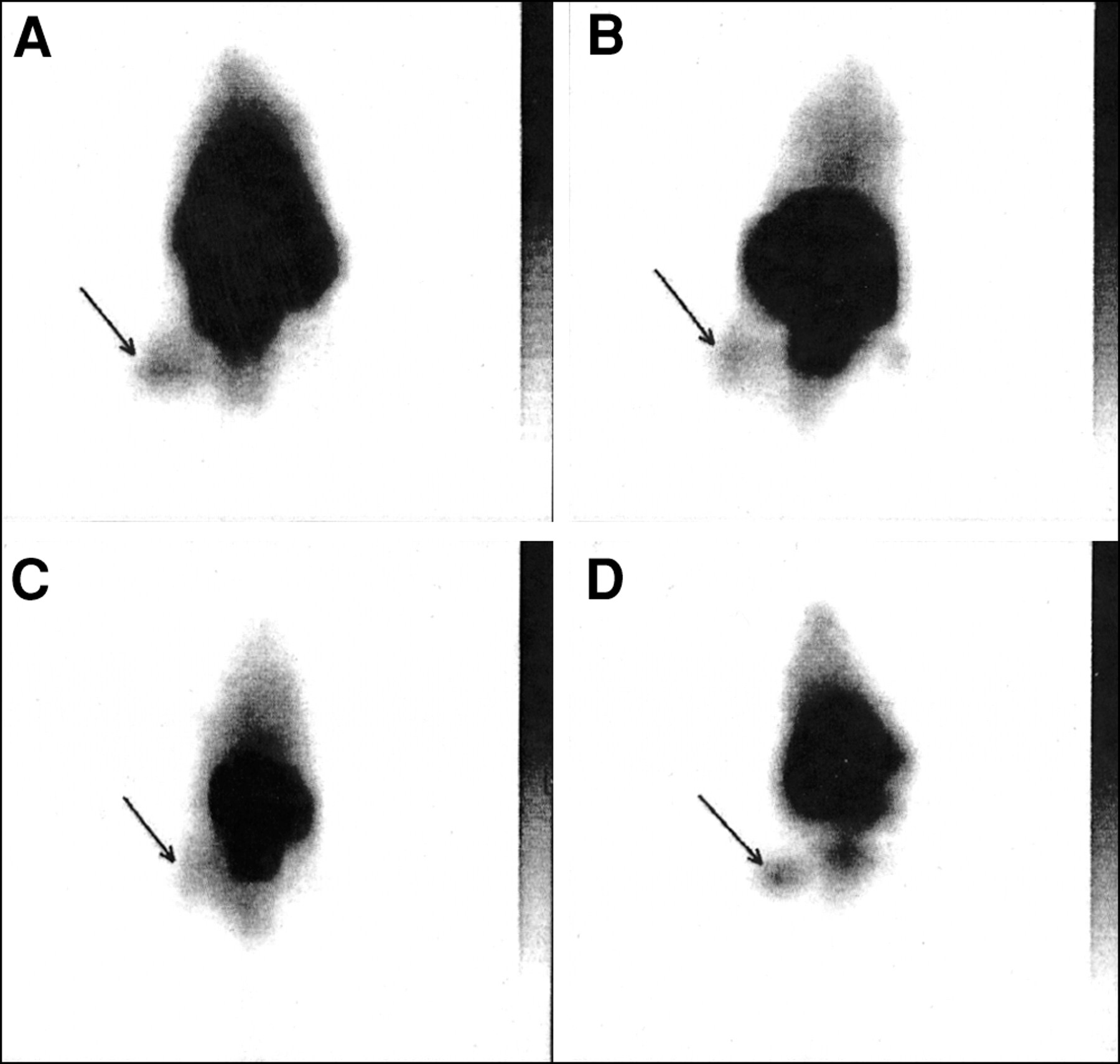
The sites of bacterial infections could be visualized within 1 h after injection of the 99mTc-labeled peptides. Typical scintigrams for UBI 29–41 and hLF 1–11 in S. aureus infected mice are depicted in Figure 1. The accumulation of 99mTc-labeled UBI 29–41 in the thigh muscle of mice infected with various microorganisms is shown in Figure 2. Within the period of analysis, significantly higher T/NT was observed for 99mTc-labeled UBI peptides as well as for hLF 1–11, hLF 21–31, and the defensins in infected thigh muscles compared with T/NT measured in sterile inflammatory sites (Table 2). The accumulation of ciprofloxacin (Fig. 1) and 99mTc-labeled hLF 4–11 in infected tissues was comparable and showed no significant difference between bacterial infections and sterile inflammatory sites. In a small group of randomly selected animals, we determined T/NT also from data obtained with entirely dissected thigh muscles. The comparison of these data revealed similar T/NT after calculations from scintigraphic data (results not shown).
