


Abstract
Our goals in this study were to determine whether 111In-trastuzumab coupled to peptides harboring nuclear localizing sequences (NLSs) could kill trastuzumab-resistant breast cancer cell lines through the emission of Auger electrons and whether the combination of radiosensitization with methotrexate (MTX) would augment the cytotoxicity of this radiopharmaceutical. Methods: Trastuzumab was derivatized with sulfosuccinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate for reaction with NLS peptides and then conjugated with diethylenetriaminepentaacetic acid for labeling with 111In. HER2 expression was determined by Western blot and by radioligand binding assay using 111In-trastuzumab in a panel of breast cancer cell lines, including SK-BR-3, MDA-MB-231 and its HER2-transfected subclone (231-H2N), and 2 trastuzumab-resistant variants (TrR1 and TrR2). Nuclear importation of 111In-NLS-trastuzumab and 111In-trastuzumab in breast cancer cells was measured by subcellular fractionation, and the clonogenic survival of these cells was determined after incubation with 111In-NLS-trastuzumab, 111In-trastuzumab, or trastuzumab (combined with or without MTX). Survival curves were analyzed according to the dose-response model, and the radiation-enhancement ratio was calculated from the survival curve parameters. Results: The expression of HER2 was highest in SK-BR-3 cells (12.6 × 105 receptors/cell), compared with 231-H2N and TrR1 cells (6.1 × 105 and 5.1 × 105 receptors/cell, respectively), and lowest in MDA-MB-231 and TrR2 cells (0.4 × 105 and 0.6 × 105 receptors/cell, respectively). NLS peptides increased the nuclear uptake of 111In-trastuzumab in MDA-MB-231, 231-H2N, TrR1, and TrR2 cells from 0.1% ± 0.01%, 2.5% ± 0.2%, 2.8% ± 0.7%, and 0.5% ± 0.1% to 0.5% ± 0.1%, 4.6% ± 0.1%, 5.2% ± 0.6%, and 1.5% ± 0.2%, respectively. The cytotoxicity of 111In-NLS-trastuzumab on breast cancer cells was directly correlated with the HER2 expression densities of the cells. On a molar concentration basis, the effective concentration required to kill 50% of 231-H2N and TrR1 cells for 111In-NLS-trastuzumab was 9- to 12-fold lower than for 111In-trastuzumab and 16- to 77-fold lower than for trastuzumab. MDA-MB-231 and TrR2 cells were less sensitive to 111In-NLS-trastuzumab or 111In-trastuzumab, and both cell lines were completely insensitive to trastuzumab. The radiation-enhancement ratio induced by MTX for 231-H2N and TrR1 cells after exposure to 111In-NLS-trastuzumab was 1.42 and 1.68, respectively. Conclusion: Targeted Auger electron radioimmunotherapy with 111In-NLS-trastuzumab can overcome resistance to trastuzumab, and MTX can potently enhance the sensitivity of HER2-overexpressing breast cancer cells to the lethal Auger electrons emitted by this radiopharmaceutical.
Amplification of the HER2/c-erbB-2 proto-oncogene is estimated to occur in 25%–30% of human breast cancers (1), has been correlated with resistance to hormone therapy (2–4) and chemotherapy (5), and is directly associated with poor long-term survival (1). Trastuzumab (Herceptin; Genentech) is a humanized IgG1 anti-HER2 monoclonal antibody (mAb) approved for immunotherapy of HER2-amplified breast cancer (1). Patients are eligible for treatment with trastuzumab on the basis of immunohistochemical staining of a tumor biopsy for HER2 protein overexpression (immunohistochemical scores of 2+ or 3+) or probing for HER2 gene amplification by fluorescence in situ hybridization (6). Even in this highly preselected population, however, only 12%–35% respond to the drug as a single agent, and only half benefit when it is combined with chemotherapy (1,7). Especially disappointing is that almost all patients who respond to trastuzumab become refractory within a year, because of the emergence of resistance (7,8). Clearly, there is a need to substantially improve the effectiveness of trastuzumab and devise alternative treatments for patients whose tumors are resistant to the drug despite HER2 overexpression or that acquire resistance with treatment.
Our group is developing a new class of targeted radioimmunotherapeutics that rely on high-linear-energy-transfer Auger electron emitters in the nanometer-to-micrometer range, such as 111In, to kill cancer cells (9–11). These densely ionizing radionuclides are highly cytotoxic and damaging to DNA when they decay near the cell nucleus, making them highly selective for killing targeted single cancer cells (12). To fully exploit Auger electrons for cancer therapy, however, the radionuclide must be delivered to tumor cells, internalized, and transported to the nucleus, in which these extremely short-range electrons are especially damaging and lethal (13). We recently reported a novel strategy to insert 111In-labeled trastuzumab mAbs directly into the nucleus of HER2-amplified breast cancer cells by conjugating them to 13-mer synthetic peptides (CGYGPKKKRKVGG) harboring the nuclear localizing sequence (NLS) of the simian virus (SV)-40 large T-antigen (11). 111In-NLS-trastuzumab was internalized and imported into the nucleus of HER2-overexpressing breast cancer cells, in which it induced frequent DNA double-strand breaks and strongly decreased the clonogenic survival of the cells. The SV-40 NLS motif has similarly been used by others as a means to route radiolabeled biomolecules into the nucleus of cancer cells for targeted Auger electron radiotherapy of malignancies (14).
Another possible approach to improving the tumoricidal effect of Auger electron radiotherapy is to amplify the potency of the targeted radiation dose through the use of radiosensitizers (15,16). The antimetabolite methotrexate (MTX), for example, is a known potent radiosensitizer that can amplify the lethal effects of ionizing radiation on tumor cells (17). MTX enters cells through the reduced folate carrier-1 and competes with intracellular folate to bind and inhibit the enzyme dihydrofolate reductase, which results in the depletion of folate substrates such as tetrahydrofolate and 5,10-methylenetetrahydrofolate. The lack of reduced folates subsequently causes a critical shortage of thymidylate (deoxythymidine-5′-monophosphate) and purine nucleotides, resulting in arrest of both DNA synthesis and repair (18). The lethal effects of ionizing radiation are therefore exacerbated in thymidylate-depleted cells that have diminished DNA repair capacity (18,19). Our goal in the current study was to capitalize on the potential synergy between targeted Auger electron radiotherapeutics and the radiosensitizing agent MTX, to devise an alternative and potentially much more effective strategy for killing HER2-positive breast cancer cells with acquired trastuzumab resistance. We hypothesized that trastuzumab-resistant breast cancer cells that overexpress HER2 receptors would be sensitive to the cytotoxic effects of 111In-trastuzumab modified with NLS peptides and that MTX would dramatically radiosensitize these cells to the DNA-damaging and lethal effects of the Auger electrons emitted from this radiotherapeutic agent.
