


Abstract
The human somatostatin receptor subtype 2 (hSSTr2)–68Ga-DOTATOC reporter system has several attractive features for potential translation to human studies. These include a low expression of hSSTr2 in most organs, a rapid internalized accumulation of 68Ga-DOTATOC in the SSTr2-expressing cells, and a rapid excretion of unbound radioligand by the renal system. We performed a series of in vitro and in vivo validation studies of this reporter system. Methods: A retroviral vector containing a dual reporter, pQCXhSSTr2-IRES-GFP (IRES: internal ribosome entry site; GFP: green fluorescent protein), was constructed and transduced into Jurkat, C6, and U87 cells. Stably transduced reporter cells were characterized in vitro using optical and radiometric methods. Multiple tumor-bearing mice were evaluated with 68Ga-DOTATOC PET studies. Results: The dual-reporter genes were incorporated into all tumor cell lines, and their expression levels were confirmed by fluorescence-activated cell sorting (FACS), GFP visualization, and reverse-transcriptase polymerase chain reaction (RT-PCR) analysis for hSSTr2. In vitro, hSSTr2 cell membrane expression was 36,000, 280,000, and 1,250,000 copies per cell for the SSTR2-transfected Jurkat, U87, and C6 cell lines. Small-animal PET of 68Ga-DOTATOC in tumor-bearing mice demonstrated that the in vivo uptake of this radioligand was directly proportional to the in vitro expression of hSSTr2. The in vivo uptake of 68Ga-DOTATOC, at 2 h after injection, was low in all organs except the kidneys (7.8 percentage of injected dose per gram [%ID/g]) and as high as 15.2 %ID/g in transduced C6 tumors. The corresponding transduced–to–nontransduced tumor uptake ratio was 64, and the tumor-to-muscle uptake ratio was around 500. Conclusion: 68Ga-DOTATOC is an excellent specific ligand for this hSSTr2 reporter system and for hSSTr2 reporter gene PET. Because DOTATOC has undergone extensive clinical testing, this human reporter system has the potential for translation to human studies.
Somatostatin receptors (SSTrs) belong to a 7–transmembrane domain family of G-protein–coupled receptors. All 5 receptor subtypes are highly homologous between mice and humans and are expressed at relatively low levels in most organs. They are moderately expressed in the brain, gastrointestinal tract, pancreas, kidneys, and spleen. In contrast, SSTrs, especially SSTr2 (somatostatin receptor subtype 2), have been shown to be highly expressed in various human tumors, including pituitary, endocrine, pancreatic, and small cell lung tumors as well as in carcinoid tumors, paraganglioma, medullary thyroid carcinoma, and pheochromocytoma (1). These observations have led to radionuclide-based imaging and therapy of SSTr2-expressing tumors (2,3).
Somatostatin and its numerous synthetic derivatives have a wide spectrum of physiologic functions. These ligands act as neurotransmitters, paracrine or autocrine regulators, or systemic regulators of cell secretion, neuromodulation, smooth muscle contractility, nutrient absorption, and cell growth (4). In tumors, somatostatin and somatostatin agonists indirectly control cell growth in vivo through inhibition of the release of growth-promoting hormones (5). Somatostatin can also act through antiangiogenic mechanisms (6) involving the inhibition of proliferation and migration of endothelial cells and monocytes (7) or by interfering with the production of proangiogenic factors from tumor cells (8). Moreover, direct antiproliferative activity in both normal and tumor cells, in which one of the main intracellular pathways responsible was the modulation of phosphotyrosine phosphatase, has also been reported (9).
Octreotide is a metabolically stable somatostatin inhibitory peptide, originally developed for the symptomatic treatment of patients with metastatic carcinoid tumors. Octreotide binds with a low nanomolar affinity to hSSTr2 and hSSTr5, with moderate affinity to hSSTr3 (200 nM), and with poor affinity to hSSTr1 and hSSTr4 (>1,000 nM) (10). 111In-diethylenetriaminepentaacetic acid (DTPA)-octreotide was subsequently developed as an hSSTr imaging agent and is approved by the Food and Drug Administration (FDA) for SPECT of neuroendocrine tumors (2).