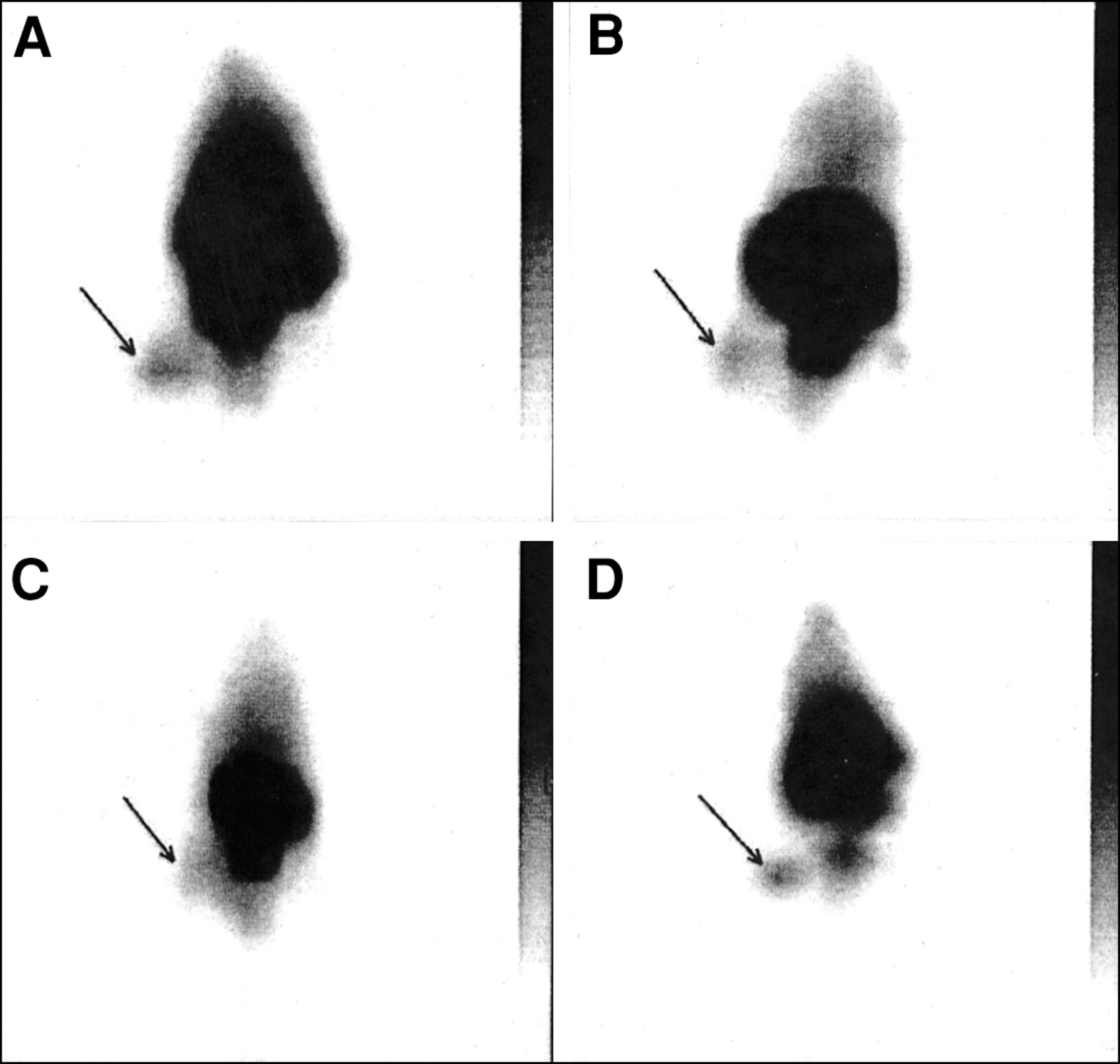
Typical scintigrams of 99mTc-labeled UBI 29–41 (A), hLF 1–11 (B), ciprofloxacin (C), and human polyclonal IgG (D) 1 h after injection into mice having thigh muscles infected with S. aureus.
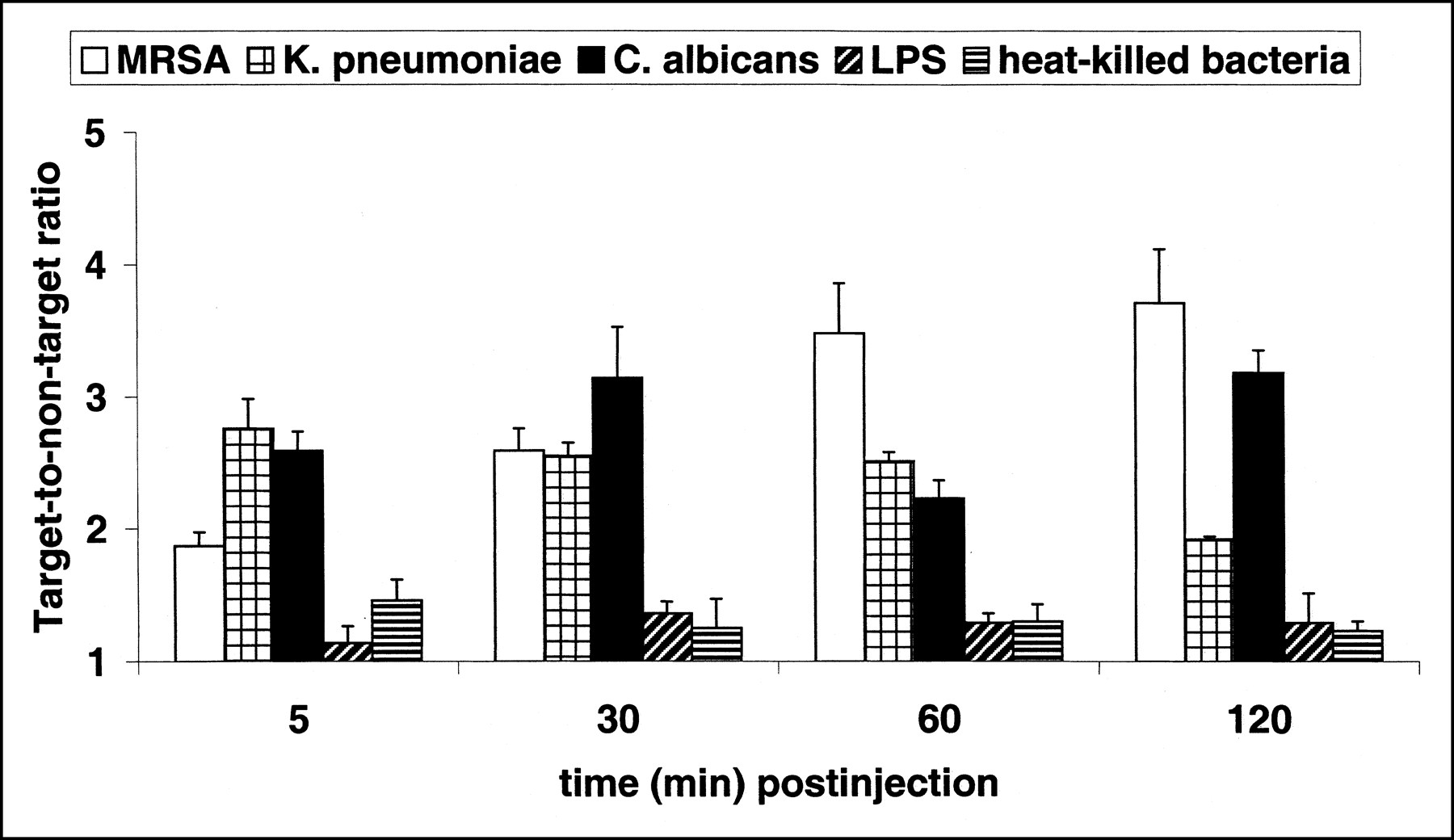
Accumulation of 99mTc-labeled antimicrobial peptide UBI 29–41 in thigh muscles of mice infected with MRSA (white bars), K. pneumoniae (square-hatched bars), or fluconazole-resistant C. albicans (black bars). Furthermore, mice were injected intramuscularly with LPS (diagonally hatched bars) or heat-killed bacteria (horizontally hatched bars) 18 h before administration of tracer. Each symbol at each time point represents mean ± SEM T/NT of at least 12 animals obtained from three independent experimental settings.
Mean T/NT of Infected/Inflamed Thigh Muscles After Injection of 99mTc-Labeled Compounds
To investigate whether the accumulation of the tracers depends on the binding to infiltrating leukocytes, we determined T/NT for the various 99mTc-labeled UBI peptides in leukocytopenic mice with an MRSA or K. pneumoniae