MATERIALS AND METHODS
Cell Culture
MDA-MB-231, SK-BR-3, and MCF-7 human breast cancer cells were obtained from the American Type Culture Collection, and the 231-H2N and trastuzumab-resistant-1 and -2 (TrR1 and TrR2) cell lines were kindly provided by Dr. Robert S. Kerbel (Sunnybrook and Woman's College Health Sciences Centre). The 231-H2N cell line was derived from MDA-MB-231 cells that were transfected to stably overexpress c-erbB-2 (HER2); TrR1 and TrR2 cells were isolated from 231-H2N tumors in athymic mice with acquired trastuzumab resistance (20). All cell lines were cultured in Dulbecco's minimal essential medium (Ontario Cancer Institute) supplemented with 10% fetal bovine serum (Sigma-Aldrich) containing 100 U of penicillin per milliliter and 100 μg of streptomycin per milliliter at 37°C in an atmosphere of 5% CO2. SK-BR-3 cells were cultured in RPMI 1640 with 10% fetal bovine serum, 100 U of penicillin per milliliter, and 100 μg of streptomycin per milliliter.
111In-Trastuzumab and 111In-Human IgG Modified with NLS Peptides
Trastuzumab or nonspecific, irrelevent human IgG (hIgG) (product no. I4506; Sigma-Aldrich) was derivatized with diethylenetriaminepentaacetic acid (DTPA) dianhydride (Sigma-Aldrich) and sulfosuccinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (sulfo-SMCC; Pierce) for reaction with synthetic 13-mer NLS peptides (CGYGPKKKRKVGG) and labeling with 111In as described previously (11). Briefly, trastuzumab or hIgG (500 μg, 10 mg/mL) was reacted with a 10-fold molar excess of DTPA for 1 h at room temperature and then purified on a Sephadex-G50 minicolumn (Sigma) before reaction with a 15-fold molar excess of sulfo-SMCC (2–5 mmol/L) at room temperature for 1 h. Maleimide-derivatized DTPA-trastuzumab or hIgG was purified on a Sephadex-G50 minicolumn, transferred to an ultrafiltration device (Microcon YM-50; Amicon), concentrated to 2–5 mg/mL, and reacted overnight at 4°C with a 60-fold molar excess of NLS peptides (5–10 mmol/L diluted in phosphate-buffered saline [PBS], pH 7.0). DTPA-trastuzumab or hIgG, modified with NLS peptides (NLS-DTPA-trastuzumab or NLS-DTPA-hIgG), was purified on a Sephadex-G50 minicolumn eluted with PBS, pH 7.5. We previously determined that 3–4 NLS peptides are conjugated to trastuzumab at a 15:1:60 molar ratio of SMCC:IgG:NLS peptide (11).
NLS-conjugated or unmodified DTPA-trastuzumab or DTPA-NLS-hIgG was radiolabeled by incubation of 37–111 MBq of mAbs with 111InCl3 (MDS-Nordion) for 60 min at room temperature. 111In-labeled mAbs were purified on a Sephadex-G50 minicolumn and buffer exchanged to PBS, pH 7.5, using a Microcon YM-50 ultrafiltration device (Amicon). The radiochemical purity was routinely greater than 97% as determined by instant thin-layer silica-gel chromatography (Pall Corp.) developed in 100 mmol of sodium citrate per liter, pH 5.0. All radioactivity measurements were made using an automatic γ-counter (Wallac Wizard-1480; Perkin Elmer).
Protein Isolation and Western Blot
Cells were grown in T-175 tissue culture flasks (Sarstedt Inc.) to 75%–80% confluency in normal culture conditions. Plates were placed on ice, rinsed with ice-cold PBS, and lysed with cold buffer containing 20 mmol of Tris (pH 7.5) per liter, 137 mmol of sodium chloride per liter, 100 mmol of sodium fluoride per liter, 10% glycerol, and protease inhibitors (Roche). Cells were scraped from plates, incubated on ice for 40 min, and centrifuged at 10,000g for 15 min. Protein concentration of the supernatant was determined using the bicinchoninic acid protein assay kit (Pierce) with bovine serum albumin as a standard.
For Western blot analysis, a 50-μg aliquot of protein was placed in 2× sodium dodecyl sulfate sample buffer containing β-mercaptoethanol and heated for 5 min. Proteins were fractionated on 4%–20% sodium dodecyl sulfate–polyacrylamide minigels and transferred to polyvinyldifluoride membranes (Roche). The polyvinyldifluoride membranes were blocked with 5% skim milk in PBS with 0.1% polysorbate 20 and probed with primary antibody overnight at 4°C. Extracellular erbB2 Ab-20 (L87 + 2ERB19; Medicorp) or β-actin (Sigma-Aldrich) was diluted in a 1:1,000 concentration of 5% milk in polysorbate 20 and detected with horseradish peroxidase–conjugated antimouse IgG (Promega) diluted in a 1:5,000 concentration. Chemiluminescence detection was performed using the Western Lightning Reagent Plus (Perkin Elmer).
Radioligand Binding Assay
The number of HER2 receptors on breast cancer cells was determined by saturation radioligand binding assay at a single excess concentration of 111In-trastuzumab. Approximately 1 × 106 cells were incubated in microtubes for 4 h at 4°C in 250 μL of medium containing 100 nmol of 111In-trastuzumab per liter. This concentration is 1,000-fold greater than the Kd value of 0.1 nmol/L reported by Carter et al. (21) for trastuzumab IgG on HER2-overxpressing SK-BR-3 cells (1.0–2.0 × 106 receptors per cell) and, therefore, was considered high enough to saturate the HER2 receptors on MDA-MB-231, 231-H2N, TrR1, and TrR2 breast cancer cells. After incubation, the cells were pelleted by centrifugation at 1,000g for 5 min, rinsed twice with ice-cold medium to remove unbound radioactivity (supernatant) from total cellular-bound radioactivity (pellets), and measured in a γ-counter. The assay was performed in the absence (total binding) or presence (nonspecific binding) of a 100 μmol/L excess of unlabeled trastuzumab. Subtraction of nonspecific binding from total binding yielded specific binding (nmol/L), which represented the maximum number of HER2 receptors on breast cancer cells assuming a 1:1 molar ratio of trastuzumab to receptor and that all receptors were saturated (i.e., bound to antibody).
