


Abstract
The successful peptide receptor imaging of tumors, as exemplified for somatostatin receptors, is based on the overexpression of peptide receptors in selected tumors and the high-affinity binding to these tumors of agonist radioligands that are subsequently internalized into the tumor cells in which they accumulate. Although in vitro studies have shown ample evidence that the ligand–receptor complex is internalized, in vivo evidence of agonist-induced internalization of peptide receptors, such as somatostatin receptors, is missing. Methods: Rats subcutaneously transplanted with the somatostatin receptor subtype 2 (sst2)–expressing AR42J tumor cells were treated with intravenous injections of various doses of the sst2 agonist [Tyr3, Thr8]-octreotide (TATE) or of the sst2 antagonist 1,4,7,10-tetraazacyclododecane-N,N′,N″,N′″,-tetraacetic acid (DOTA)-Bass and were sacrificed at various times ranging from 2.5 min to 24 h after injection. The tumors and pancreas were then removed from each animal. All tissue samples were processed for sst2 immunohistochemistry using sst2-specific antibodies. Results: Compared with the sst2 receptors in untreated animals, which localized at the plasma membrane in pancreatic and AR42J tumor cells, the sst2 receptors in treated animals are detected intracellularly after an intravenous injection of the agonist TATE. Internalization is fast, as the receptors are already internalizing 2.5 min after TATE injection. The process is extremely efficient, as most of the cell surface receptors internalize into the cell and are found in endosomelike structures after TATE injection. The internalization is most likely reversible, because 24 h after injection the receptors are again found at the cell surface. The process is also agonist-dependent, because internalization is seen with high-affinity sst2 agonists but not with high-affinity sst2 antagonists. The same internalization properties are seen in pancreatic and AR42J tumor cells. They can further be confirmed in vitro in human embryonic kidney–sst2 cells, with an immunofluorescence microscopy–based sst2 internalization assay. Conclusion: These animal data strongly indicate that the process of in vivo sst2 internalization after agonist stimulation is fast, extremely efficient, and fully functional under in vivo conditions in neoplastic and physiologic sst2 target tissues. This molecular process is, therefore, likely to be responsible for the high and long-lasting uptake of sst2 radioligands seen in vivo in sst2-expressing tumors.
Peptide receptor targeting of cancer is gaining increasing interest in the field of nuclear medicine. The molecular basis for this tumor imaging method is the expression of specific peptide receptors in many cancers (1). One of the cellular mechanisms considered the basis of successful peptide receptor targeting in vivo is the agonist-induced receptor internalization in these tumor cells (2). The accumulation of internalized radioligands into tumor cells through this mechanism may be crucial not only for the quality of tumor imaging but also for the therapeutic efficacy of radioligand application (1). The best examples of peptide receptors representing highly promising targets for in vivo cancer targeting are primarily somatostatin receptors but also gastrin-releasing peptide receptors, cholecystokinin receptors, and glucagon-like peptide 1 receptors (1,3–5).
Although the somatostatin receptors, in particular the sst2 subtype, are well established and most successful among the peptide receptor–targeting candidates (5–7), information on sst2 internalization is limited to in vitro systems. Somatostatin radiotracer uptake is widely measured in various cell culture systems (8–11) and can be used to test new somatostatin candidates. Alternatively, cell cultures can be used to determine the agonist-induced internalization of sst2 receptors monitored by immunofluorescence microscopy as a complementary test (12). Because of technical difficulties, there are no data, however, on in vivo agonist-induced receptor internalization in tumors.
The aim of this study was to evaluate whether the internalization of receptors after agonist stimulation can be observed in vivo in cancers and, if so, to determine what characteristics internalization had. We chose the sst2 subtype of somatostatin receptors for the evaluation of internalization. Indeed, somatostatin receptors have been found to be excellent targets in most neuroendocrine tumors; moreover, the most frequently expressed receptor among the 5 somatostatin receptor subtypes in these tumors is sst2. It is well established that the AR42J cell line expresses sst2 abundantly (8,13) and can be implanted subcutaneously into rats. With this in vivo tumor model, we have, therefore, evaluated the ability of the somatostatin agonist [Tyr3, Thr8]-octreotide (TATE) to trigger sst2 internalization under in vivo conditions at various times and at various concentrations. In addition, in the same animals, a physiologic somatostatin target organ—the pancreas (also known to express sst2 (14))—was evaluated under the same conditions. We investigated morphologically the trafficking behavior of the sst2 receptor from the plasma membrane into the cells under these conditions, in tumoral and in normal tissues. Immunohistochemical methods were used for this purpose, with specific and well-established sst2 antibodies (R2-88, SS-800, and UMB-1) (15,16). Moreover, we compared the results with a well-established method monitoring sst2 internalization in vitro: an immunofluorescence microscopy–based internalization assay (12).
