


Abstract
System L amino acid transport is increased in various types of cancer. The tracer 123I-2-iodotyrosine (2IT), which is accumulated via system L, could thus serve to allow visualization of cancer in vivo. Here, we studied the transport of 125I-2IT by h4F2hc-hLAT1, the major transporter subserving system L in growing cells, using the Xenopus laevis oocyte expression system. We compared the apparent affinity of 125I-2IT with that of tyrosine, tested the influence of intracellular methionine availability on the influx rate of this substrate, and then compared the transport of 2IT with that of the other tracers—iodo-α-methyltyrosine (IMT), fluoroethyltyrosine (FET), and 2-fluorotyrosine (2FT)—by measuring their transstimulating effect on phenylalanine efflux. Methods: Transport experiments were performed with Xenopus laevis oocytes expressing h4F2hc-hLAT1 (the functional transporter) and oocytes expressing only h4F2hc (negative control). The values obtained for the functional transporter were corrected for endogenous background transport by subtracting the values for the negative controls. Results: The apparent affinity for 125I-2IT and 3H-tyrosine was 29.3 ± 9.3 μmol/L and 21.2 ± 4.2 μmol/L, respectively. The influx rate of 125I-2IT was, similarly to that of 3H-phenylalanine, transstimulated by a factor of ≥3 when the oocytes were preinjected with methionine or phenylalanine. The proportion of preinjected 3H-phenylalanine that effluxed within 90 s in the presence of an extracellular 2IT concentration of 0.1 mmol/L was 4.1% ± 0.5%, compared with 3.3% ± 0.4% for extracellular IMT, 1.3% ± 0.3% for FET, 9.3% ± 0.8% for 2FT, and 9.1% ± 0.5% for phenylalanine. Conclusion: 2IT has a high affinity for h4F2hc-hLAT1, comparable to that of natural tyrosine, and its influx rate is transstimulated by intracellular amino acids. The 2IT influx rate is comparable to that of IMT but lower than that of phenylalanine. In contrast to FET, which is only poorly transported, 2FT displays a high influx rate equal to that of phenylalanine.
System L amino acid transporters are the main pathway for the cellular uptake of large branched and neutral amino acids (1). This type of transport activity is strongly increased in many cancer cells (2–4). In vitro studies have shown that several 123I- and 18F-labeled aromatic amino acid tracers, including 123I-2-iodotyrosine (2IT), accumulated mainly via L-type transport (5–7). Therefore, these tracers offer the opportunity to study the system L transport activity of human cancer in vivo with SPECT or PET.
At present, 3 transporter subtypes that generate L transport have been cloned: LAT1-4F2hc, LAT2-4F2hc, and LAT3 (8–11). In this study, we focused on the LAT1-4F2hc subtype because studies have shown that it is overexpressed in many transformed cell lines and that its expression correlates with tumor growth (2–4). At the molecular level, the LAT1 transporter is a heterodimer comprising a heavy-chain glycoprotein (h4F2hc) and a catalytic light chain (hLAT1) that are covalently linked by a disulfide bond. Both subunits are required for functional cell-surface expression. The transporter functions as an obligatory exchanger with a 1:1 stoichiometry, and the activity of transport depends on the availability of intracellular substrate amino acids (12). This means that net transport of a given amino acid can be obtained only in exchange for other amino acids that might have been taken up by another (unidirectional) amino acid transporter. Several authors have suggested that methionine could serve as the recycling exchange substrate because it is intensely accumulated via system A and also is a good intracellular substrate for LAT1-mediated exchange (1,13).
The aim of our study was to functionally characterize the interaction of 125I-2IT with the LAT1 transporter. For this purpose we coexpressed the h4F2hc and hLAT1 subunits in Xenopus laevis oocytes. First, the affinity of 125I-2IT for the extracellular side of the h4F2hc-hLAT1 transporter was measured and compared with natural tyrosine. Second, we tested the influence of intracellular amino acid availability on the influx rate of 125I-2IT. Finally, we indirectly measured the influx rate of 2IT in comparison with that of similar SPECT (iodo-α-methyltyrosine [IMT]) and PET (fluoroethyltyrosine [FET] and 2-fluorotyrosine [2FT]) tracers using an efflux assay (14).
MATERIALS AND METHODS
Complementary RNA (cRNA) Synthesis
Plasmids containing the complementary DNA (cDNA) of h4F2hc (vector pSPORT) (15) and hLAT1 (pcDNA1/Amp-pSP64T) (11) were linearized using the restriction sites HindIII and EcoRV, respectively. cRNA was synthesized with T7 polymerase (Promega) according to standard protocols.
Expression in Xenopus laevis Oocytes
Oocytes were treated with collagenase A for 20–40 min at room temperature in Ca2+-free buffer containing NaCl (82.5 mmol/L), KCl (2 mmol/L), MgCl2 (1 mmol/L), and N-(2-hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) (HEPES; 10 mmol/L) at pH 7.4 and then kept at 16°C in ND96 buffer containing NaCl (96 mmol/L), KCl (2 mmol/L), MgCl2 (1 mmol/L), CaCl2 (1.8 mmol/L), and HEPES (5 mmol/L) at pH 7.4.
Oocytes were injected with a mixture of 5 ng of h4F2hc and 5 ng of hLAT1 cRNA or with 5 ng of h4F2hc cRNA alone dissolved in 50 nL of water. Transport assays were performed after 24 h of expression.