In Vitro Nuclear Importation of 111In-NLS-Trastuzumab and 111In-Trastuzumab
Breast cancer cells were plated into T-75 tissue culture flasks (Sarstedt Inc.) at a density of 1 × 107 cells per flask and cultured overnight. After the medium was aspirated, cells were rinsed twice with PBS, pH 7.5, and incubated in medium containing 100 nmol of 111In-NLS-trastuzumab or 111In-trastuzumab per liter for 24 h at 37°C. The 24-h time point was chosen because we have shown previously that nuclear accumulation of 111In-NLS-trastuzumab increases for up to 24 h in HER2-overexpressing breast cancer cells (11). After the incubation period, the medium was decanted, and the cells were rinsed twice with ice-cold PBS, pH 7.5, to remove unbound radioactivity that was measured in a γ-counter. The cells were then harvested by scraping them into 1 mL of Nuclei EZ Lysis buffer (Sigma-Aldrich) and lysed according to the manufacturer's instructions. Nuclei (pellets) were separated from the cytoplasmic or membrane fraction (supernatant) by centrifugation for 5 min at 1,000g, and the radioactivity in each fraction was measured in a γ-counter. We previously determined that this cell fractionation procedure yields a very pure nuclear fraction (22).
Clonogenic Assays
Approximately 2 × 106 breast cancer cells were incubated with 111In-NLS-trastuzumab (1–450 nmol/L; 190 ± 8.9 MBq/mg) alone or concurrently with a single, nontoxic concentration of MTX (i.e., concentration of drug resulting in >90% cell survival) in 1 mL of culture medium in microtubes for 24 h at 37°C. Controls consisted of cells treated with normal saline, MTX (0.001–100 μmol/L), 111In-trastuzumab (182 ± 10.5 MBq/mg), unlabeled trastuzumab, or 111In-labeled nonspecific hIgG modified with NLS (171.0 ± 12.2 MBq/mg). For treatments including MTX, cells were serum-starved by lowering the serum concentration from 10% to 1% to reduce the concentrations of thymidine, 5-methyl tetrahydrofolate, and purine ribonucleosides that counteract the effects of MTX (23). After treatment, the cells were centrifuged at 1,000g for 5 min and washed twice with normal culture medium containing 10% serum. Sufficient cells were then plated in triplicate in 12-well plates and cultured in normal medium at 37°C. After 10–14 d, colonies of a least 50 cells were stained with methylene blue and counted. The surviving fraction was calculated by dividing the number of colonies formed for treated cells by the number for untreated cells. Survival curves were derived by plotting the surviving fraction values versus the log molar concentration of antibody used, and the effective concentration to kill 50% (EC50) was estimated using Origin 6.0 (Microcal Software, Inc.), applying the dose-response equation y = A1 + ([A2 − A1]/[1 + 10(log EC50−x) × p]), where p is the slope (set to equal −1), and A1 and A2 are the amplitudes of the baseline and maximum response, respectively. The mean inactivation dose (MID) (area under the survival curve) and the radiation-enhancement ratio (RER) for MTX (RER = MID [111In-NLS-trastuzumab]/[111In-NLS-trastuzumab + MTX]) were also determined, and an RER value greater than 1 indicated radiosensitization by MTX (24).