MATERIALS AND METHODS
AR42J Animal Tumor Model
Animals were kept, treated, and cared for in compliance with the guidelines of the Swiss regulations (approval 789). A total of 10–12 million AR42J cells, freshly suspended in sterile phosphate-buffered saline (PBS), were subcutaneously implanted in one flank of Lewis rats (49–55 g). After inoculation (11–15 d), the rats weighing 120–150 g showed solid palpable tumor masses (tumor weight, 70–150 mg) and were used for the in vivo internalization experiments (8). Under isoflurane anesthesia, a first set of rats (2 per group) was injected with 0.21 mg of the sst2 agonist TATE in 0.2 mL of sodium chloride solution (0.9%; 0.1% bovine serum albumin) into the lateral tail vein; this set of rats was sacrificed at 2.5 min, 10 min, 20 min, 1 h, 6 h, and 24 h after injection. A second set of rats (2 per group) was injected with 0.21 mg, 21 μg, 2.1 μg, or 0.21 μg of TATE and sacrificed at 1 h after injection. A third group of animals (2 per group) was injected with 0.21 mg of the sst2 antagonist 1,4,7,10-tetraazacyclododecane-N,N′,N″,N′″,-tetraacetic acid (DOTA)-Bass (17) and sacrificed at 1, 6, or 24 h after injection. Untreated rats injected with only PBS were used as controls. The tumors and pancreas of each animal were collected. All samples were cut in half. One half of the samples was immersed in a 4% formalin solution for 24 h and paraffin-embedded for immunohistochemical investigations. The other half was frozen in dry ice and stored at −80°C for further in vitro receptor autoradiography.
Immunohistochemistry of sst2 Receptors
Immunohistochemistry was performed as described before (15,18–20). All samples were tested with the sst2-specific antibody R2-88 (generously provided by Dr. Agi Schonbrunn) and, to obtain duplicate confirmatory data, also with the commercially available polyclonal sst2-specific antibody SS-800 (Gramsch Laboratories) and with the monoclonal antibody UMB-1 (SS-8000RM; Biotrend GmbH) (15,16). Formalin-fixed, paraffin-embedded tissue sections (4 μm thick) were used. As reported previously (18), the best antigen-retrieval method for R2-88 immunohistochemistry was boiling in the microwave in 5% urea buffer (pH 9.5). It was also the best method for UMB-1 immunohistochemistry. For SS-800, boiling in the microwave in 10 mM citrate buffer (pH 6.0) was preferable (15). Both primary antibodies were applied in a 1:1,000 dilution. The secondary antibody was a biotinylated goat antirabbit immunoglobulin. Antibody binding was visualized using the VECTASTAIN Elite ABC Kit (Vector). Staining was performed with diaminobenzidine and counterstaining with hemalum. For negative control, the primary antibodies were preabsorbed with 100 nM corresponding antigen peptide. In all experiments, a well-characterized gastroenteropancreatic neuroendocrine tumor strongly expressing sst2, as determined by receptor autoradiography, was included as a positive control.
Receptor Autoradiography
The somatostatin receptor autoradiography was performed as described before (21). Briefly, 20-μm-thick frozen tissue sections were cut from each of the collected AR42J tumor and pancreas samples and were incubated for 2 h at room temperature with the universal somatostatin radioligand 125I-[Leu8, d-Trp22, Tyr25]-somatostatin-28 (125I-[LTT]-SS-28). The peptide [LTT]-SS-28 was provided by Dr. Jean Rivier and was iodinated enzymatically using the lactoperoxidase method (Anawa), at a specific activity of 74,000 GBq/mmol (2,000 Ci/mmol). Displacement experiments were performed using sst1-, sst2-, sst3-, sst4-, and sst5-selective analogs as described previously (21). Nonspecific binding was determined in serial tissue sections incubated with the radioligand in 1 μM somatostatin. The slides were exposed to Biomax MR film (Kodak) for 7 d at 4°C.
In Vitro sst2 Internalization Assay Based on Immunofluorescence Microscopy
The human embryonic kidney 293 (HEK293) cell line expressing the T7-epitope–tagged human sst2 (HEK-sst2) receptor (kindly provided by Dr. Stephan Schulz) was cultured at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, 100 units of penicillin per milliliter, 100 μg of streptomycin per milliliter, and 500 μg of G418 per milliliter (Gibco). HEK-sst2 cells were grown on 35-mm 4-well plates (Cellstar; Greiner Bio-One GmbH) coated with poly-d-lysine (20 μg/mL) (Sigma-Aldrich). Cells were treated either with vehicle alone or with agonist at 37°C and 5% CO2 in growth medium for the times indicated. Additionally, in parallel experiments, cells treated with 1 μM agonist for 30 min at 37°C and 5% CO2 were subsequently washed with PBS and incubated for 30 min in an agonist-free medium at 37°C and 5% CO2. Then the cells were processed for immunofluorescence microscopy as described previously (12,22) using the sst2A-specific antibody R2-88 as primary antibody (18,23) and Alexa Fluor 488 goat antirabbit IgG (H+L) (Molecular Probes, Inc.) as secondary antibody. The cells were imaged using a DM RB immunofluorescence microscope (Leica) and a DP10 camera (Olympus).