infection. These data were compared with the respective values of accumulation in immunocompetent mice. The results revealed no significant differences in intensity and rate of accumulation of the 99mTc-labeled peptides in the infected tissues of leukocytopenic and immunocompetent mice. This finding indicated that the accumulation of radiolabeled UBI peptides is not dependent on the interaction of these tracers with infiltrating leukocytes.
Detection of Infection with C. albicans in Mice
Immunocompetent mice were used to study the accumulation of 99mTc-labeled peptides, ciprofloxacin, and IgG in experimental thigh muscle infections with C. albicans. On the scintigrams, the C. albicans infections in the thigh muscles of mice could be seen within 1 h after injection of 99mTc-labeled compounds. For example, the time-dependent accumulation of various tracers in the thigh muscles of mice infected with viable C. albicans is shown in Figure 3. At 30 min after injection, all tracers visualized the infected thigh muscles, and the mean T/NT over the interval from 30 to 120 min after injection was high for all tracers tested (Table 2).
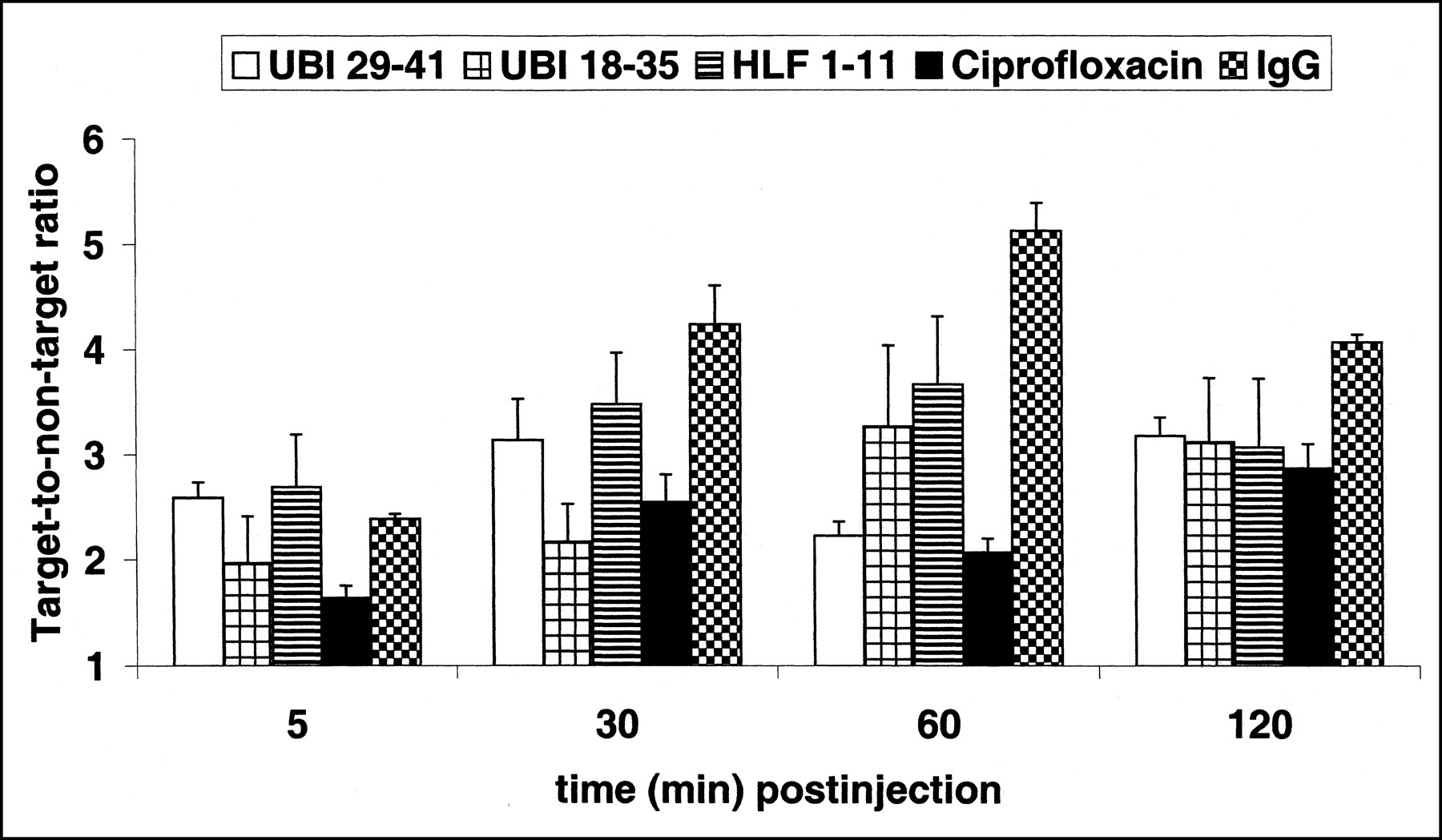
Accumulation of 99mTc-labeled UBI 29–41 (white bars), UBI 18–35 (square-hatched bars), hLF 1–11 (horizontally hatched bars), ciprofloxacin (black bars), and IgG (checkered bars) in thigh muscles of mice infected 18 h earlier with fluconazole-resistant C. albicans. Each symbol at each time point represents mean ± SEM T/NT of at least six animals obtained from three independent experimental settings.
Sterile Inflammations in Mice
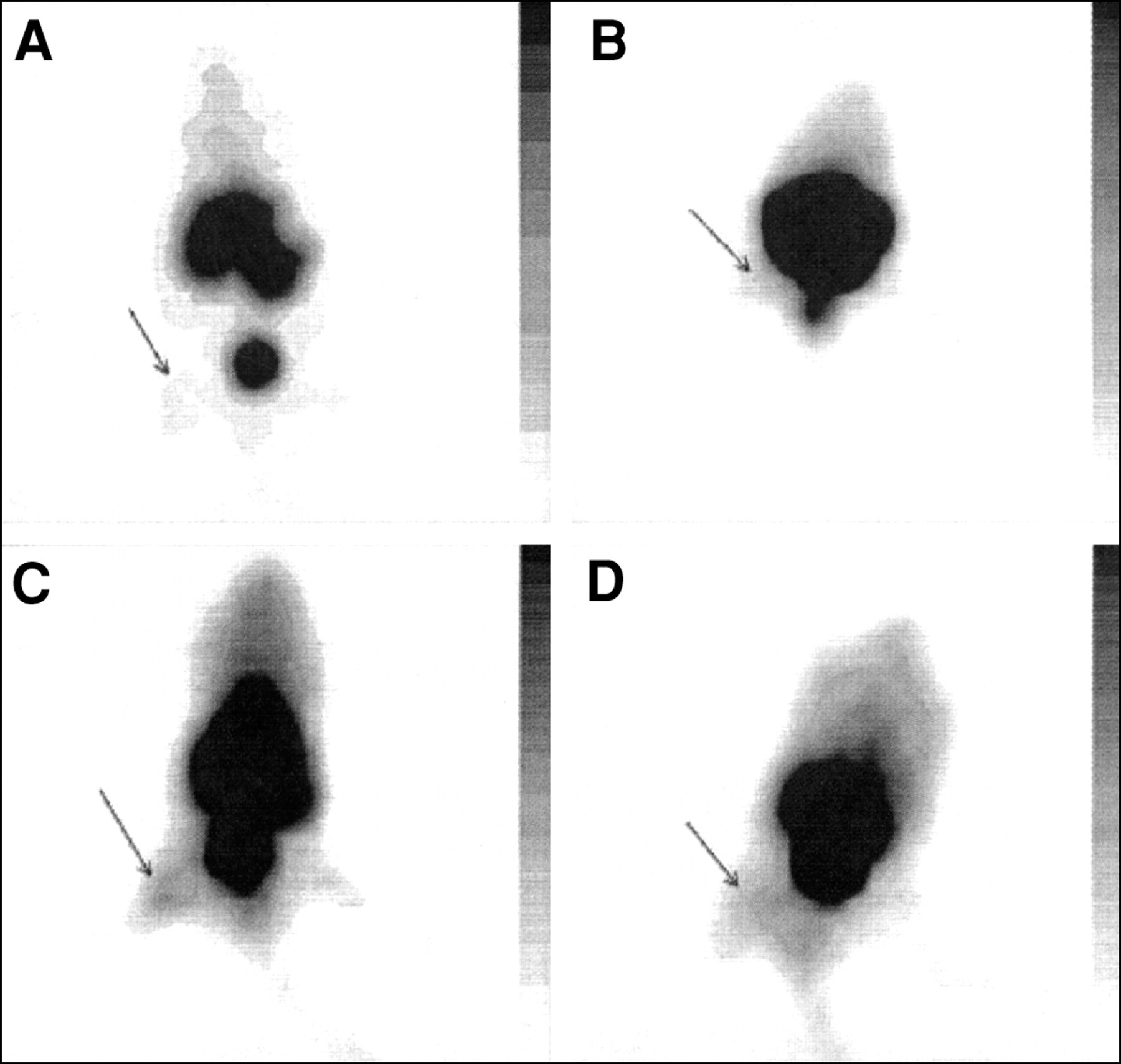
The accumulation of radiolabeled tracers was also studied in animals with sterile inflammatory processes to select compounds that discriminate between infection and sterile inflammation. The results for typical scintigrams of various radiolabeled tracers in mice with thigh muscles injected with heat-killed S. aureus are shown in Figure 4. Good accumulation in mice with sterile inflammatory processes (induced by injections with LPS or heat-killed microorganisms) was observed for 99mTc-labeled ciprofloxacin and IgG (Fig. 4).