Amino Acids
Radioiodination of 1 mg of 2IT was performed by Cu1+-assisted nucleophilic exchange in acidic and reducing conditions at 100°C during 60 min, yielding >98% 125I-2-IT and a radiochemical purity of >98% (5). 3H-Tyrosine and 3H-phenylalanine were obtained from Sigma-Aldrich. 2IT, 2FT, and FET were obtained from ABX. IMT was produced as synthesized by Gulhke and Biersack (16).
Transport Assays
Time Course.
Amino acid uptake as a function of time was measured to determine the time window in which the transport rate is constant. Groups of 6–8 oocytes expressing both h4F2hc and hLAT1 or h4F2hc alone were washed 6 times in transport buffer (NaCl [100 mmol/L], KCl [2 mmol/L], MgCl2 [1 mmol/L], CaCl2 [1 mmol/L], and HEPES [10 mmol/L] at pH 7.4) and preincubated with transport buffer for 2 min at 26°C. The buffer was replaced by 0.1 mL of new transport buffer supplemented with 1 mmol/L of 2IT (with 125I-2-IT as a tracer) or with 1 mmol/L of tyrosine (with 3H-tyrosine as a tracer) and incubated for 0.5, 1, 3, 7, 15, 30, or 60 min at 26°C. Subsequently, the medium was removed rapidly and the cells were washed 6 times with ice-cold transport buffer and transferred into individual counting tubes containing 0.25 mL of 2% sodium dodecyl sulfate. The oocytes were lysed by shaking the tubes for 30 min. Radioactivity was determined by liquid scintillation counting.
Concentration Dependency.
Oocytes expressing h4F2hc-hLAT1 or only h4F2hc were incubated for 30 s at 26°C with varying concentrations of 2IT or tyrosine (0.01, 0.02, 0.05, 0.1, and 0.5 mmol/L) supplemented with the appropriate tracer. The cells were further processed as described for the time-course experiments. To estimate the apparent affinity for the independent experiments, curves were fitted to the data according to Michaelis–Menten using KaleidaGraph (version 3.51, Synergy Software).
Transstimulation of Influx.
Oocytes expressing h4F2hc-hLAT1 or only h4F2hc were injected with a preload of unlabeled amino acid (1 nmol of methionine or phenylalanine in 50 nL of H2O) 2 h before the transport assay and kept in ND96 buffer at 16°C. Control oocytes were injected with 50 nL of H2O alone. With an intraoocyte amino acid distribution volume of 400 nL, this leads to an intracellular concentration of ∼2.5 mmol/L of the preloaded amino acid (12). Prior to the transport assay, the cells were washed 6 times in transport buffer and preincubated for 2 min at 26°C. The buffer was replaced by a 2IT or phenylalanine concentration of 0.1 mmol/L, supplemented with the appropriate tracer and incubated for 30 s at 26°C. After the incubation, the medium was removed rapidly and the cells were further processed as described above. Results are expressed as the ratio between tracer uptake in amino acid–preinjected oocytes and tracer uptake in H2O-injected oocytes (controls).
Transstimulation of Efflux.
Because of the 1:1 obligatory exchange property of the hLAT1-h4F2hc transporter, it is possible to measure the uptake rate of an unlabeled substrate by measuring the efflux of an intracellularly preloaded labeled substrate. This is a particularly interesting methodology for the evaluation of tracers that are labeled with short-lived isotopes such as 18F. Oocytes expressing h4F2hc-hLAT1 or only h4F2hc were loaded with 50 nL of H2O containing 1 nmol of phenylalanine and 0.37 MBq of 3H-phenylalanine. The oocytes were then washed 6 times in ND96 buffer, transferred to individual tubes, and kept for 2 h in 0.2 mL of ND96 buffer at 16°C. Subsequently, 0.05 mL of buffer was removed to measure leakage. Oocytes were washed 6 times in transport buffer and preincubated for 2 min at 26°C. Next, the medium was replaced by 0.2 mL of fresh transport buffer (negative control) or 0.2 mL of transport buffer containing 0.1 mmol/L of unlabeled 2IT, IMT, FET, 2FT, or phenylalanine (positive control) and incubated for 90 s at 26°C. The radioactivity of the medium and the radioactivity of the oocyte were then counted separately.
Statistical Analysis
The data measured for oocytes expressing both h4F2hc and hLAT1 (the functional transporter) were corrected for endogenous background transport by subtraction of the data measured for oocytes expressing only h4F2hc. Experiments were repeated 3 times with different batches of oocytes. Values obtained for phenylalanine (reference) were used to normalize the data of the transstimulation experiments, in order to correct for variability between different batches of oocytes. All data are expressed as the mean ± SE unless otherwise stated.
RESULTS
Time Course and Concentration Dependency
Xenopus laevis oocytes expressing h4F2hc-hLAT1 rapidly accumulated the substrate 125I-2IT, whereas oocytes expressing h4F2hc alone showed only minor uptake (Fig. 1). As for tyrosine, the linear phase of uptake (near-constant transport rate) was limited to the first minutes of incubation. Therefore, we selected the shortest incubation technically possible for subsequent experiments: 30 s for uptake measurements and 90 s for efflux measurements.