RESULTS
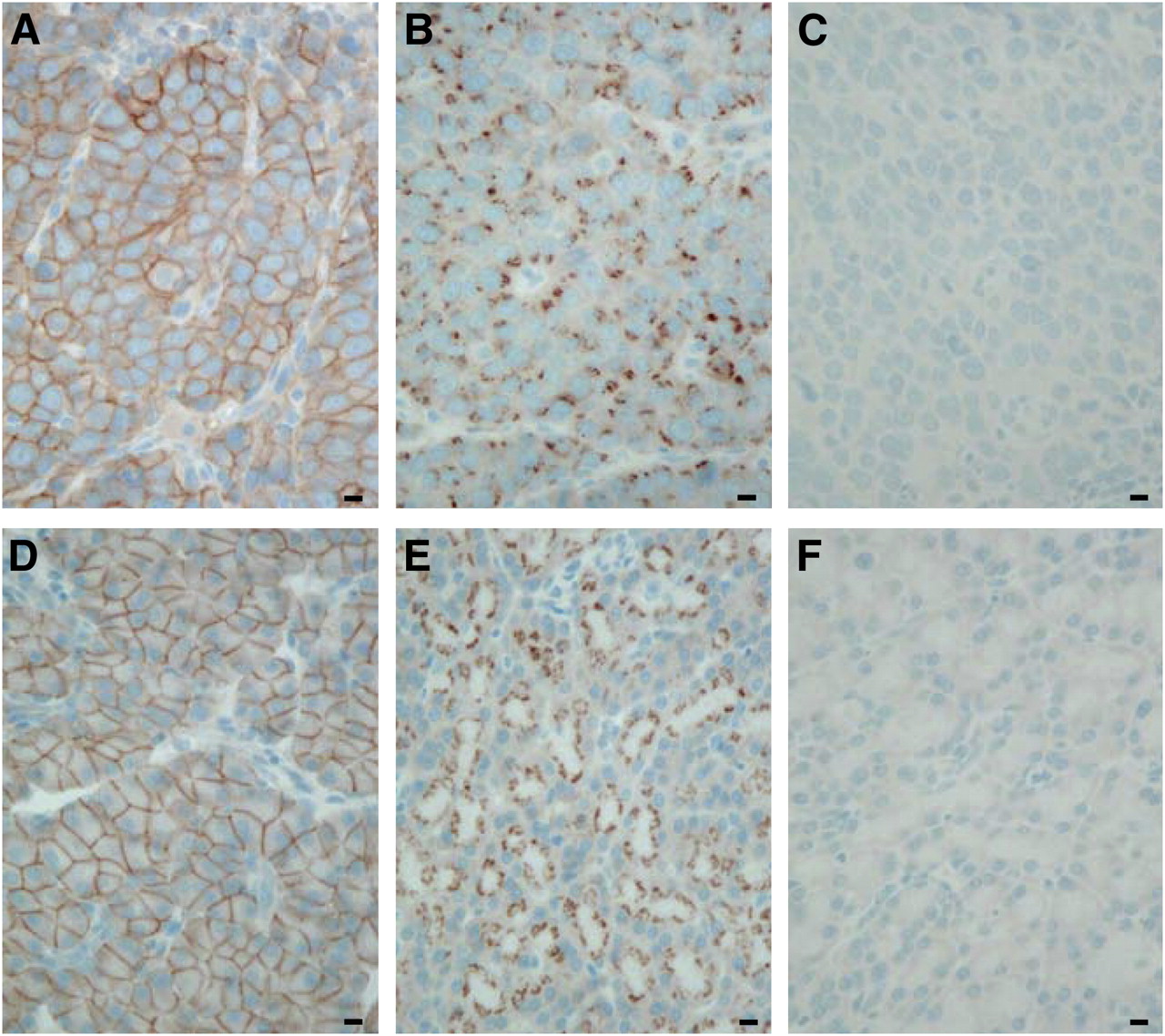
In untreated animals, sst2 receptors are localized at the plasma membrane both in AR42J tumor cells (Fig. 1A) and in the pancreas (Fig. 1D), as determined by immunohistochemistry using the R2-88 antibody. R2-88-stained sst2 receptors are homogeneously distributed in both tissues, with all cells stained predominantly at the plasma membrane. A drastic change in the immunohistochemical pattern of the sst2 cellular localization is observed, however, after an intravenous application of TATE (0.21 mg) in a AR42J tumor–bearing rat that was sacrificed at 1 h after injection. The sst2 receptors are no longer localized on the cell surface at 1 h after agonist treatment but have moved inside the cells (Figs. 1B and 1E), demonstrated both in the AR42J tumor cells (Fig. 1B) and in the pancreatic acini (Fig. 1E). No staining is seen after preabsorption with the antigen peptide (Figs. 1C and 1F).