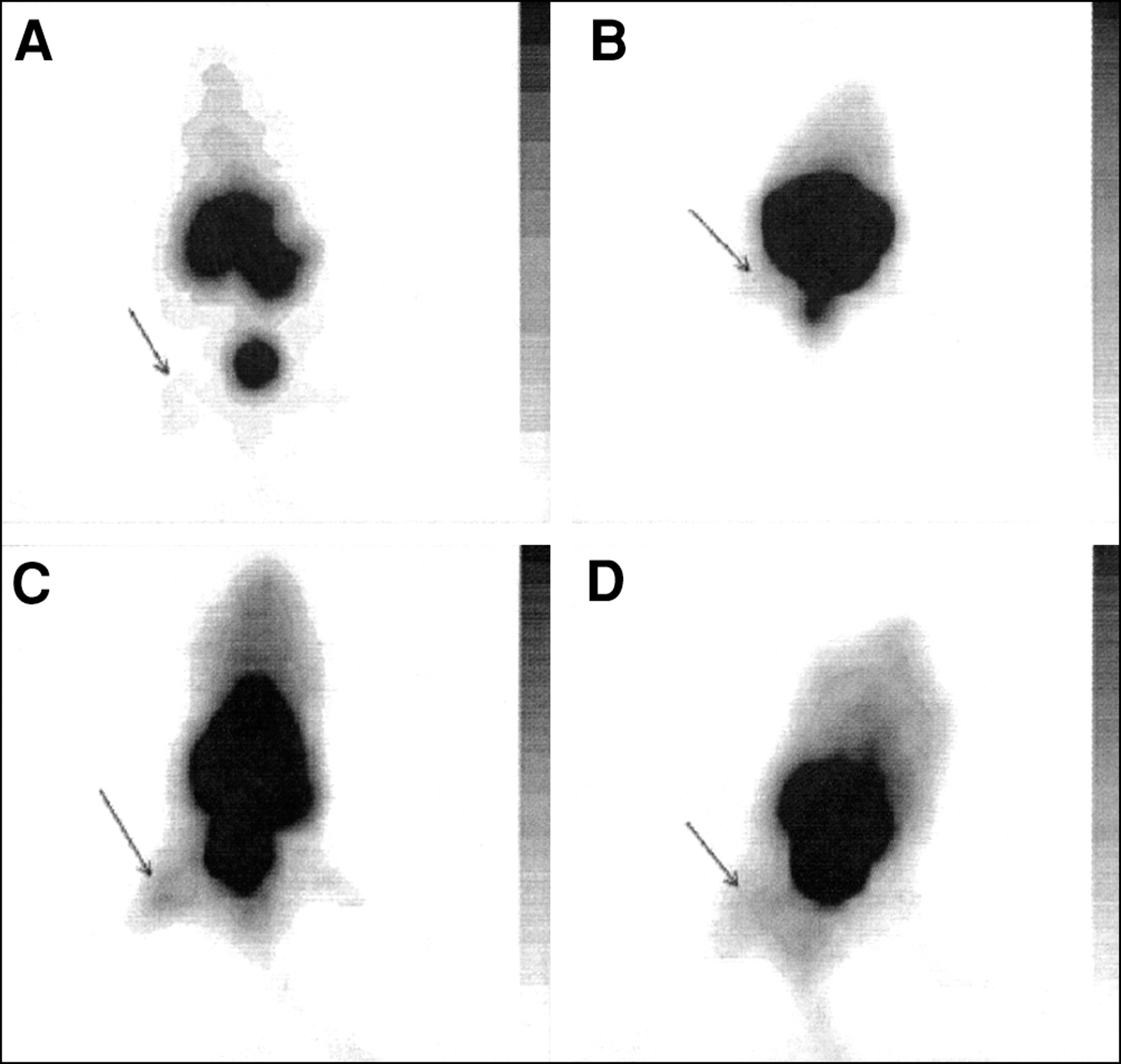
Typical scintigrams of 99mTc-labeled UBI 29–41 (A), hLF 1–11 (B), ciprofloxacin (C), and human polyclonal IgG (D) 1 h after injection into mice having thigh muscles injected with heat-killed S. aureus.