DOTATOC was subsequently developed for labeling with 90Y or 177Lu and was initially used for targeted radiotherapy. Clinical trials (phase II/III) have been performed in many European centers (3). DOTATOC has also gained widespread use as a PET agent when chelated with 68Ga (11,12). In addition, other PET nuclides, including 94mTc, 18F, 86Y, and 64Cu, have been coupled to octreotide or similar somatostatin analogs for quantitative PET of hSSTr2 expression. The preclinical or clinical validation of these radiolabeled somatostatin derivatives, some of which are currently used in clinical studies, indicates that the hSSTr2–radiolabeled ligand combination could be used for transgene imaging in patients.
The hSSTr2 gene has been incorporated into a replication-incompetent adenoviral vector (Ad-CMVhSSTr2) and used as a radionuclide-based reporter gene for noninvasive imaging of xenografts (13). However, the radioligands (FDA-approved 111In-DTPA-octreotide (14,15), 99mTc-labeled P829 (16), and 94mTc-demotate 1 (17)) that have been used to image hSSTr2 expression target several SSTrs and produce comparatively poor-quality images. The relatively high background radioactivity and moderate transduced–to–nontransduced tumor uptake ratios have limited the clinical translation of hSSTr2 reporter gene imaging since the original reports in 1999 (14).
To further investigate hSSTr2 as a potential reporter gene of human origin, we constructed a constitutively expressing dual-reporter retroviral vector (pQCXhSSTr2-IRES-GFP; IRES: internal ribosome entry site, GFP: green fluorescent protein), transduced several cell lines with this reporter, and chose 67/68Ga-DOTATOC as the radioligand for in vitro validation and in vivo imaging. We chose 67/68Ga-DOTATOC because of its high and selective affinity for the hSSTr2 receptor and because of the high-quality, low-background PET images that have been obtained in patients with neuroendocrine tumors. The focus of the current study was to investigate the feasibility of imaging hSSTr2 reporter gene expression with 68Ga-DOTATOC in mice to obtain preclinical data in support of future studies in patients.
MATERIALS AND METHODS
General
All chemicals were obtained from commercial sources and used without further purification. DOTATOC and gallium-DOTATOC were synthesized as reported earlier (18). 67GaCl3 was purchased from MDS Nordion, and 68GaCl3 was eluted from a commercial 68Ge/68Ga generator (Cyclotron Co Ltd.). U87, C6, and Jurkat cell lines were obtained from American Type Culture Collection and propagated as recommended. Chinese hamster ovary (CHO) cells selectively transfected with hSSTr1, hSSTr2, hSSTr3, and hSSTr4 were kindly provided by Dr. Friedrich Raulf of Novartis. Radioactivity was quantified with a Wizard 3″ 1480 γ-counter (PerkinElmer) or a dose calibrator (CRC-30BC; Capintec).
Plasmid Construction and Cell Transduction
The pQCXhSSTr2-IRES-GFP retroviral vector was developed using an approach similar to that described for the pQCXhNET-IRES-GFP retroviral vector (19). Briefly, to construct the pQCXhSSTr2-IRES-GFP vector, cDNA for SSTr2 was amplified from the construct kindly provided by Dr. Graeme Bell (University of Chicago). The resulting polymerase chain reaction (PCR) product was used for ligation into the pQCX-IRES-GFP backbone to obtain the final plasmid where the gene SSTr2 was separated from the GFP by an IRES element.
U87, C6, and Jurkat cell lines were transduced with the pQCXhSSTr2-IRES-GFP retroviral vector. Briefly, the DNA of vector-encoding plasmid (10 μg) was transfected into the GPG29 packaging cell line using LipofectAMINE 2000 (Invitrogen). All cell lines were stably transduced by incubating 50% confluent cell cultures with a virus-containing medium for 12 h in the presence of polybrene (8 mg/mL; Sigma). Cells were sorted using a fluorescence-activated cell sorter (BD Bioscience).