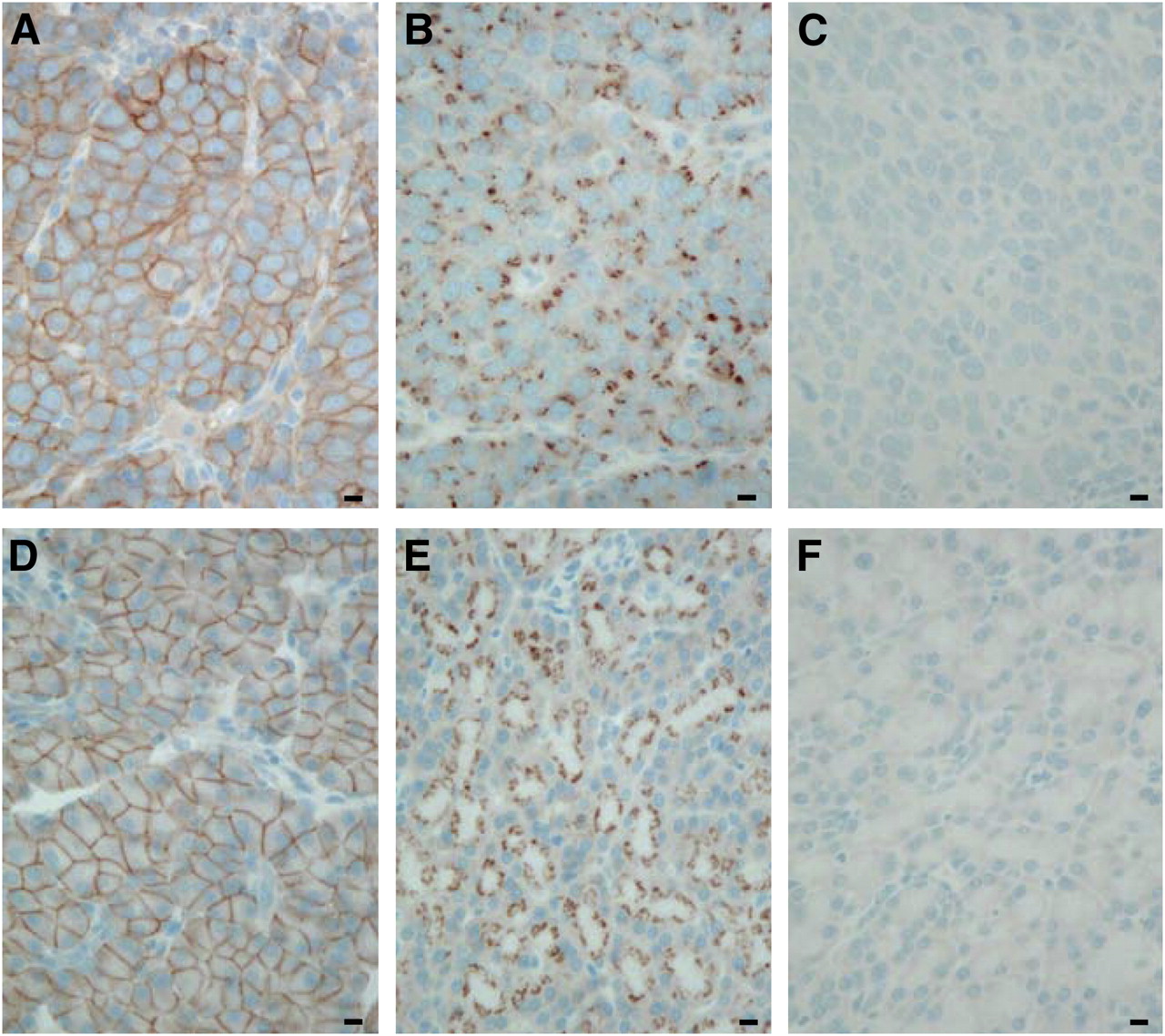
sst2 immunohistochemistry with R2-88 in transplanted AR42J tumor (A–C) and rat pancreas (D–F). (A and D) Tissues taken from untreated animals show membrane-bound sst2. Bars = 0.01 mm. (B and E) Tissues taken from animals sacrificed at 1 h after intravenous injection of TATE (0.21 mg). sst2 receptors have moved from plasma membrane into intracellular structures. Bars = 0.01 mm. (C and F) Preabsorption experiment (control) showing no staining in excess antigen peptide. Bars = 0.01 mm.
In addition, we can confirm with another morphologic method (subtype-selective somatostatin receptor autoradiography) that the AR42J tumor cells express a high density of sst2 receptors. Strong binding of the 125I-[LTT]-SS-28 ligand, which can completely be displaced by sst2-selective ligands but not by sst1-, sst3-, sst4-, or sst5-selective ligands (data not shown), is observed in AR42J tumors, indicating that the strong immunohistochemical staining with R2-88 indeed represents sst2 receptors.
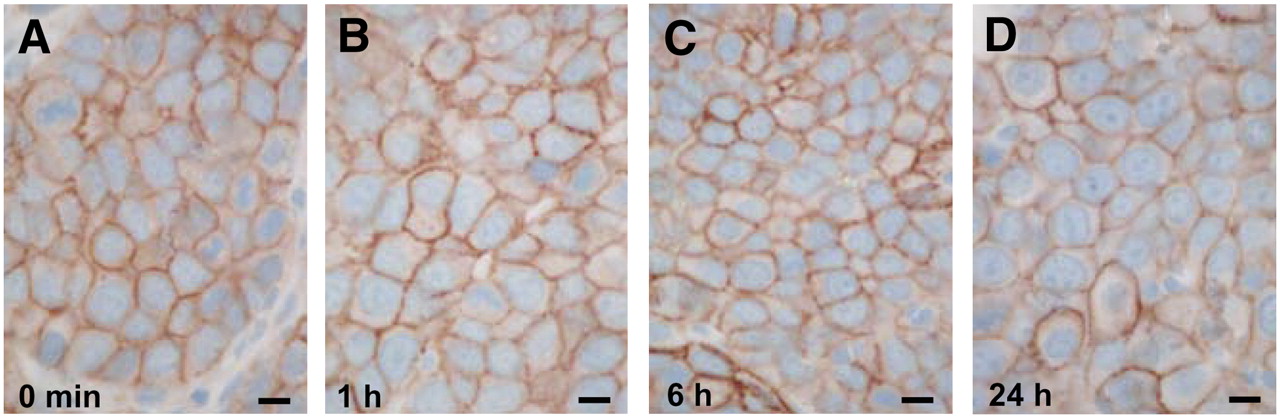
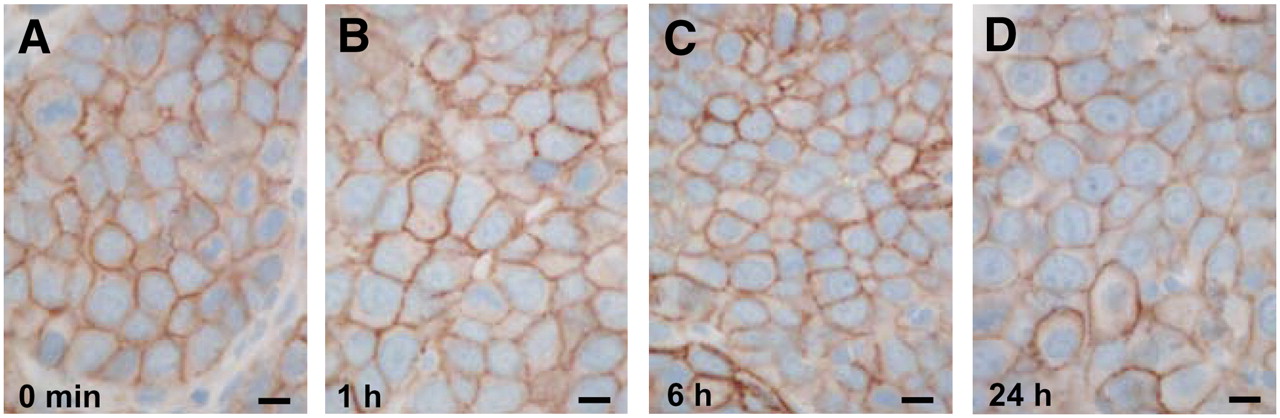
As shown in the kinetic study in Figures 2A–2F, the process of sst2 internalization is essentially achieved 10 min after TATE injection in rats bearing the AR42J tumor and is still detectable at 6 h after injection. Most likely, the effect is fully reversible 24 h after injection (Fig. 2F), because the tumor cells reveal at the 24-h time point an immunohistochemical sst2 distribution similar to that of the untreated control. A similar time course and reversibility of sst2 internalization is observed in the pancreatic tissues (Figs. 2G–2K).

Time course of sst2 internalization in AR42J tumors (A–F) and rat pancreas (G–K), as determined by R2-88 immunohistochemistry. Animals sacrificed at 10 min, 20 min, 1 h, or 6 h after TATE injection (0.21 mg), compared with untreated animals (0 min), showed massive sst2 internalization in tumor and pancreatic cells. However, 24 h after injection of TATE (0.21 mg), sst2 receptors are again found at plasma membrane. Absence of internalization in pancreatic cells at 1 h after injection of sst2 antagonist DOTA-Bass is shown in L. Bars = 0.01 mm.