Statistical Methods
Data are presented as mean ± SEM. Statistical comparisons were made using the Student t test. A P value of less than 0.05 was considered significant.
RESULTS
Expression Levels of HER2 Extracellular Domain
Trastuzuamb binds HER2 on the C-terminal portion of the extracellular domain (ECD) near the juxtamembrane region in domain IV of the receptor (25). We therefore evaluated the expression of the HER2-ECD by Western blot in a panel of human breast cancer cell lines, including trastuzumab-sensitive SK-BR-3, trastuzumab-insensitive MDA-MB-231 and its HER2-transfected subclone (231-H2N), and 2 trastuzumab-resistant subclones of 231-H2N cells (TrR1 and TrR2) (11,20). As shown in Figure 1A, the HER2-ECD was abundantly expressed at a high level in whole-cell lysates from SK-BR-3 cells and at an intermediate level in 231-H2N and TrR1 cells. In contrast, the HER2-ECD was not detected in the lysates from MDA-MB-231 or TrR2 cells (Fig. 1A), in agreement with previously reported data (20).
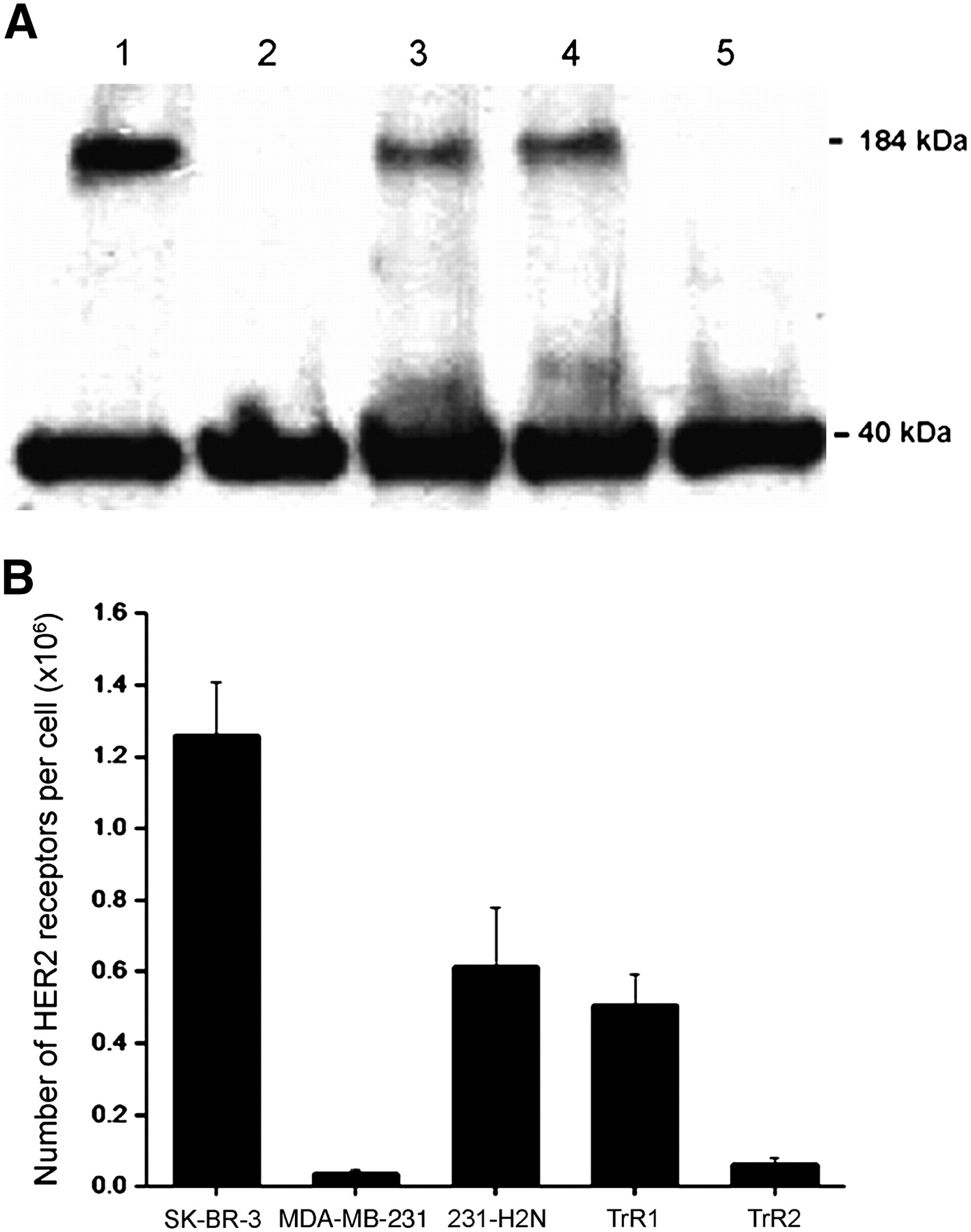
(A) Western blot for expression of HER2 ECD (184 kDa) in SK-BR-3 (lane 1), MDA-MB-231 (lane 2), 231-H2N (lane 3), TrR1 (lane 4), and TrR2 (lane 5) human breast cancer cells. β-actin (40 kDa) was used as loading control. (B) Number of HER2 receptors on these cells determined by radioligand binding assay using saturating concentration of 111In-trastuzumab, with or without excess unlabeled trastuzumab (10 μmol/L).
The number of HER2 receptors in these cells was also quantified by radioligand binding assay using a single saturating concentration of 111In-trastuzumab, with or without excess unlabeled trastuzumab (10 μmol/L). The number of HER2 receptors for SK-BR-3 cells was approximately 2-fold greater than that for 231-H2N and TrR1 cells (12.6 × 105 receptors per cell vs. 6.1 × 105 and 5.1 × 105 receptors per cell, respectively) and approximately 10-fold higher than the number of HER2 receptors for MDA-MB-231 and TrR2 cells (0.4 × 105 and 0.6 × 105 receptors per cell, respectively) (Fig. 1B).
Nuclear Importation of 111In-NLS-Trastuzumab and 111In-Trastuzumab in Breast Cancer Cells
Nuclear importation of 111In in breast cancer cells increased in accordance with the HER2 density of the cells when exposed to 111In-NLS-trastuzumab or 111In-trastuzumab (Fig. 2). After 24 h, the nuclear uptake of 111In-NLS-trastuzumab in MDA-MB-231, 231-H2N, TrR1, and TrR2 breast cancer cells, with respect to the total amount of radioactivity added to incubation medium, was 0.5% ± 0.1%, 4.6% ± 0.1%, 5.2% ± 0.6%, and 1.5% ± 0.2%, respectively, compared with 0.1% ± 0.01%, 2.5% ± 0.2%, 2.8% ± 0.7%, and 0.6% ± 0.1% for 111In-trastuzumab.
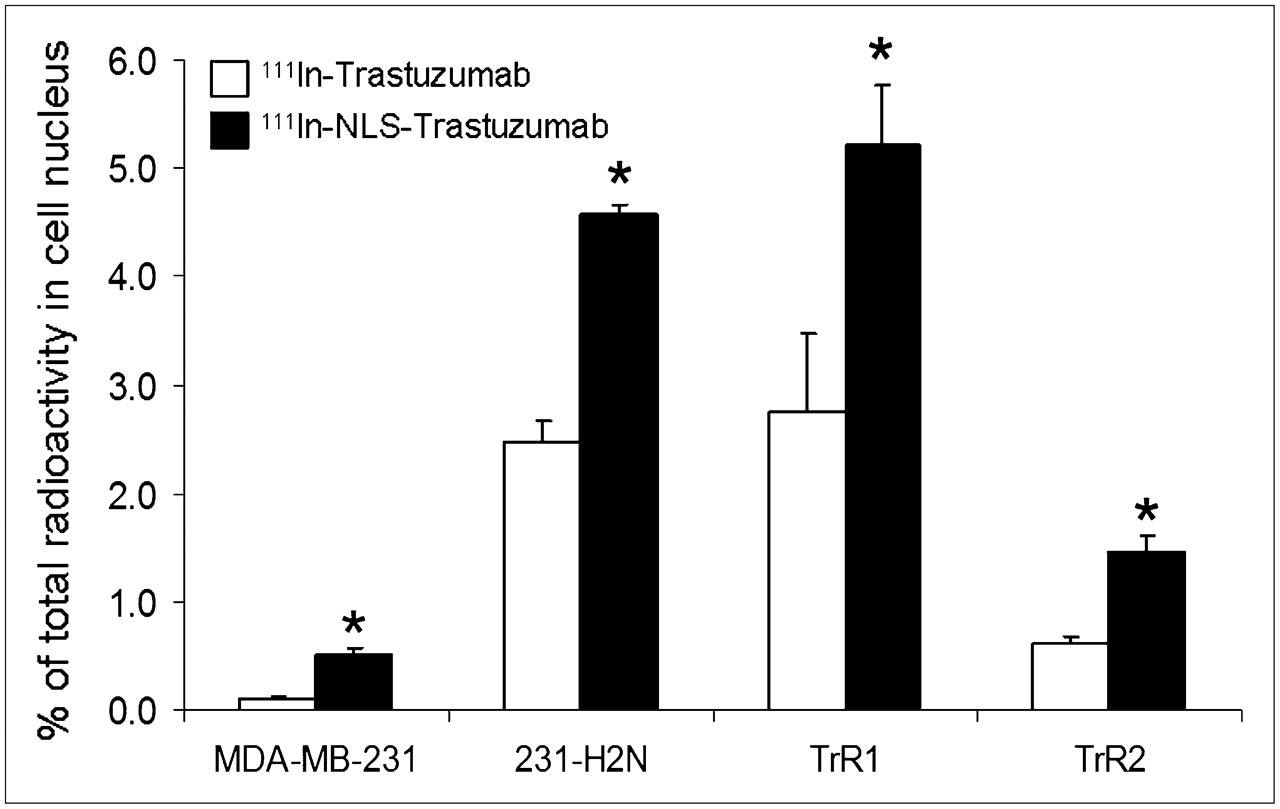
Nuclear importation of 111In-trastuzumab and 111In-NLS-trastuzumab by MDA-MB-231, 231-H2N, TrR1, and TrR2 human breast cancer cells with total amount of radioactivity added to incubation medium. Values shown are mean ± SEM of 4 independent determinations (*P < 0.05).