Semiquantitative Reverse-Transcriptase PCR Analysis
Total RNA was isolated using TRIzol reagent (Invitrogen) and treated with RNase-free DNase I (AmBion) according to the manufacturer's instructions. The following primer sets were used for cDNA synthesis (Promega):
hSSTr2 gene: (F) 5′-GTCACCCGAATGGTGTCCATC-3′;
(R) 5′-ATTTGTCCTGCTTACTGTCACTC-3′;
GFP gene: (F) 5′-CCCTCGTGACCACCCTGACCT-3′;
(R) 5′-CCATGTGATCGCGCTTCTCGT-3′;
β-actin gene: (F) 5′-TGGTGGGCATGGGTCAGAAGGAT-3′;
(R) 5′-CTAGAAGCATTTGCGGTGGACGAT-3′.
68/67Ga-DOTATOC Radiosynthesis
68Ga-DOTATOC was synthesized by mixing 200 μL of the 68Ga/68Ge generator eluate (0.1 M HCl) with 10 μL of DOTATOC (10 nmol) and 40 μL of 1.0 M NH4OAc (pH 3.8) for 10–12 min at 95°C. The radioligand was purified from unchelated 68Ga using a SepPak C18 cartridge (Waters) preconditioned with 5 mL of ethanol and 10 mL of H2O. Unchelated 68Ga and NH4OAc buffer were eluted with 5 mL of H2O, and 68Ga-DOTATOC was eluted with 3 mL of ethanol. Final quality control of the radioligand was performed by high-performance liquid chromatography using a Jupiter C18 column (Phenomenex) with an eluent of 0.05% trifluoroacetic acid/MeCN gradient (0–15 min, 9.5%–47.5% MeCN; 16–19 min, 95% MeCN; and 19–20 min, 9.5% MeCN). 67Ga-DOTATOC was prepared similarly.
Uptake and Dissociation of 67Ga-DOTATOC in hSSTr2-Transduced Cells
Uptake Studies.
Multiple samples containing approximately 1.0 kBq of 67Ga-DOTATOC (1.6 pmol) were incubated with 1.0 × 106 cells suspended in a total volume of 0.5 mL of phosphate-buffered saline (1.0 mM Ca(II), 0.5 mM Mg(II)). The samples were gently shaken at 24°C or 37°C for between 5 min and 4 h. At various time points, triplicate samples were isolated by rapid filtration through a glass microfiber filter (934-AH; Whatman) and washed with 3 × 2 mL of ice-cold tris-buffered saline (pH 7.4) and assayed for radioactivity.
Dissociation Studies.
hSSTr2-transduced cells were incubated as described in the previous section for either 0.25 or 1 h before a 300-fold excess of gallium-DOTATOC (0.5 nmol) was added. At various times, the cells were isolated and the radioactivity measured as described in the previous section.
The resultant uptake data were expressed as 67Ga-DOTATOC molecules per cell and plotted as a function of time.
Binding Affinity of natGa (Gallium)-DOTATOC and hSSTr2 Expression Level
Saturation binding studies were performed using 67Ga-DOTATOC and varied concentrations of gallium-DOTATOC. Briefly, triplicate samples containing 1.0 × 106 cells and 0.1–500 pmol of 67/natGa-DOTATOC (∼1 kBq) in 0.5 mL of 1% bovine serum albumin/phosphate-buffered saline (1.0 mM Ca(II) and 0.5 mM Mg(II), pH 7.4) were incubated at 37°C (C6-SSTr2) or room temperature. After an incubation of 60 min, the cells were isolated by rapid filtration through glass microfiber filters and washed with 3 × 2 mL of ice-cold tris-buffered saline (pH 7.4). The cell uptake was calculated as molecules per cell. The data were plotted as a function of gallium-DOTATOC concentration, and the dissociation constant (Kd) and maximum number of binding sites (Bmax) per cell were estimated using a least-squares fitting routine (GraphPad Prism 5; GraphPad Software).