As expected from previous in vitro studies in HEK-sst2 cells (12), no sst2 internalization was observed in AR42J tumor cells at 1 h after an sst2 antagonist injection (0.21 mg/animal) (Fig. 3). Even at 6 or 24 h after injection, no visible internalization occurred. The same result was seen in the pancreas at the 1-h time point (Fig. 2L).
Lack of sst2 internalization in AR42J tumors from animals sacrificed at 1 (B), 6 (C), or 24 h (D) after intravenous injection of sst2 antagonist DOTA-Bass (0.21 mg), compared with untreated animals (A), as determined by R2-88 immunohistochemistry. Bars = 0.01 mm.
An agonist dose-dependence for sst2 internalization could also be observed: 0.21-mg, 21-μg, or 2.1-μg doses of TATE per animal triggered comparable sst2 internalization in AR42J tumor cells and in pancreatic tissues. However, a 0.21-μg dose was insufficient to produce visible sst2 internalization in both tissues (data not shown).
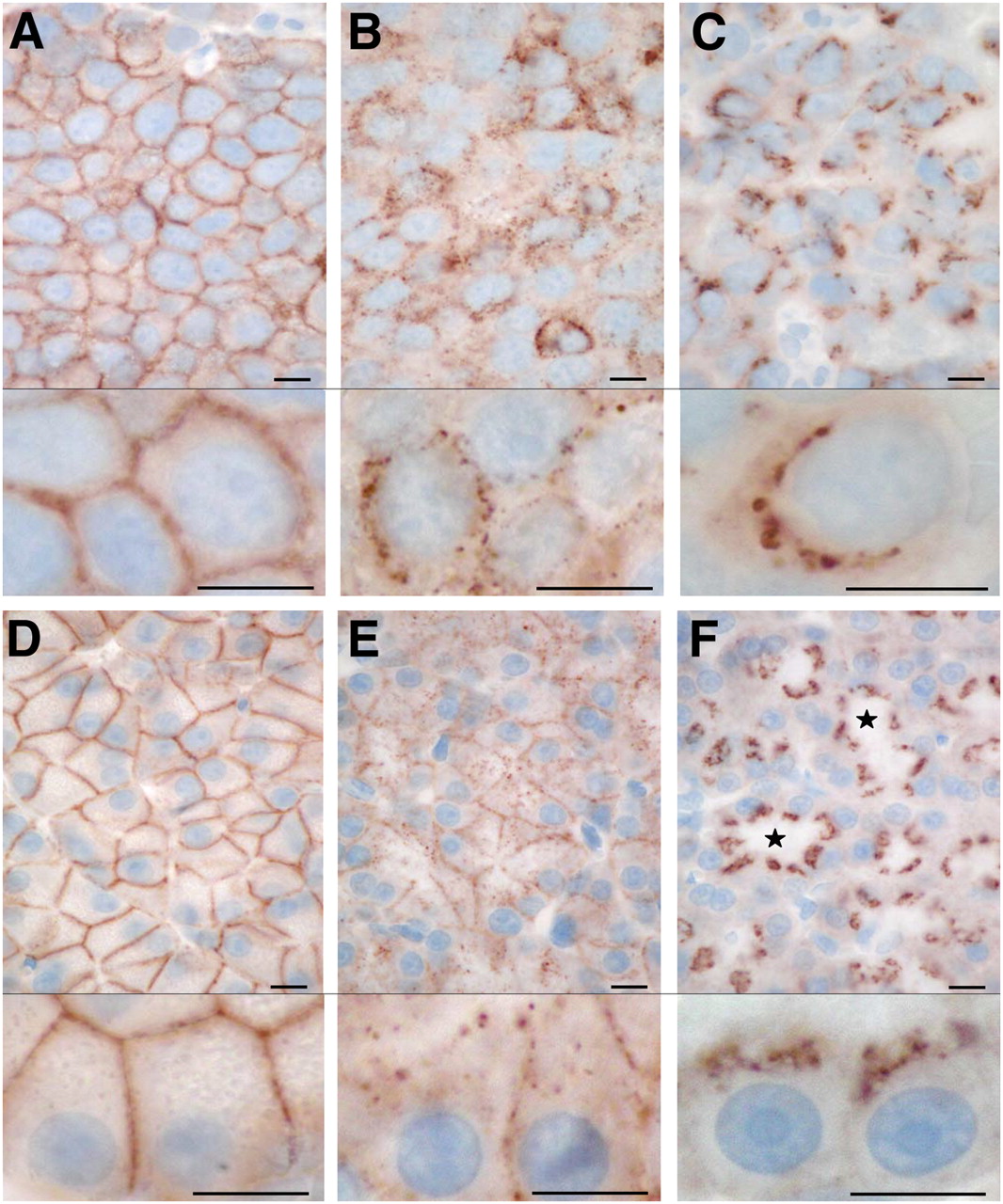
To evaluate how rapidly the internalization process takes place after TATE injection (0.21 mg), a short time of 2.5 min after injection was used. Figure 4 shows sst2 immunohistochemical staining at a higher magnification in AR42J tumors (Figs. 4A–4C) and pancreas (Figs. 4D–4F) and compares this 2.5-min time with a 1-h time and a control (without TATE injection). Whereas the control tissue shows a homogeneous distribution of the receptors at the plasma membrane, the tissues taken 2.5 min after TATE stimulation reveal already a marked change in the topographic sst2 distribution. At 2.5 min, the membranes are generally less intensely stained and, in addition, multiple dotlike structures appear, both within the cell and on the plasma membrane (Figs. 4B and 4E). This distribution is compatible with