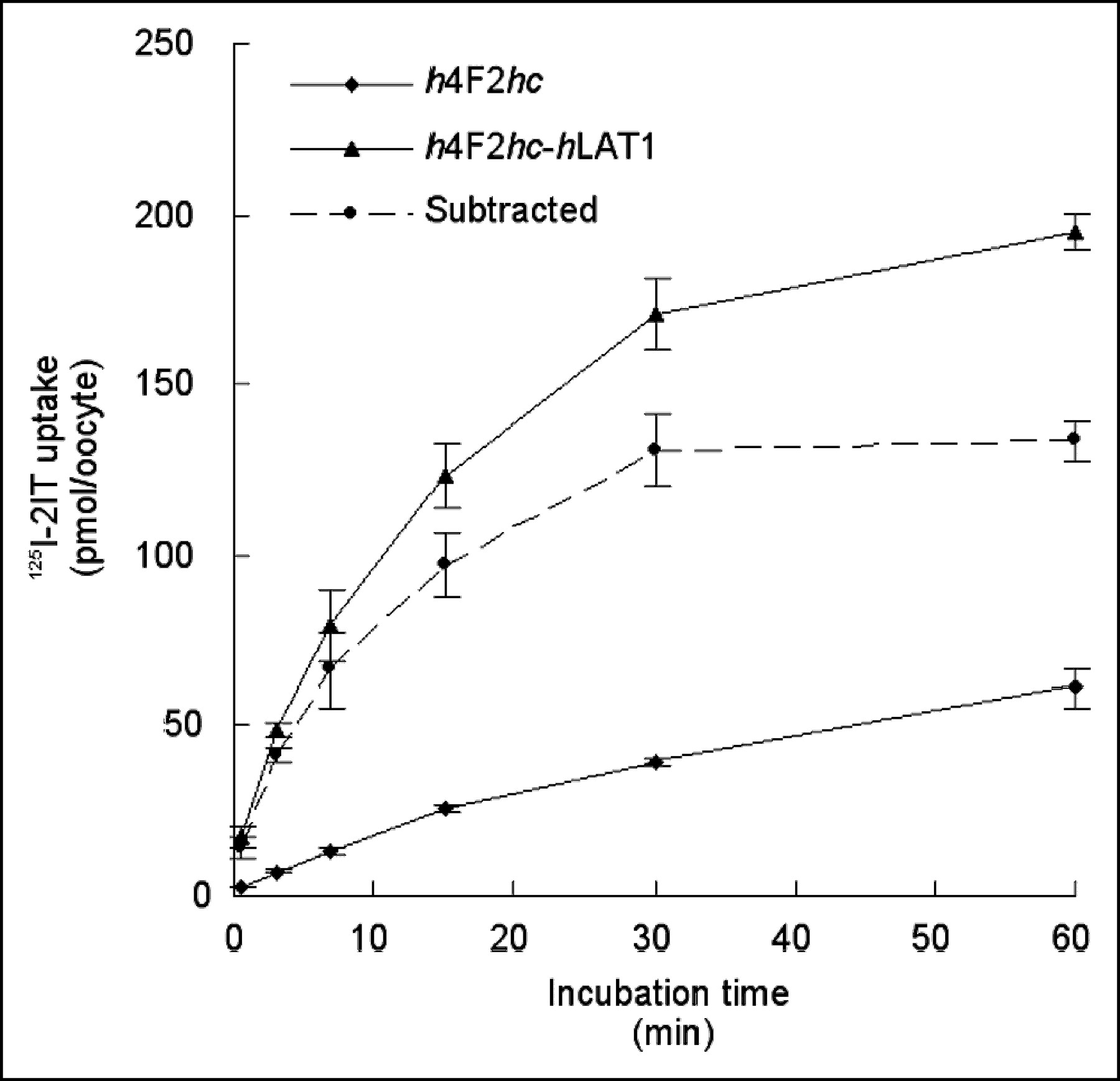
Time course of 125I-2IT (1 mmol/L) uptake in oocytes expressing h4F2hc-hLAT1 or only h4F2hc. Dashed line is subtracted curve: h4F2hc-hLAT1 minus h4F2hc, representing transport activity via LAT1 corrected for endogenous transport. Graph represents single experiment with 6–7 oocytes per datum point.
Incubation (30 s) of the oocytes expressing h4F2hc-hLAT1 with different concentrations of 2IT and tyrosine showed a saturable transport rate for both (Fig. 2). Curves corresponding to Michaelis–Menten kinetics were fitted to the data of the individual experiments (n = 3), and the mean derived apparent affinity (±SD) was 29.3 (±9.3) μmol/L for 2IT and 21.2 (±4.2) μmol/L for tyrosine. The mean Vmax (±SD) for 2IT uptake was equal to that for tyrosine uptake: 671 ± 81 pmol/oocyte/h and 672 ± 91 pmol/oocyte/h, respectively.