Effect of 111In-NLS-Trastuzumab and 111In-Trastuzumab on the Survival of Breast Cancer Cells
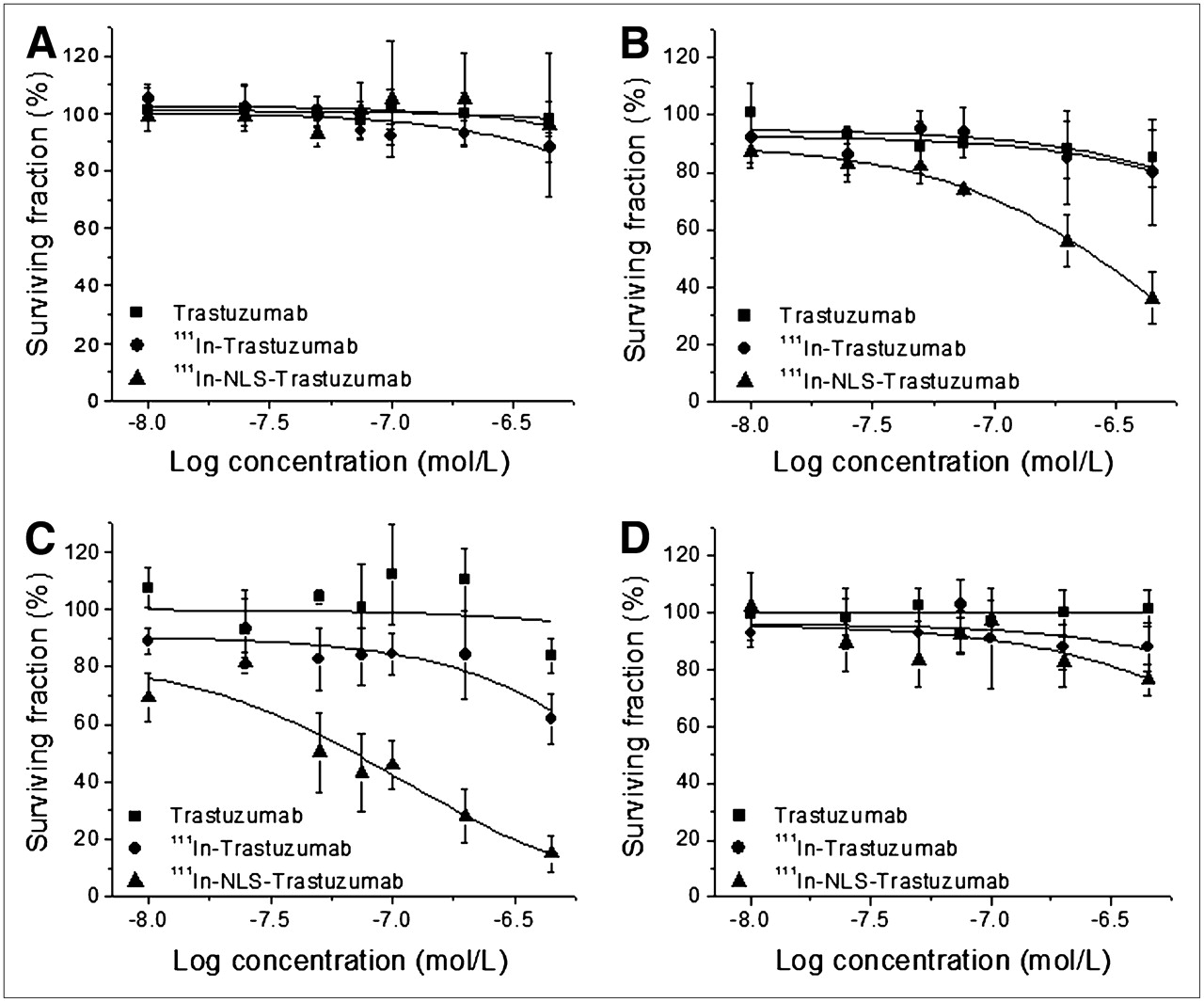
The cytotoxicity of 111In-NLS-trastuzumab against trastuzumab-sensitive and trastuzumab-resistant breast cancer cells was evaluated by clonogenic assay. A strong dose-dependent decrease in colony formation of 231-H2N and TrR1 cells treated with increasing amounts of 111In-NLS-trastuzumab (Figs. 3B and 3C) was demonstrated. The EC50 of 231-H2N cells for 111In-NLS-trastuzumab was 9- and 16-fold lower than that for 111In-trastuzumab or unlabeled trastuzumab (Table 1). Similar results were observed for TrR1 cells, in which the EC50 for 111In-NLS-trastuzumab was significantly lower than that for 111In-trastuzumab or unlabeled trastuzumab. In contrast, TrR2 and MDA-MB-231 cells were less sensitive to 111In-NLS-trastuzumab or 111In-trastuzumab, and both cell lines were completely refractory to unlabeled trastuzumab (Figs. 3A and 3D; Table 1). No significant toxicity toward 231-H2N cells after treatment with nonspecific, irrelevant 111In-NLS-hIgG control immunoconjugates (data not shown) was observed.

Clonogenic survival of trastuzumab-insensitive MDA-MB-231 (A), trastuzumab-sensitive 231-H2N (B), and trastuzumab-resistant TrR1 (C) and TrR2 (D) human breast cancer cells after exposure to increasing concentrations of trastuzumab, 111In-trastuzumab, or 111In-NLS-trastuzumab. Each point represents mean ± SEM of 3–5 experiments performed in triplicate.