a fast internalization process of some of the receptors into the cells but also with a possible clustering of some of the receptors in specialized plasma membrane structures just before they internalize (22,24,25). The 2.5-min time reflects a transient phase of sst2 internalization that is considerably different from the receptor distribution at 1 h after TATE injection. At 1 h, indeed, the plasma membrane appears depleted from sst2 receptors; the sst2 receptors are predominantly localized within the cells and are highly concentrated in large intracellular structures resembling endosomes (Figs. 4C and 4F). These observations are valid for both the AR42J tumor cells and the pancreas. Interestingly, the receptor distribution in the pancreatic acinar cells reveals a strong polarity, with the receptors being visible, at rest, in all parts of the plasma membrane except at the luminal side. The internalized sst2 receptors, however, are preferentially localized in intracellular areas situated between the nucleus and ductular lumen of these acinar cells. Such a polarity of sst2 distribution is not readily visible in the AR42J tumor cells.

Localization of internalized sst2 receptors at high magnification in AR42J tumors (A–C) and rat pancreas (D–F), as determined by R2-88 immunohistochemistry. Tissues from untreated animals (A and D) are compared with tissues from animals sacrificed 2.5 min (B and E) and 1 h (C and F) after TATE injection (0.21 mg). sst2 internalization is seen at 2.5-min and 1-h time points. Note polarity of sst2 receptor distribution in pancreatic acinar cells both in untreated animals (D) and animals treated for 1 h (F), in which internalized sst2 are located on luminal side of cells. ⋆ = lumen. Bars = 0.01 mm.
All above-mentioned immunohistochemical results could be reproduced with the 2 commercially available sst2 antibodies, SS-800 and UMB-1. As reported previously, however, SS-800 had a lower sensitivity and higher nonspecific staining (15).