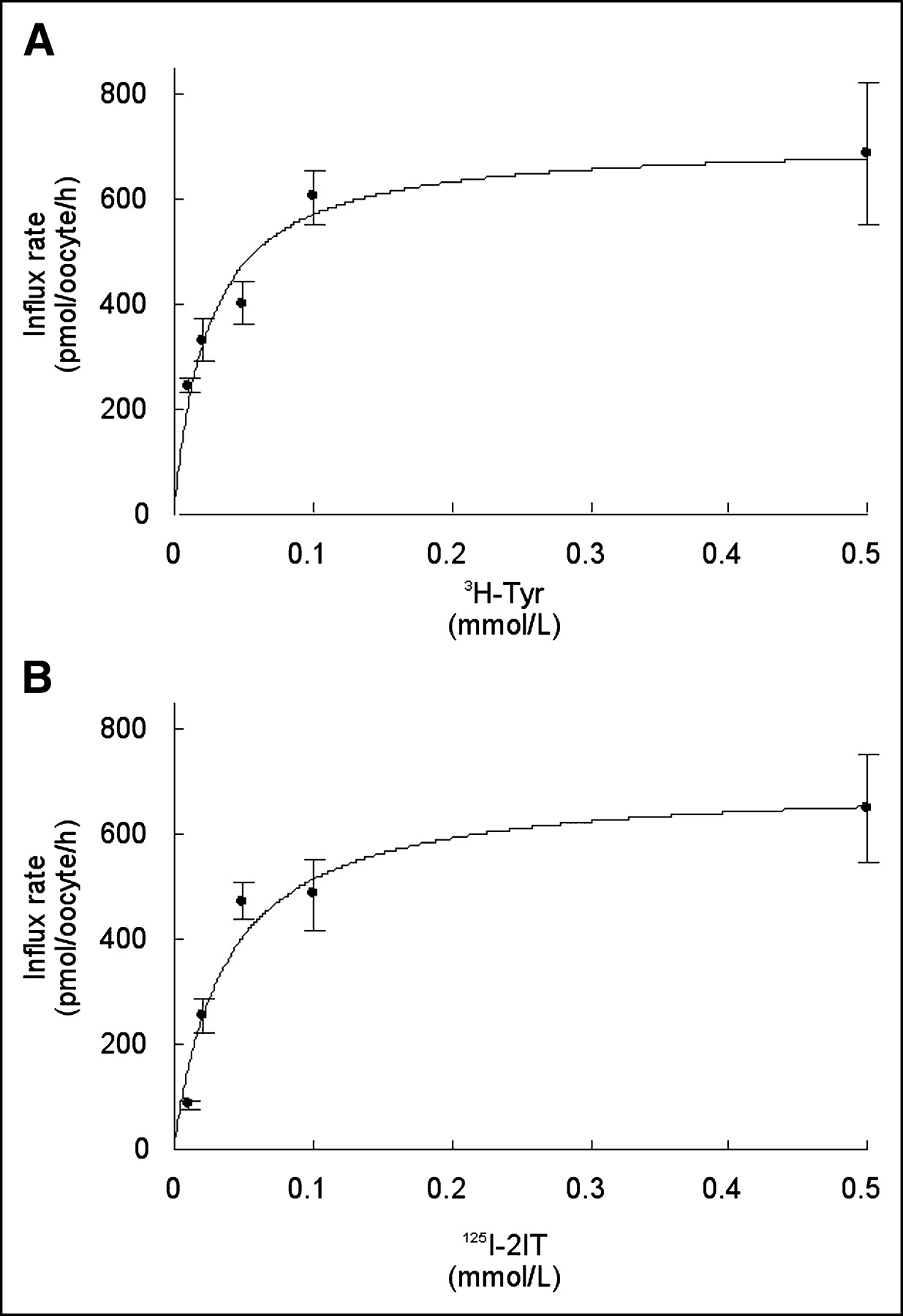
Concentration dependency of influx rate of tyrosine (A) and 2IT (B) via h4F2hc-hLAT1. Both curves show the hLAT1-mediated influx at different substrate concentrations (0.01–0.5 mmol/L). Graph represents single experiment with 5–8 oocytes per datum point. These experiments were repeated 3 times with different batches of oocytes. Tyr = tyrosine.
Transstimulation of 125I-2IT Influx by Preinjected Amino Acids
We measured a significant increase in the influx rate for 125I-2IT when the oocytes were preloaded with 1 nmol of unlabeled methionine or phenylalanine: 3.41 (±0.26)-fold for methionine preloading and 3.38 (±0.32)-fold for phenylalanine preloading (Fig. 3). The effect of the same preloading procedure on 3H-phenylalanine influx was slightly higher: 4.10 (±0.30)-fold for methionine preloading and 4.32 (±0.33)-fold for phenylalanine preloading.

Transstimulation of h4F2hc-hLAT1–dependent 125I-2IT and 3H-phenylalanine uptake by preloading the oocytes with 1 nmol of unlabeled phenylalanine or methionine. Values represent the ratio between influx rate into amino acid–preloaded oocytes and influx rate measured in H2O-injected oocytes (controls). Phe = phenylalanine.