Bacterial Infections in Rabbits
Because of the favorable results in mice, we assessed the accumulation of the 99mTc-labeled peptides UBI 29–41, UBI 18–35, and UBI 22–35 in rabbits with bacterial infections of the thigh muscle or front leg muscle. Figure 5A is a scintigram of a rabbit with a K. pneumoniae infection in the thigh muscle 1 h after injection of 99mTc-labeled UBI 29–41. Figure 5B is the same scintigram with lower contrast to depict the biodistribution of this peptide 1 h after injection. The highest T/NT in rabbits was observed 4 h after injection of 99mTc-labeled peptides; for example, for UBI 29–41 (Fig. 6), the highest T/NT was approximately 6. Two rabbits with a K. pneumoniae infection in the front leg were injected with 99mTc-IgG, and the infected area became visible 2 h after injection. The highest values of T/NT (4–5) of 99mTc-IgG in infected tissues were observed 4 h after injection (data not shown).
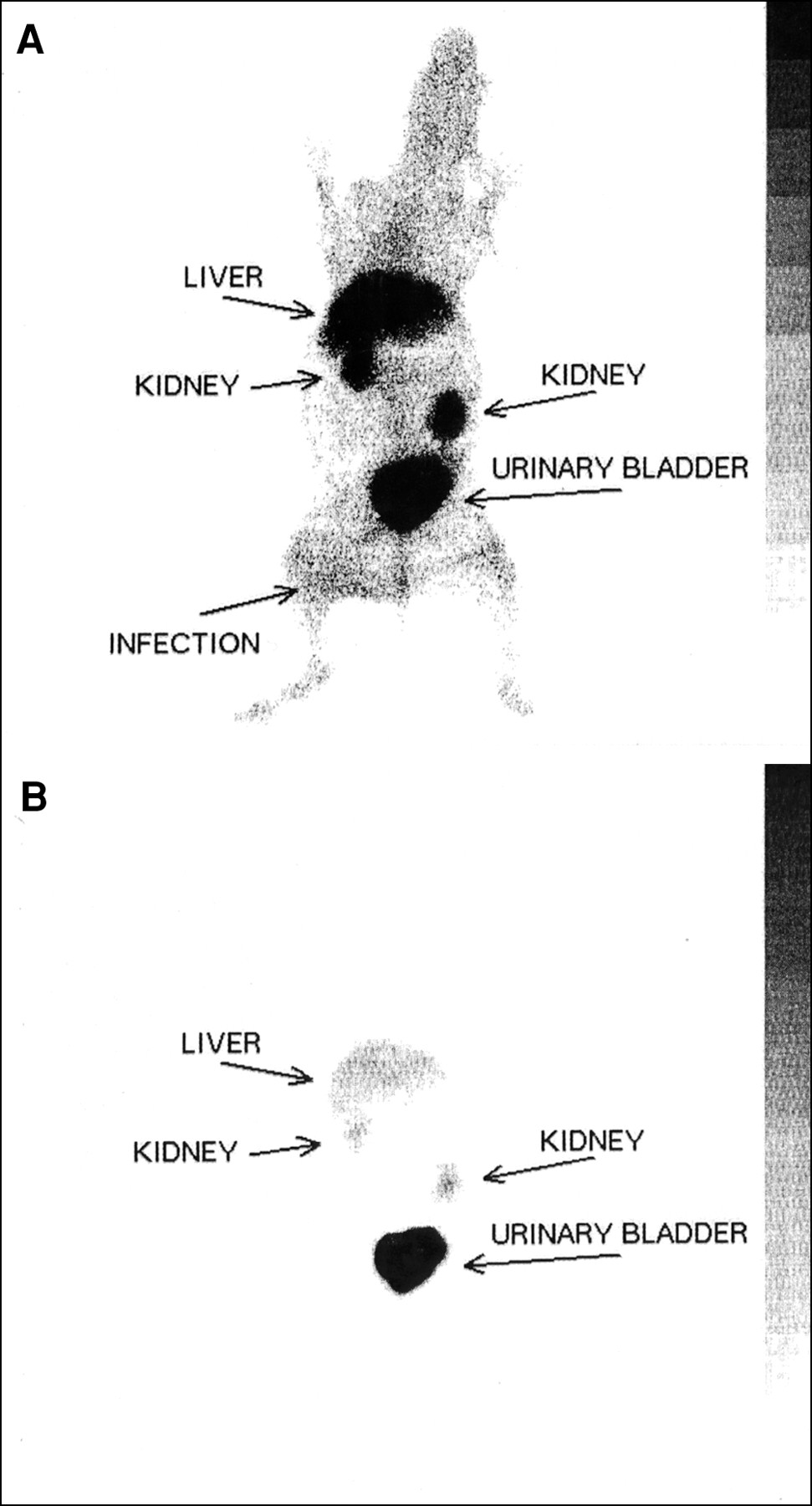
(A) Typical scintigram of 99mTc-labeled UBI 29–41 1 h after injection into rabbits with thigh muscles infected with S. aureus. (B) Biodistribution of 99mTc-labeled UBI 29–41 1 h after injection with lower contrast.