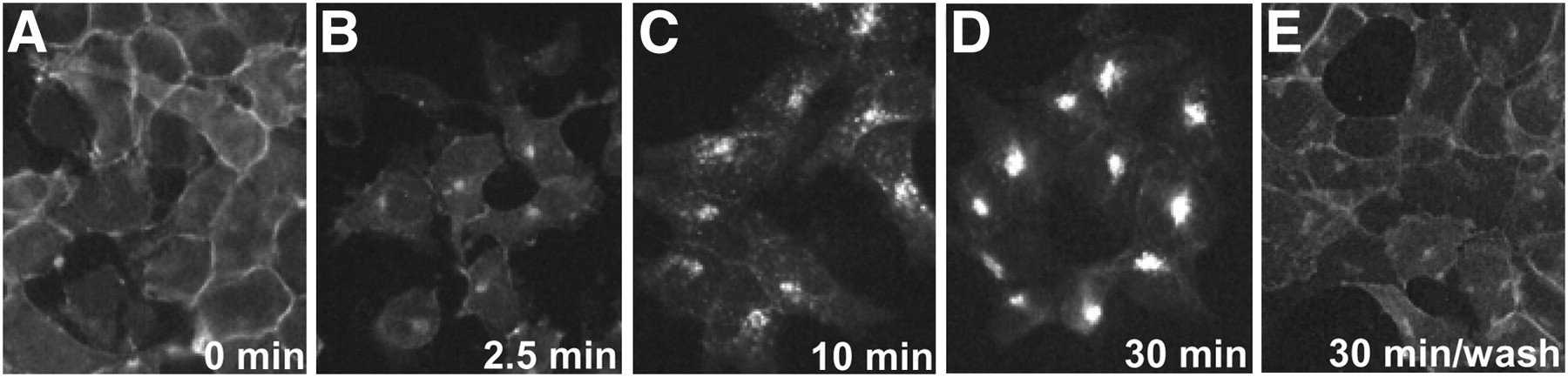
We have compared the in vivo sst2 internalization at 2.5 min with the in vitro agonist-induced sst2 internalization at the same time point observed in cell cultures using HEK293 cells stably expressing the sst2A receptor. Figure 5 illustrates that an early phase of agonist-induced sst2 internalization is also noticed in vitro at 2.5 min, as shown by the monitoring of sst2 trafficking with immunofluorescence microscopy (Fig. 5B). Furthermore, this in vitro sst2 internalization is also completed rapidly within minutes, as seen in the 10- and 30-min times (Figs. 5C and 5D). Finally, when the agonist is removed from the medium (washing step), the sst2 receptors are rapidly relocated to the cell surface as soon as 30 min after washing (Fig. 5E). These results reveal a high degree of correlation between in vivo and in vitro internalization.
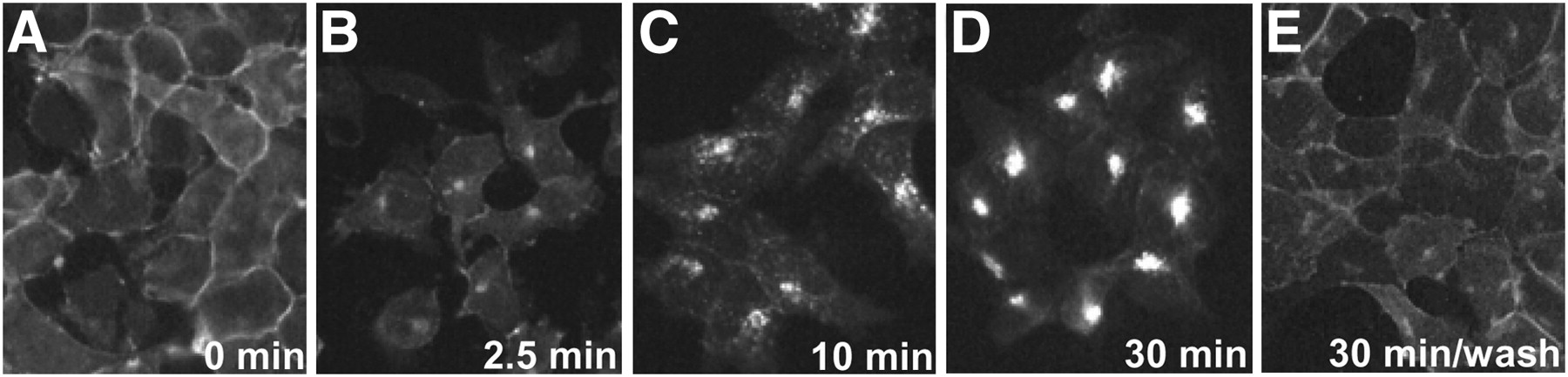
Agonist-induced internalization of sst2 receptor in HEK293. HEK293 cells stably expressing T7-epitope–tagged sst2A receptor (HEK-sst2 cells) were treated either with vehicle alone (A) for 30 min or with 1 μM TATE for 2.5 (B), 10 (C), or 30 min (D) at 37°C and 5% CO2. Moreover, in parallel experiments, cells treated with 1 μM agonist for 30 min at 37°C and 5% CO2 were subsequently washed with PBS and then incubated for 30 min in agonist-free medium at 37°C and 5% CO2 (E). Cells were then fixed, permeabilized, labeled with R2-88 antiserum, and processed for immunocytochemistry. Internalization is already observed at 2.5 min and is completed at 30 min. After subsequent washing and incubation in agonist-free medium, sst2 receptors are back at plasma membrane.
DISCUSSION
Generally, peptide receptor internalization has been evaluated by in vitro methods, namely by following the agonist-induced receptor trafficking by immunofluorescence-based microscopy (12) or by following the uptake of agonist radioligands bound to the internalizing receptor (8). Both methods are using cell culture techniques in vitro. No information is available on the in vivo receptor internalization in cancer tissues, particularly not for the clinically most relevant sst2 receptors.
This study unequivocally shows that sst2 receptors internalize in vivo fast and efficiently, after intravenous application of sst2 agonists. These results are based on an immunohistochemical detection method of sst2 that is not only highly specific, using extensively characterized sst2 antibodies (15), but also morphologic in nature, permitting the identification of the trafficking behavior of the sst2 receptor protein at the cellular level.