Transstimulation of Phenylalanine Efflux by Extracellular Substrates
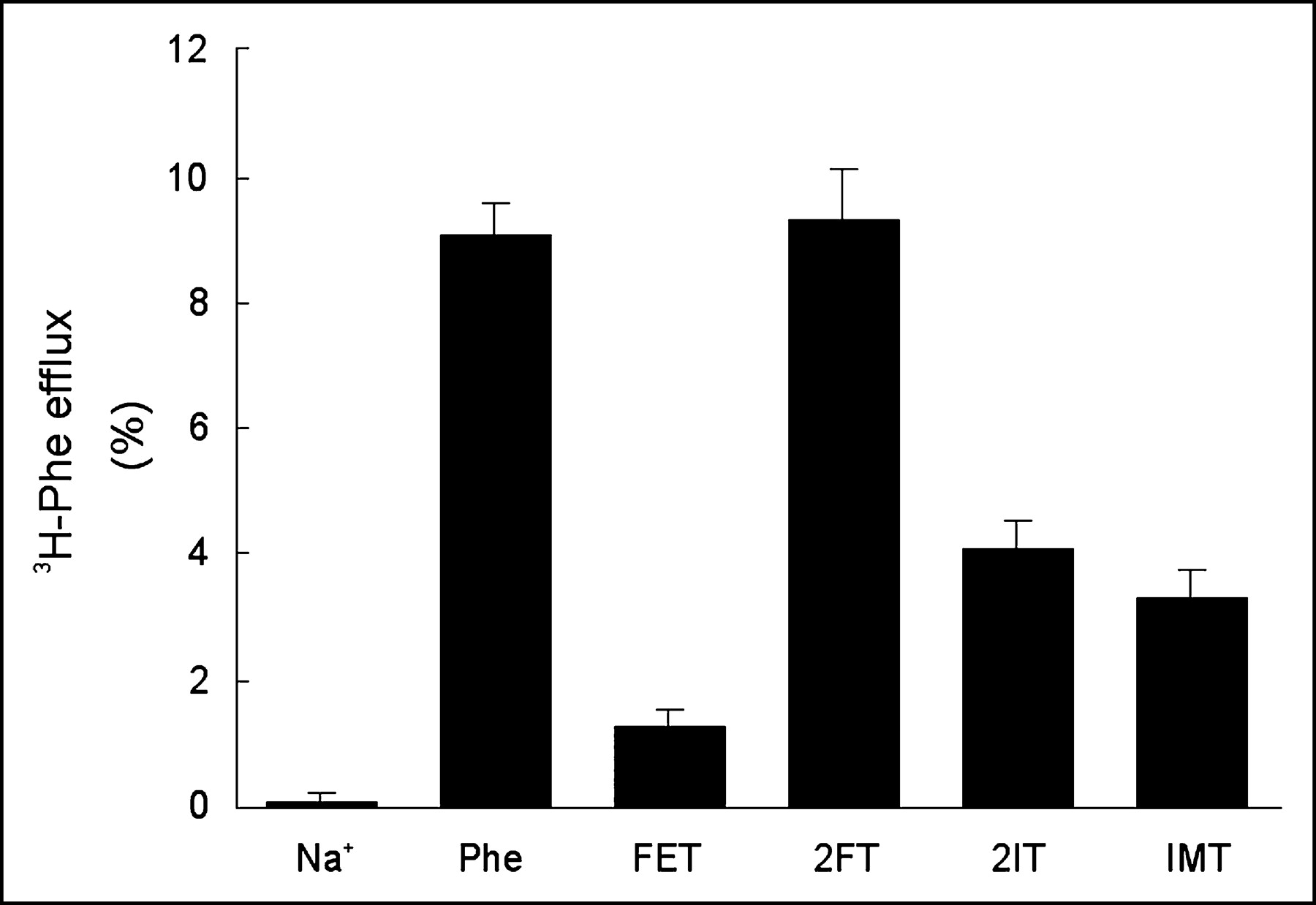
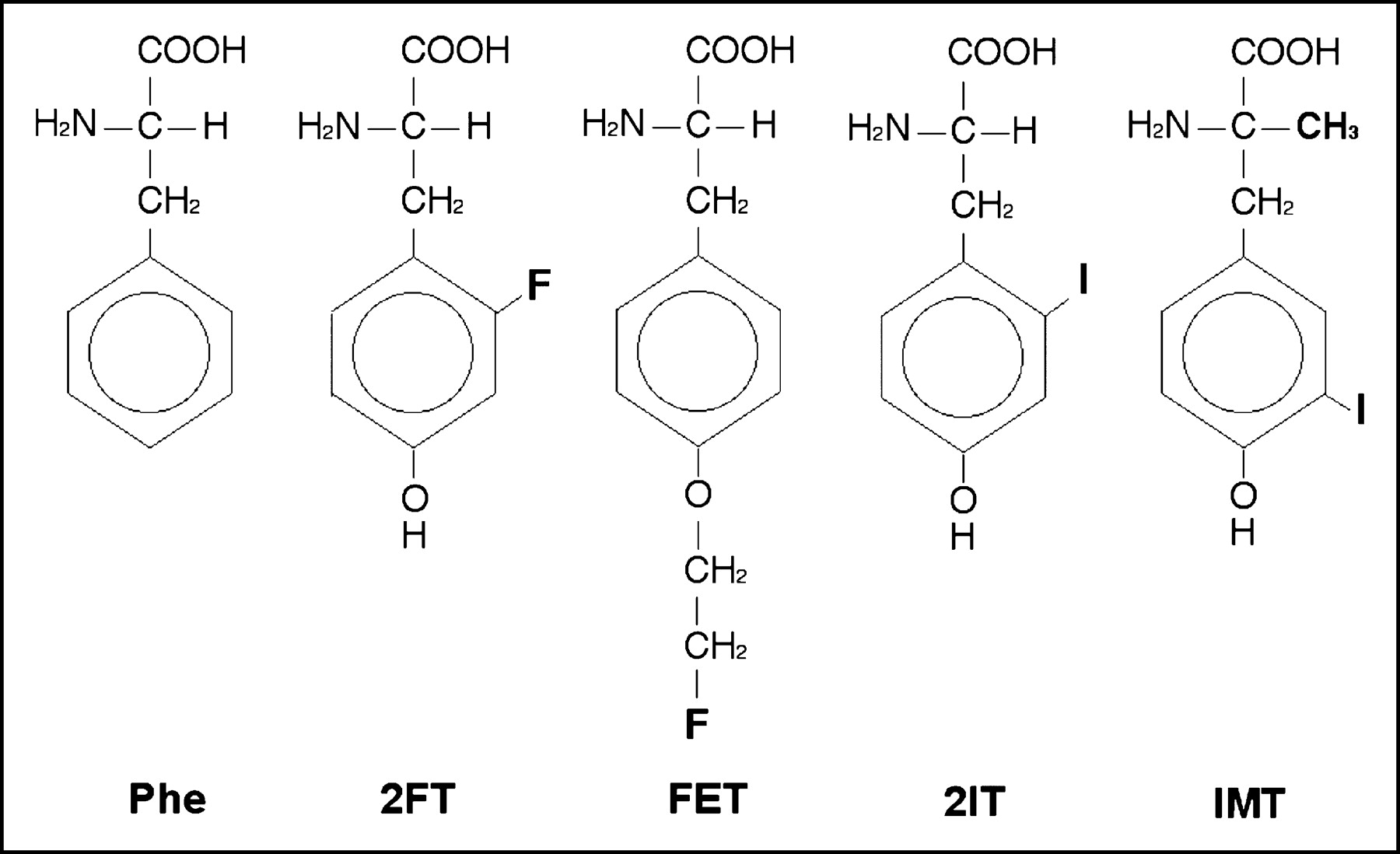
Nonlabeled reference products of the aromatic tracer amino acids for SPECT (2IT and IMT) and PET (2FT and FET) were given at a concentration of 0.1 mmol/L to measure their capacity to transstimulate the efflux of preloaded 3H-phenylalanine (Fig. 4). Their structure formulas are shown in Figure 5. There was no measurable amino acid leakage from the preinjected oocytes during the 2-h preincubation in ND96 buffer at 16°C or during the 90-s incubation in transport buffer lacking amino acids at 26°C (negative control). Addition of 0.1 mmol of phenylalanine per liter to the incubation medium induced the efflux of 9.1% ± 0.5% of the preinjected, labeled phenylalanine within 90 s (positive control). Compared with phenylalanine, an equimolar amount of 2FT had a similar effect on phenylalanine efflux (9.3% ± 0.8%), whereas FET induced only little efflux (1.3% ± 0.3%). Both iodinated compounds showed an intermediate ability to induce efflux: 4.1% ± 0.5% for 2IT and 3.3% ± 0.4% for IMT.
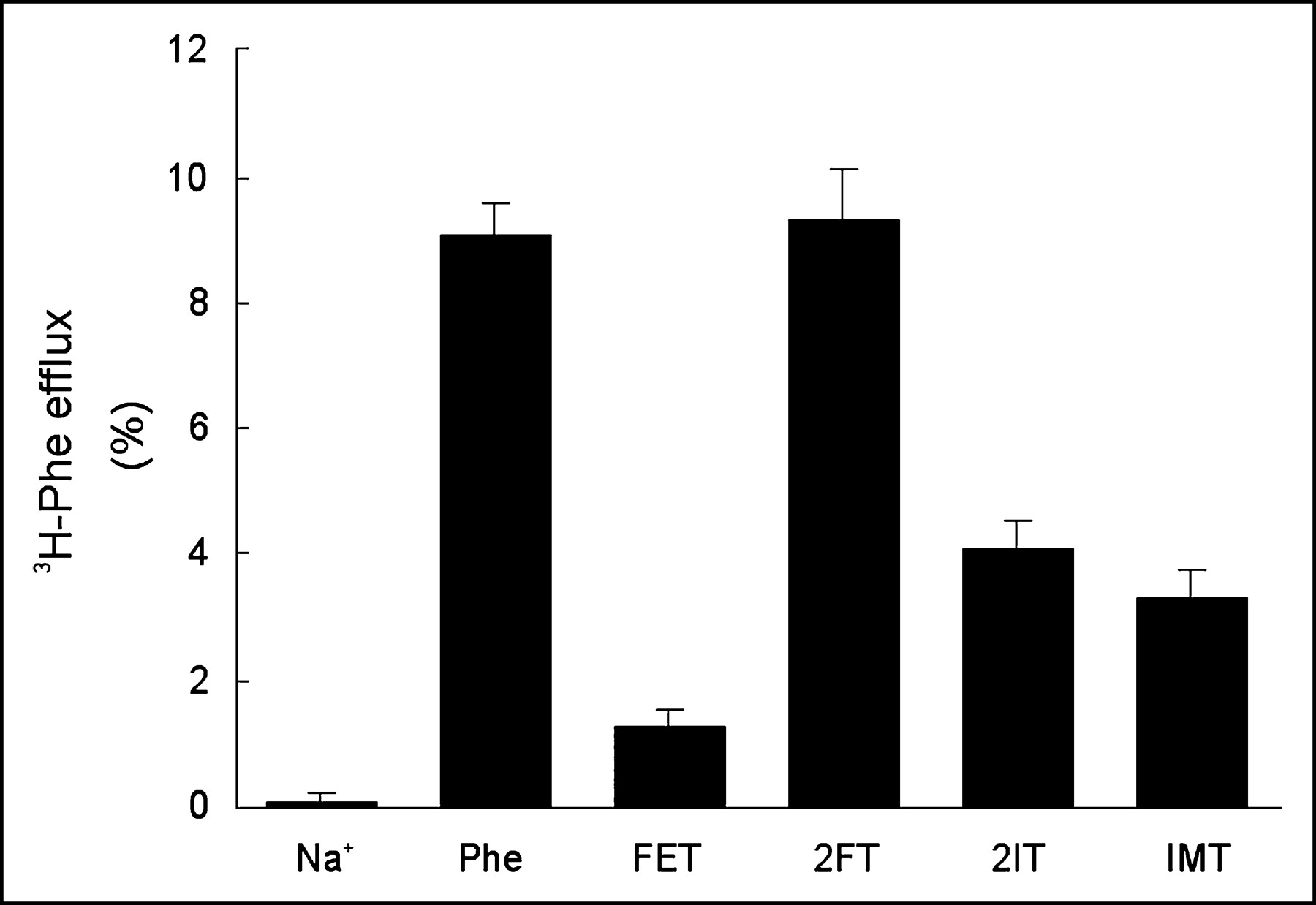
Transstimulation of h4F2hc-hLAT1–dependent 3H-phenylalanine efflux by a 0.1 mmol/L concentration of extracellular unlabeled tracer reference compounds (FET, 2FT, 2IT) or L-phenylalanine (positive control). Data represent the percentage of 3H-phenylalanine effluxed after 90 s of incubation. The result obtained in the absence of extracellular substrate represents the negative control. Phe = phenylalanine.

Chemical structure of the different amino acid analogs used for the efflux assays. Phe = phenylalanine.
DISCUSSION