EC50 for Trastuzumab, 111In-Trastuzumab, and 111In-NLS-Trastuzumab on Breast Cancer Cells with Different HER2 Expression and Trastuzumab-Resistance Status*
Effect of MTX Treatment on the Survival of Breast Cancer Cells
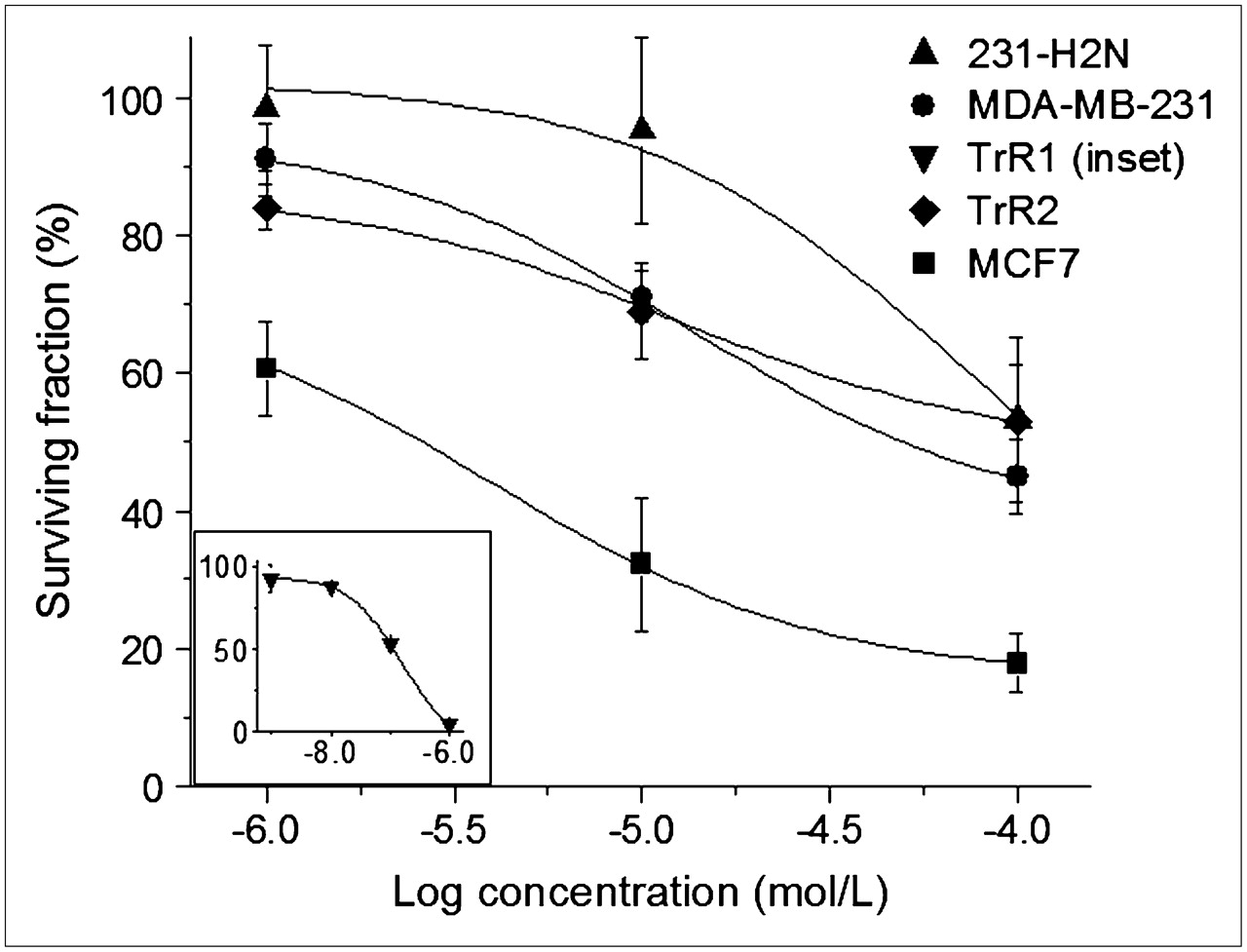
The toxicity of exposure to MTX for 24 h for the different breast cancer cell lines was assessed by clonogenic assay before its ability to sensitize these cells to Auger electron radiation was evaluated. As illustrated in Figure 4, MTX exerted a dose-dependent reduction in the clonogenic survival of serum-starved breast cancer cells. MCF-7 human breast cancer cells have previously been shown to be MTX-sensitive (26) and were therefore used as positive controls. This cell line exhibited a 50-fold greater sensitivity to MTX than did MDA-MB-231, 231-H2N, and TrR2 cells (EC50, 2.3 ± 0.6 μmol/L vs. 103.3 ± 13.4 μmol/L, 91.7 ± 13.9 μmol/L, and 102.4 ± 10.7 μmol/L, respectively). In contrast, TrR1 cells gave an EC50 of 0.14 ± 0.03 μmol/L, which was 10 times lower than that for MCF-7 cells. The reason for this high sensitivity of TrR1 cells to MTX is not known.

Clonogenic survival of serum-starved MCF7, MDA-MB-231, 231-H2N, TrR1 (inset), and TrR2 human breast cancer cells after exposure to increasing concentrations of MTX. Each point represents mean ± SEM of 3 experiments performed in triplicate.
Effect of Combined Treatment with 111In-NLS-Trastuzumab or Unlabeled Trastuzumab and Low-Dose MTX
The cytotoxicity of 111In-NLS-trastuzumab was most evident against 231-H2N and TrR1 cells. Therefore, a fixed and noncytotoxic concentration of MTX resulting in greater than 90% survival of 231-H2N and TrR1 cells was chosen to evaluate the effect of combined treatment with 111In-NLS-trastuzumab or unlabeled trastuzumab. For 231-H2N and TrR1 cells, the lowest noncytotoxic dose of MTX was determined to be 10 μmol/L and 0.01 μmol/L, respectively. Figure 5 compares the survival curves of serum-starved cells after combined treatment with low-dose MTX and increasing concentrations of 111In-NLS-trastuzumab or unlabeled trastuzumab and of serum-starved cells treated with 111In-NLS-trastuzumab alone. The EC50 for 111In-NLS-trastuzumab against 231-H2N and TrR1 cells treated concurrently with low-dose MTX was approximately 5-fold lower than that for cells treated with only 111In-NLS-trastuzumab (Table 2). The RER induced by MTX for 231-H2N and TrR1 cells after exposure to 111In-NLS-trastuzumab was estimated to be 1.42 and 1.68, respectively. Low-dose MTX had no effect on the EC50 value elicited by trastuzumab in TrR1 and 231-H2N cells, compared with treatment with trastuzumab alone (Tables 1 and 2).