In vivo sst2 internalization is efficient in 2 aspects. From a kinetic point of view, in vivo sst2 internalization is an extremely rapid process, taking place within minutes after exposure to the agonist; it is also a powerful mechanism that can relocate many of the plasma membrane receptors to inside the cell.
In vivo sst2 internalization is a highly specific mechanism that is dependant on the type of analog and the type of interaction this analog has with the receptor binding site. It is strongly triggered by an sst2 agonist, but it is not elicited by an equally affine sst2 receptor antagonist, even at the highest concentrations.
In addition, in vivo sst2 internalization is most likely a fully reversible process. Twenty-four hours after TATE injection, the once internalized sst2 receptors appear to be transported back to the plasma membrane, because no visible amount of receptors is left in the intracellular structures. Our previous studies in sst2-expressing HEK293 cells had shown in double labeling experiments that the internalized sst2 receptors costained with an early endosomal marker protein, the mannose 6-phosphate-receptor (22). The pattern of intracellular sst2 receptor distribution seen in the present study is well compatible with an sst2 localization in endosomal structures (22,24,25). The in vivo data suggest that, after internalization, these sst2 receptors are recycled back to the plasma membrane, where they are detected in amounts comparable to the prestimulation levels (24). Furthermore, the data suggest that similarly rapid and potent relocation of internalized sst2 receptors to the plasma membrane is also found in cultured HEK-sst2 cells, only 30 min after the agonist has been removed from the medium. This massive relocation of the internalized sst2 to the cell membrane in such a short time argues in favor of recycled sst2 rather than de novo synthesized sst2 (26).
This in vivo internalization study has evaluated 2 different tissues concomitantly: the AR42J tumor cells and the normal rat pancreatic acinar cells. The same sst2 receptor trafficking behavior can be observed in both tissues, indicating the general reproducibility and significance of this process. The only distinct feature concerns the polarity of the sst2 localization in the pancreatic cells, which may be related to the distinct functions of the 2 tissues. A physiologically relevant tissue such as the exocrine pancreas has its secretory acinar cells consisting of apically and luminally distinct functional areas that may require a specific and polarized sst2 distribution and trafficking. Conversely, the lack of polarity of sst2 distribution in the tumor cells may be related to the more independent and autonomous characteristics of tumor cells that are not or are much less functionally integrated in the host surroundings.
The findings described in this study are important for therapy with somatostatin analogs, particularly in view of the targeted radiotherapy with peptide analogs, such as 177Lu-DOTA-TATE. The fact that internalization is such a rapid process and the relocation of somatostatin receptors on the plasma membrane is so fast offers implications for therapy with both cold and radiolabeled analogs and opens new perspectives in the modalities of fractionation of peptide receptor radiotherapy cycles.
CONCLUSION
These in vivo animal data strongly indicate that the process of sst2 internalization after agonist stimulation is extremely efficient and fully functional under in vivo conditions in neoplastic and physiologic sst2 target tissues. The data provide a strong argument that this molecular process is responsible for the high and long-lasting uptake of sst2 agonist radioligands seen in vivo in sst2-expressing tumors.
Acknowledgments
We thank Dr. Agi Schonbrunn (Houston) for the generous supply of the R2-88 antibody. This study was supported in part by Swiss National Science Foundation grant 32-0000-118333.
Footnotes
-
COPYRIGHT © 2009 by the Society of Nuclear Medicine, Inc.
References
- Received for publication December 19, 2008.
- Accepted for publication February 12, 2009.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A Rapid and Safe Infusion Protocol for 177Lu Peptide Receptor Radionuclide Therapy
- Long-Acting Somatostatin Analog Therapy Differentially Alters 68Ga-DOTATATE Uptake in Normal Tissues Compared with Primary Tumors and Metastatic Lesions
- In Vitro Evaluation of Molecular Tumor Targets in Nuclear Medicine: Immunohistochemistry Is One Option, but Under Which Conditions?
- Radiation Dosimetry for 177Lu-PSMA I&T in Metastatic Castration-Resistant Prostate Cancer: Absorbed Dose in Normal Organs and Tumor Lesions
- Highly Increased 125I-JR11 Antagonist Binding In Vitro Reveals Novel Indications for sst2 Targeting in Human Cancers
- The Somatostatin Analog 188Re-P2045 Inhibits the Growth of AR42J Pancreatic Tumor Xenografts
- Free Somatostatin Receptor Fraction Predicts the Antiproliferative Effect of Octreotide in a Neuroendocrine Tumor Model: Implications for Dose Optimization
- Clinical PET of Neuroendocrine Tumors Using 64Cu-DOTATATE: First-in-Humans Study
- Evaluation of 177Lu-DOTA-sst2 Antagonist Versus 177Lu-DOTA-sst2 Agonist Binding in Human Cancers In Vitro
- Treatment with Octreotide Does Not Reduce Tumor Uptake of 68Ga-DOTATATE as Measured by PET/CT in Patients with Neuroendocrine Tumors
- PET of Somatostatin Receptor-Positive Tumors Using 64Cu- and 68Ga-Somatostatin Antagonists: The Chelate Makes the Difference
- Imaging Expression of the Human Somatostatin Receptor Subtype-2 Reporter Gene with 68Ga-DOTATOC
- Radiolabeled Bicyclic Somatostatin-Based Analogs: A Novel Class of Potential Radiotracers for SPECT/PET of Neuroendocrine Tumors
- Renal Toxicity of Radiolabeled Peptides and Antibody Fragments: Mechanisms, Impact on Radionuclide Therapy, and Strategies for Prevention