We have studied the transport of the amino acid SPECT tracer 125I-2IT by the h4F2hc-hLAT1 transporter using the Xenopus laevis oocyte expression system. This allowed us to study the uptake of tracers by a single transporter and, by measuring the efflux of preinjected amino acids, to quantify the influx of unlabeled amino acids.
Cell culture studies have shown that the different aromatic amino acid tracers used for SPECT and PET are largely accumulated through BCH-inhibitable sodium-independent transport, suggesting the involvement of an L-type transporter (5–7,17,18). Currently, 3 transporters have been identified that exhibit L-type transport: h4F2hc-hLAT1, h4F2hc-hLAT2, and hLAT3 (8–10). The first 2 are heterodimers composed of a catalytic subunit (light chain: hLAT1 or hLAT2) covalently linked to a glycoprotein (heavy chain: h4F2hc). Both subunits are necessary for functional expression. These heterodimeric transporters exhibit an obligatory exchange mechanism, meaning that for each amino acid taken up, they also transport one out. The hLAT3 transporter was only recently cloned and appears to be distinct from the first 2 subtypes: hLAT3 does not require a glycoprotein for functional expression and allows both influx and efflux of substrates through facilitated diffusion without the obligatory exchange property. Another difference is that hLAT3 has millimolar affinity (low affinity) for its substrates, whereas h4F2hc-hLAT1 and h4F2hc-hLAT2 show micromolar affinities (high affinity) for their substrates. Northern blot analyses showed a high expression of h4F2hc-hLAT1 in a wide variety of cancer cell types. In contrast, h4F2hc-hLAT2 is found mostly on the basolateral membrane of transporting