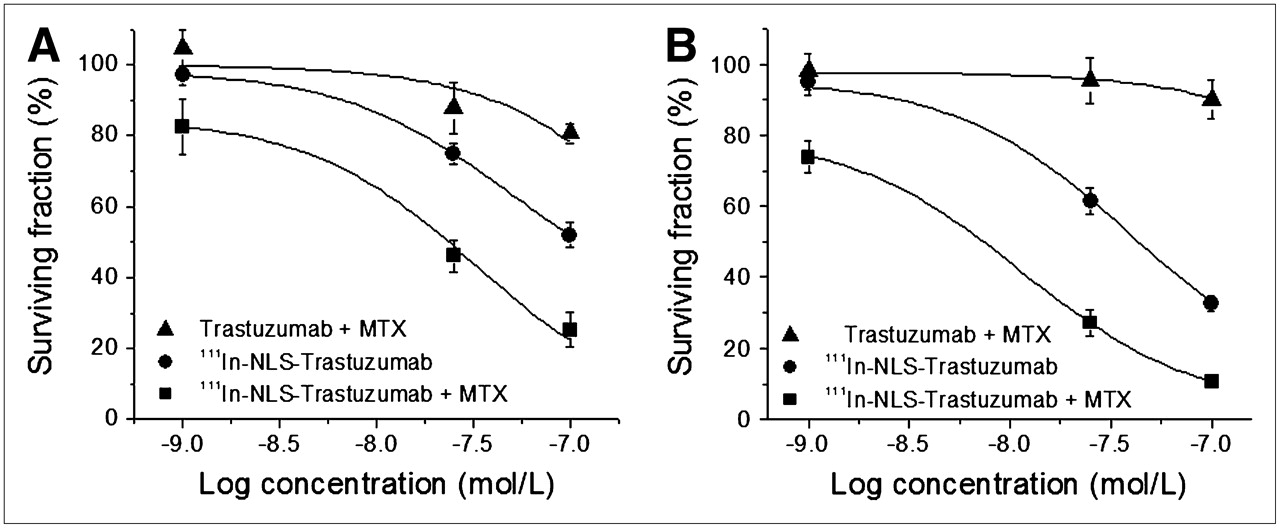
Clonogenic survival of serum-starved 231-H2N (A) and TrR1 (B) breast cancer cells after exposure to trastuzumab plus MTX, 111In-NLS-trastuzumab alone, or 111In-NLS-trastuzumab plus MTX. Each point represents mean ± SEM of 3 experiments performed in triplicate.
EC50 for 111In-NLS-Trastuzumab Alone, 111In-NLS-Trastuzumab Plus MTX, and Trastuzumab Plus MTX on Serum-Starved 231-H2N and TrR1 Breast Cancer Cells*
DISCUSSION
Our results demonstrated that 111In-labeled trastuzumab, modified with 13-mer peptides [CGYGPKKKRKVGG] harboring the NLS of SV-40 large T-antigen (underlined), can kill HER2-positive breast cancer cells resistant to trastuzumab. Furthermore, noncytotoxic concentrations of MTX potently radiosensitized these cells to the high-linear-energy-transfer and lethal Auger electron radiation emitted by 111In-NLS-trastuzumab. Resistance to trastuzumab is a major challenge in the treatment of metastatic breast cancer (27). Even in responding patients, the effectiveness of trastuzumab is limited and the duration of response short (7,8). If the results of this study are promising after in vivo testing, then the combination of radiosensitizing chemotherapy and targeted Auger electron radiotherapy using 111In-NLS-trastuzumab represents a viable strategy to address this resistance issue.
The 231-H2N cell line is a HER2-transfected variant of MDA-MB-231 human breast cancer cells; these cells are growth-inhibited by trastuzumab when grown as orthotopic tumors in severe combined immunodeficient mice (20). In our study, however, only a 20% reduction in the clonogenic survival of 231-H2N cells was observed after incubation with trastuzumab (Fig. 3). Many mechanisms are linked to the therapeutic effect of trastuzumab, some of which, like angiogenesis suppression, are observed only in an in vivo tumor xenograft model (27). 231-H2N cells were nevertheless efficiently killed by exposure to increasing concentrations of 111In-NLS-trastuzumab. TrR1 cells were similarly killed by 111In-NLS-trastuzumab, which is intriguing because this variant cell line was derived from a mouse bearing a 231-H2N tumor xenograft with acquired resistance to trastuzumab (20). TrR2 cells were also isolated from trastuzumab-resistant 231-H2N tumor xenografts (20); however, this variant cell line was less sensitive to 111In-NLS-trastuzumab or 111In-trastuzumab and was completely refractory to trastuzumab, likely because of a 10-fold lower HER2 density (Figs. 1 and 3).
The mechanism of resistance of TrR1 and TrR2 cells to trastuzumab is not known, but multiple mechanisms by which HER2-positive breast cancer cells are able to evade the antiproliferative effects of the drug exist, including the loss of protein tyrosine phosphatase function (28), downregulation of the cyclin-dependent kinase inhibitor p27kip1 (29), or upregulation of epidermal growth factor or insulinlike growth factor-I receptors that compensate for the loss of HER2 signaling (30,31). Despite the emergence of these resistant pathways that block the growth-inhibiting effects of trastuzumab, the HER2-mediated internalization and nuclear importation of 111In-NLS-trastuzumab in 231-H2N and TrR1 cells was preserved, irrespective of their trastuzumab resistance status (Fig. 2). Thus, to our knowledge, these findings suggest for the first time that 111In-NLS-trastuzumab could potentially be used to treat trastuzumab-resistant tumors in patients. Moreover, it is possible that targeted Auger electron radiotherapy may be able to overcome other forms of treatment resistance in breast cancer and other malignancies. For example, we previously demonstrated that 111In-NLS-trastuzumab can kill MDA-MB-361 and SK-BR-3 breast cancer cells (11), which are known to have high levels of the multidrug-resistant transporter p-glycoprotein and are resistant to paclitaxel (32). In addition, we have shown that 111In-labeled anti-CD33 mAbs conjugated to NLS peptides can kill HL-60/MX-1 myeloid leukemia cells that are resistant to mitoxantrone and patient AML specimens that display high levels of multidrug-resistant transporters, including p-glycoprotein (33).