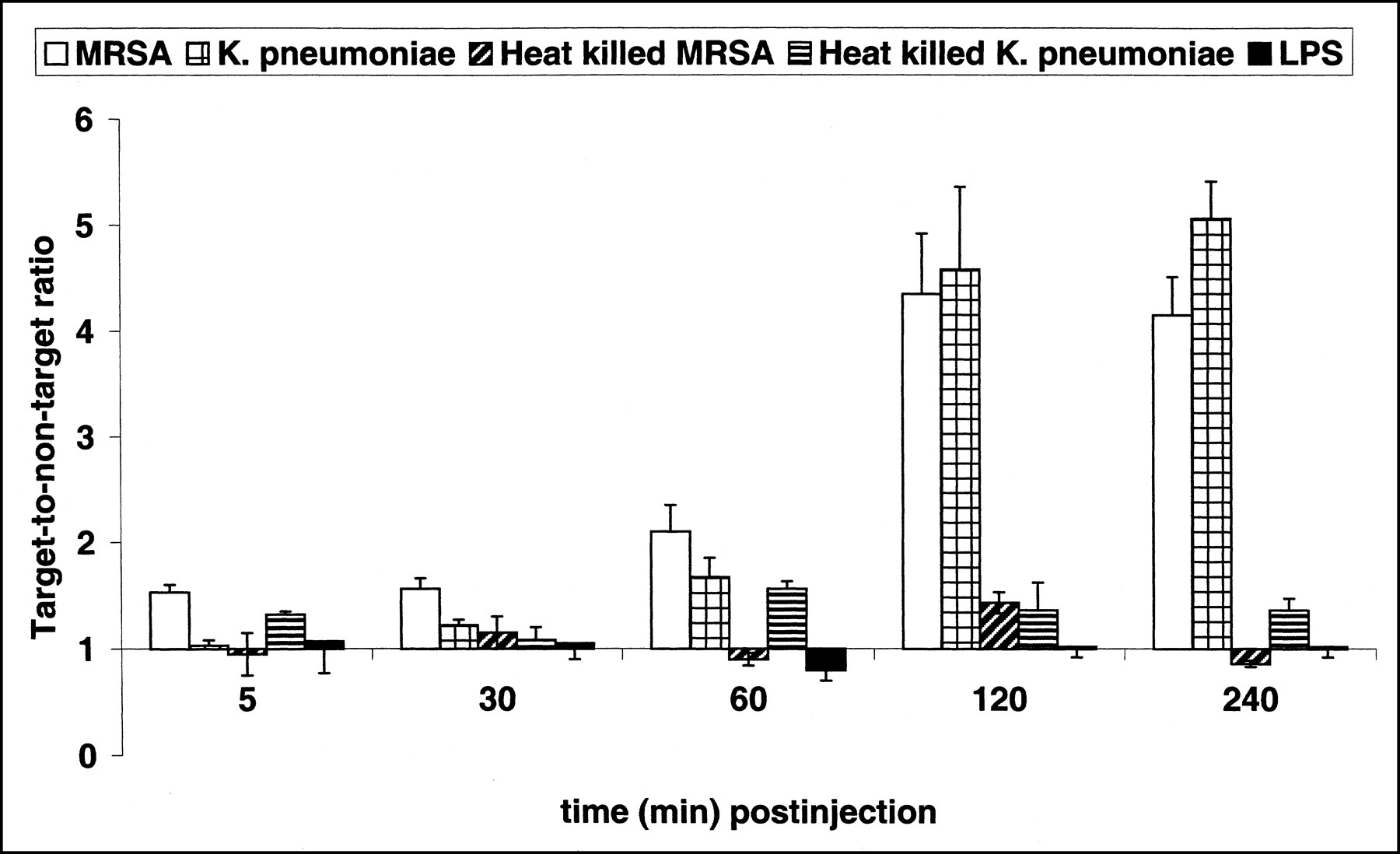
Accumulation of 99mTc-labeled UBI 29–41 in thigh muscles of rabbits injected with MRSA (white bars), K. pneumoniae (square-hatched bars), heat-killed MRSA (diagonally hatched bars), heat-killed K. pneumoniae (horizontally hatched bars), or 50 mg LPS (black bars). Each symbol at each time point represents mean ± SEM T/NT of at least three animals obtained from independent experimental settings.
Sterile Inflammations in Rabbits
As observed in mice, the 99mTc-labeled UBI peptides did not visualize inflamed thigh muscles or front leg muscles in rabbits (Fig. 6).
DISCUSSION
This investigation focused on the possibilities and limitations of various 99mTc-labeled antimicrobial compounds, including antimicrobial peptides and ciprofloxacin, to detect bacterial or C. albicans infections but not sterile inflammatory processes (induced by LPS or heat-killed microorganisms) in mice and rabbits. Some 99mTc-labeled synthetic peptides derived from human UBI and hLF, as well as natural defensins, can be used to detect infections with bacteria and C. albicans, discriminating these from sterile inflammation. This conclusion is based on the evidence discussed in this section.