epithelia and is believed to function predominantly as an efflux pathway (1,19). We therefore hypothesize that the in vivo tumor uptake of aromatic amino acid tracers found on SPECT and PET scans reflects predominantly the high-affinity LAT1 pathway. This hypothesis is supported by a recent study on isoform selectivity showing that the rate of 123I-IMT uptake is almost 6-fold higher when mediated by h4F2hc-hLAT1 than when mediated by h4F2hc-hLAT2 (20,21). However, the potential role of the newly discovered LAT3 transporter (low affinity) in the cancer cell accumulation of aromatic amino acid SPECT and PET tracers needs to be addressed in future studies.
Our data show that the affinity of 125I-2IT for h4F2hc-hLAT1 is comparable to that of tyrosine, suggesting that binding of 125I-2IT by the transporter protein is not significantly hampered by the bulky iodine atom. However, high-affinity interaction does not necessarily lead to efficient transport. For instance, melphalan, an anticancer drug, and the thyroid hormones that interact with high affinity with h4F2hc-hLAT1 are transported at a relatively slow rate (14). These findings suggest that the structural modifications of these amino acid analogs interfere with the conformational changes required for the translocation of the substrates. The results from the present efflux assay are compatible with such a possibility in the case of 2IT, which is transported at a lower rate than 2FT, which has a relatively small fluorine atom in the ortho position instead of the larger iodine. FET carrying a fluoroethyl group in the para position is transported at the lowest rate. The 2 SPECT tracers 2IT and IMT show a comparable intermediate transport rate.