The cytotoxicity of 111In-NLS-trastuzumab on breast cancer cells was also directly correlated with the HER2 expression densities of the cells, with the greatest toxicity observed for 231-H2N and TrR1 cells and the least toxicity found for MDA-MB-231 and TrR2 cells. These results confirm those previously reported for 111In-NLS-trastuzumab against trastuzumab-sensitive SK-BR-3 (1–2 × 106 receptors/cell) and MDA-MB-361 (0.5 × 106 receptors/cell) breast cancer cells, in which the radiopharmaceutical was significantly more potent at killing these cells than unlabeled trastuzumab (11). Nuclear localization is critically important for exploiting the lethality of Auger electron–emitting radiopharmaceuticals toward cancer cells because more than 99% of the low-energy Auger electrons emitted by 111In travel less than 1 μm in tissue. Consequently, the radiation-absorbed dose deposited in the nucleus is amplified 17 times when 111In is deposited in the nucleus, compared with when it is localized in the cytoplasm, and more than 34 times when 111In is deposited in the nucleus, compared with when it is localized on the cell membrane (14). Indeed, microdosimetry analysis of MDA-MB-361 breast cancer cells exposed to 111In-NLS-trastuzumab revealed that the 80% of radioactivity bound to the cell surface and imported into the cytoplasm accounts for approximately 10% of the radiation-absorbed dose to the nucleus, whereas the 20% of radioactivity imported into the nucleus accounts for nearly 90% of the radiation dose to the nucleus (11).
Trastuzumab is commonly combined with taxane- or platinum-based chemotherapy regimes (27), but it was recently demonstrated that significant clinical benefit could be achieved in patients refractory to trastuzumab by combining the drug with oral low doses of cyclophosphamide and MTX (34). The goal of low-dose chemotherapy is to lessen the side effects of treatment and inhibit tumor growth, possibly through an antiangiogenic effect (20,34). In these studies, however, we exploited a new opportunity to use similarly low, clinically achievable levels of MTX (<10 μmol/L) to enhance the radiosensitivity of breast cancer cells to 111In-NLS-trastuzumab. The differential sensitivity of breast cancer cells to MTX was not further assessed, but antimetobolites are among the most potent radiosensitizers and are often active at concentrations below those necessary to induce cytotoxicity (35). Indeed, other studies have demonstrated that the efficacy of Auger electron radiotherapy can be significantly enhanced by MTX. For example, the administration of MTX before the injection of 5-[125I]iodo-2′-deoxyuridine (125IUdR) in rats with intrathecal TE-671 human rhabdomyosarcoma enhanced 125IUdR uptake by tumor cells and augmented the therapeutic efficacy of this Auger electron–emitting radiopharmaceutical (36). Paclitaxel, docetaxel (Taxotere; Sanofi-Aventis), gemcitabine, topotecan, 5-fluorouracil, doxorubicin, tirapazamine, or halogenated pyrimidines have also been combined with targeted α- or β-radioimmunotherapy and have been shown to inhibit the growth of various malignant cell lines in vitro or provide antitumor effects in mouse xenograft models and in human clinical trials more effectively than either treatment alone (15,16).
MTX exerts its toxicity by inhibiting purine de novo synthesis, which is thought to impede DNA synthesis and repair and augment cell killing with ionizing radiation (18). Alternatively, the redistribution of cells to a more radiosensitive phase of the cell cycle (i.e., at the G1/S boundary or early S phase) or elimination of radioresistant S phase cells have also been suggested as mechanisms for MTX radiosensitization (17). The latter mechanism seems unlikely, however, because nontoxic concentrations of MTX were used to radiosensitize breast cancer cells to the DNA-damaging and lethal effects of the Auger electron emissions from 111In-NLS-trastuzumab. Regardless of the mechanism, there is the potential that this combination may also increase the radiotoxicity on normal cells that accumulate the agent in vivo (e.g., liver and kidney). However, toxicity assessments in mice currently in progress show the regimen to be well tolerated, although it should be appreciated that trastuzumab does not cross-react with mouse c-erbB-2 (homolog of human HER2) and thus it remains that cardiotoxicity could potentially be a side effect associated with this treatment (37). As in the case with trastuzumab, it may be possible to evaluate this only in clinical trials of the radiopharmaceutical.
CONCLUSION
We demonstrated that 13-mer synthetic peptides harboring the NLS of SV-40 large T-antigen efficiently routed 111In-trastuzumab to the nucleus of trastuzumab-sensitive and trastuzumab-resistant human breast cancer cell lines overexpressing HER2, in which the nanometer-to-micrometer Auger electrons rendered the radiotherapeutic agent lethal to these cells. In addition, 111In-NLS-trastuzumab, combined with low doses of MTX, is more cytotoxic to these cells than either of these agents alone. Overcoming trastuzumab resistance represents an important challenge to improving outcomes for patients with HER2-amplified metastatic breast cancer. Achieving a synergistic interaction by combining MTX and targeted Auger electron radioimmunotherapy could dramatically improve the response, compared with either of these therapies alone.
Acknowledgments
This research was supported by grant 016456 from the Canadian Breast Cancer Research Alliance, with funds from the Canadian Cancer Society, the Canadian Breast Cancer Foundation (Ontario Chapter), the Connaught Fund, and the Leslie Dan Faculty of Pharmacy. This research was also supported by the Canadian Breast Cancer Foundation–Ontario Region awarded to Danny Costantini, the Canadian Institutes of Health Research (CIHR) awarded to Kristin McLarty, and a Life Sciences Committee Undergraduate Research Opportunity Award to Katherine Bateman. Both Danny Costantini and Kristin McLarty are alumni of the CHIR–Excellence in Radiation Research for the 21st Century (EIRR21) Strategic Training Program.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication February 13, 2008.
- Accepted for publication May 28, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Targeting Micrometastases: The Effect of Heterogeneous Radionuclide Distribution on Tumor Control Probability
- Dual-Receptor-Targeted Radioimmunotherapy of Human Breast Cancer Xenografts in Athymic Mice Coexpressing HER2 and EGFR Using 177Lu- or 111In-Labeled Bispecific Radioimmunoconjugates
- Auger Radiopharmaceutical Therapy Targeting Prostate-Specific Membrane Antigen
- Kinase Activator-Receiver Preference in ErbB Heterodimers Is Determined by Intracellular Regions and Is Not Coupled to Extracellular Asymmetry
- Evaluation of 99mTc-peptide-ZHER2:342 Affibody(R) molecule for in vivo molecular imaging
- ErbB-2 Blockade and Prenyltransferase Inhibition Alter Epidermal Growth Factor and Epidermal Growth Factor Receptor Trafficking and Enhance 111In-DTPA-hEGF Auger Electron Radiation Therapy
- Antitumor Effects and Normal-Tissue Toxicity of 111In-Nuclear Localization Sequence-Trastuzumab in Athymic Mice Bearing HER-Positive Human Breast Cancer Xenografts
- Methotrexate, Paclitaxel, and Doxorubicin Radiosensitize HER2-Amplified Human Breast Cancer Cells to the Auger Electron-Emitting Radiotherapeutic Agent 111In-NLS-Trastuzumab