99mTc-labeled UBI and hLF peptides and defensins, which all have shown preferential binding to microorganisms in vitro (5), rapidly detected infections in mice and rabbits with significantly higher scintigraphic intensity (P < 0.01) than that for sterile inflammatory sites. Earlier, we reported chemotactic and bactericidal activities of low doses of defensins injected into mice, and these activities make defensins less favorable for the detection of infections, for example, in humans (11). In addition, with 99mTc-labeled hLF peptides, significantly higher accumulation (P < 0.01) was observed in both bacterial and fungal infections than in sterile inflamed tissues. However, their accumulation in liver and deposits in intestines (5) make 99mTc-labeled hLF peptides less favorable for imaging infections. On the basis of these considerations, we prefer 99mTc-labeled UBI peptides, because they can be prepared synthetically under good manufacturing conditions in large amounts, and no adverse effects have been found under the conditions of our experiments.
Another conclusion is that 99mTc-labeled ciprofloxacin accumulates with the same intensity (except T/NT in C. albicans infections was significantly higher) in both infected and sterile inflamed tissues, as was seen with 99mTc-labeled IgG. This finding is in agreement with the recently reported observation that radiolabeled ciprofloxacin accumulates in inflamed tissues in rabbits (8,9). Moreover, we have reported (10), as have others (14,15), on the binding of ciprofloxacin to bacterial and mammalian DNA, DNA-gyrases, and the eukaryotic counterpart topoisomerase II. In agreement, we observed good binding of 99mTc-labeled ciprofloxacin to DNA purified from bacteria (S. aureus and K. pneumoniae) or human leukocytes, as well as to intact microorganisms and (activated) human leukocytes and endothelial cells (10). Another disadvantage of using a low dose of ciprofloxacin in clinical studies could be the emergence of resistant bacteria (16–19). Also, ciprofloxacin affects the immune system, for example, the upregulation of interleukin-2 gene expression in human cells (20).
All currently available tracers for the detection of infections are based on nonspecific entrapment by binding to leukocytes present at the site of infection or inflammation. It is unlikely that the binding of 99mTc-labeled UBI peptides and defensins to leukocytes contributes significantly to the accumulation of these peptides in infected tissues, because these 99mTc-labeled compounds did not accumulate in sterile inflamed tissues in mice and rabbits. Furthermore, 99mTc-labeled UBI peptides and defensins bound preferentially to bacteria over (activated) human leukocytes (5) in vitro and in vivo. In addition, the accumulation of 99mTc-labeled UBI peptides in infected thigh muscles of leukocytopenic animals is not different from that in bacterial infected thigh muscles in immunocompetent mice. Because the inoculum used to infect leukocytopenic mice contained a much lower number of pathogens than that used in immunocompetent mice, we were able to achieve a comparable number of microorganisms in immunocompetent mice. These studies were performed to exclude the possibility that host cells, including infiltrating leukocytes, contribute significantly to the accumulation of 99mTc-labeled UBI peptides at the site of infection. Recently, a good correlation was also observed for two 99mTc-labeled UBI peptides between the number of microorganisms and the accumulation of the peptides (unpublished results).
CONCLUSION
Taken together, 99mTc-labeled antimicrobial peptides, such as UBI 29–41, UBI 18–35, and UBI 31–38, allow rapid and intense visualization of foci of bacterial or C. albicans infections in mice and rabbits. These peptides show significantly lower accumulation (P < 0.01) in sterile inflammatory processes. On the basis of data from in vitro binding studies (5), we conclude that these peptides bind preferentially to microorganisms compared with host (inflammatory) cells at the site of infection. Furthermore, it is likely that 99mTc-labeled ciprofloxacin does not distinguish significantly between infections and inflammatory lesions. Of course, an extensive study of the possible toxicologic effects of synthetic UBI peptides in laboratory animals must be performed before these peptides can be considered for studies in humans. It is encouraging that no toxicologic or immunobiologic side effects of the UBI peptides were observed in our experiments.
Acknowledgments
The authors thank Graciela Rodriguez, Bep Ravensbergen, Maria van den Barselaar, Bram Sinon, and Petra Dibbets-Schneider for their technical support and Dr. Wim Calame for his critical reading of the manuscript.
Footnotes
Received Aug. 30, 2000; revision accepted Dec. 18, 2000.
For correspondence or reprints contact: Ernest K.J. Pauwels, DSc, Department of Radiology, Division of Nuclear Medicine, Leiden University Medical Center, C4-Q, P.O. Box 9600, NL-2300 RC Leiden, The Netherlands.
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- 68Ga-NOTA-UBI-29-41 as a PET Tracer for Detection of Bacterial Infection
- Improved Bacterial Detection Using Immobilized Acyl-Lysyl Oligomers
- Comparison of 99mTc- and 18F-Ubiquicidin Autoradiography to Anti-Staphylococcus aureus Immunofluorescence in Rat Muscle Abscesses
- Leukocyte Receptor-Binding Radiopharmaceuticals for Infection and Inflammation Scintigraphy
- Antimicrobial Peptide 99mTc-Ubiquicidin 29-41 as Human Infection-Imaging Agent: Clinical Trial
- 99mTc-Labeled Antimicrobial Peptide Ubiquicidin (29-41) Accumulates Less in Escherichia coli Infection than in Staphlococcus aureus Infection
- 99mTc-Labeled UBI 29-41 Peptide for Monitoring the Efficacy of Antibacterial Agents in Mice Infected with Staphylococcus aureus
- Specificity of 99mTc-Ciprofloxacin Imaging
- Inability of 99mTc-Ciprofloxacin Scintigraphy to Discriminate Between Septic and Sterile Osteoarticular Diseases
- Mechanisms of Antimicrobial Peptide Action and Resistance
- 99mTc-Labeled Antimicrobial Peptides for Detection of Bacterial and Candida albicans Infections