Our data suggest that 2FT is the best tracer for imaging LAT1 transport. Human tumors expressing LAT1 should transport 2FT at a high rate. However, the experimental setting using Xenopus oocytes does not fully reflect the situation of in vivo tumor imaging in humans. Potential confounding factors are rapid efflux of the tracer from the tumor (leading to false-negative findings) and accumulation in inflammatory tissues (leading to false-positive findings). These issues can be addressed using an in vivo animal model.
The obligatory exchange function of LAT1 implies that uptake of an extracellular substrate needs to be coupled to extrusion of an intracellular substrate. The net influx of aromatic amino acids therefore depends on the availability of intracellular exchange substrates. Such intracellular substrates might enter the cell by parallel, unidirectional transporters with an overlapping substrate selectivity. Methionine has been suggested to be such a recycling substrate, since it is a poor uptake substrate but a relatively good efflux substrate for LAT1 (1,13). Once extruded, it is taken up again in the cell by the unidirectional system A, for which it has a high affinity. In our study, the influx rate of 125I-2IT was strongly transstimulated by methionine preloading. This characteristic is in accordance with the observation that the intracellular amino acid concentration is rate limiting for amino acid accumulation via LAT1 (12). Induction of the recycling mechanism by stimulation of system A–mediated methionine accumulation could provide a way to increase global LAT1-mediated tracer accumulation into tumors (22,23).
The rationale for using amino acid tracers for the metabolic imaging of cancers relies on the fact that cancer cells need to transport large amounts of nutrients in order to sustain the growth of the tumor. Animal studies have shown high tumor uptake and negligible uptake in inflammatory lesions of the aromatic amino acid tracers 18F-FET and 123I-2IT (24,25). Therefore, these tracers might provide a more tumor-specific imaging modality complementary to 18F-FDG PET.
Aside from tumor detection based on increased demand for metabolic substrates, the h4F2hc-hLAT1–transported tracers could also provide information about tumor heterogeneity: Several cancer cell lines have been reported to have defective h4F2hc expression but conservation of hLAT1 expression (2). Because hLAT1 needs h4F2hc for functional expression, it is expected that tumors lacking h4F2hc expression will not intensely accumulate the aromatic amino acid tracers. Similar observations have been reported for a rat colon cancer model with multiple liver metastasis, in which h4F2hc expression was defective in several of the liver metastasis (4). Lesions with defective h4F2hc expression were significantly smaller than lesions with a functional h4F2hc-hLAT1 transporter, consistent with the suggestion that functional system L expression is closely related to tumor growth. Therefore, we expect that the aromatic amino acid tracers will reveal only the lesions with conserved h4F2hc-hLAT1 expression and thus with the biggest growth potential. This phenomenon might explain the heterogeneity of 18F-2FT tumor accumulation observed in a recent clinical study by Hustinx et al., who reported lymphoma patients with coexisting 18F-FDG–positive/18F-2FT–negative and 18F-FDG–positive/18F-2FT–positive lesions (26).
CONCLUSION
125I-2IT has a high affinity for LAT1, comparable to that of natural tyrosine. The influx rate of 2IT is comparable to that of IMT and is amplified when the availability of the intracellular exchange substrate (for instance, methionine) is increased. The influx rate of 2FT is comparable to that of phenylalanine, whereas FET is only poorly transported.
The molecular characterization of the interaction of 2IT tracer with the h4F2-hLAT1hc transporter presented here provides further insight into the mechanism and regulation of the in vivo tumor accumulation of this amino acid tracer.
Acknowledgments
This work was supported by a grant from the Medical Foundation Horlait-Dapsens to one of the authors. The laboratory of another of the authors is supported by a grant of the Swiss National Science Foundation.
Footnotes
Received Dec. 26, 2003; revision accepted Apr. 12, 2004.
For correspondence or reprints contact: Tony Lahoutte, MD, Department of Nuclear Medicine, Academic Hospital Free University Brussels (AZ-VUB), Laarbeeklaan 101, 1090 Jette, Belgium.
E-mail: tony.lahoutte{at}az.vub.ac.be
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Comparison of the Amino Acid Tracers 18F-FET and 18F-DOPA in High-Grade Glioma Patients
- Transport of 3-Fluoro-L-{alpha}-Methyl-Tyrosine by Tumor-Upregulated L-Type Amino Acid Transporter 1: A Cause of the Tumor Uptake in PET
- Changes in Tumor Metabolism as Readout for Mammalian Target of Rapamycin Kinase Inhibition by Rapamycin in Glioblastoma
- Tumor Cell Metabolism Imaging
- 6-L-18F-Fluorodihydroxyphenylalanine PET in Neuroendocrine Tumors: Basic Aspects and Emerging Clinical Applications
- L-Type Amino Acid Transporters LAT1 and LAT4 in Cancer: Uptake of 3-O-Methyl-6- 18F-Fluoro-L-Dopa in Human Adenocarcinoma and Squamous Cell Carcinoma In Vitro and In Vivo
- 18F-FET PET Compared with 18F-FDG PET and CT in Patients with Head and Neck Cancer
- PET with O-(2-18F-Fluoroethyl)-L-Tyrosine in Peripheral Tumors: First Clinical Results
- Molecular Transport Mechanisms of Radiolabeled Amino Acids for PET and SPECT