hSSTr2-Mediated 67Ga-DOTATOC Uptake in hSSTr-Expressed or Wild-Type (WT) Cells
The specific accumulation of 67Ga-DOTATOC was studied (with an approach similar to the one used for the uptake and dissociation studies) using hSSTr-transduced cells and corresponding WT cells. The incubation was 2 h at 37°C. One nanomole of gallium-DOTATOC or somatostatin-14 was used for blocking.
Small-Animal PET 68Ga-DOTATOC Imaging
All animal experiments were approved by the Institutional Animal Care and Use Committee of Memorial Sloan-Kettering Cancer Center. Male athymic nu/nu mice (age, 8 wk; Harlan) were pretreated with 4.5 Gy of external irradiation 24 h before subcutaneous implantation of tumor cells. Cells were suspended in 200 μL of cell culture medium or, in the case of Jurkat cells, 200 μL of 50% Matrigel (BD Bioscience). One group of animals was injected with 8 × 106 C6 cells and 8 × 106 U87 (both WT and SSTr2-transduced). A second group of animals received 15 × 106 Jurkat cells (WT and SSTr2-transduced), followed 3 wk later by 8 × 106 C6-SSTr2 and 8 × 106 U87-SSTr2 cells. Nine to 14 d after the final inoculation, small-animal PET and tissue sampling were performed.
For the small-animal PET and biodistribution studies, 68Ga-DOTATOC (3.7–11.4 MBq, 1.0 nmol) was injected into the penile vein of the anesthetized mouse (2% isoflurane). Small-animal PET was performed at 1 and 2 h after injection on an R4 microPET scanner (Concorde Microsystems), with the tumors centered in the field of view. Ten-minute scans were acquired, with an energy window of 250–750 keV and a coincidence timing window of 6 ns. Region-of-interest analysis of the acquired images was performed using ASIPro software (Siemens), and the observed maximum pixel value was corrected for partial-volume effects based on the size of the tumor.
Directly after the last small-animal PET study, the mice were euthanized by CO2 inhalation, and the organs of interest were removed, weighed, and counted in a γ-counter, along with a known volume of the injectate. Data were expressed as %ID/g.
In Vivo Fluorescence Imaging
Tumor-bearing mice were anesthetized with 2% isoflurane and imaged on a Maestro imager (Cambridge Research Instruments) running Maestro Imaging Software (version 2.4), as described previously (20). GFP images were acquired at 445- to 490-nm excitation and using a 515-nm long-pass emission filter at f/11 for 300 ms. For spectral deconvolution, an image matrix was collected on the Maestro, with excitation at 445–490 nm, and 110 emission images were acquired in 2-nm steps between 500 and 720. Fluorescence region and autofluorescence regions were identified and spectrally unmixed using the instrument's software.
Statistical Analysis
The statistical significance of differences between mean values was determined using the Student t test. P values less than 0.05 were considered to be statistically significant.
RESULTS
Generation of hSSTr2-GFP Reporter Cells
A retroviral vector (pQCXhSSTr2-IRES-GFP) encoding both hSSTr2 and GFP genes was constructed (Supplemental Fig. 1A; supplemental materials are available online only at http://jnm.snmjournals.org) and used to transduce Jurkat, U87, and C6 cells. Stably transduced cell lines were obtained after FACS-sorting based on GFP expression (Supplemental Figs. 1B and 1D). U87- and C6-transduced cells expressed a significantly higher level of GFP expression than did Jurkat cells. After FACS purification, more than 95% of C6 and U87 cells were GFP-positive, whereas only 80% of Jurkat cells expressed GFP (Supplemental Fig. 1B). hSSTr2 and GFP expression was confirmed by RT-PCR (Supplemental Fig. 1C) and fluorescence microscopy (Supplemental Fig. 1D).
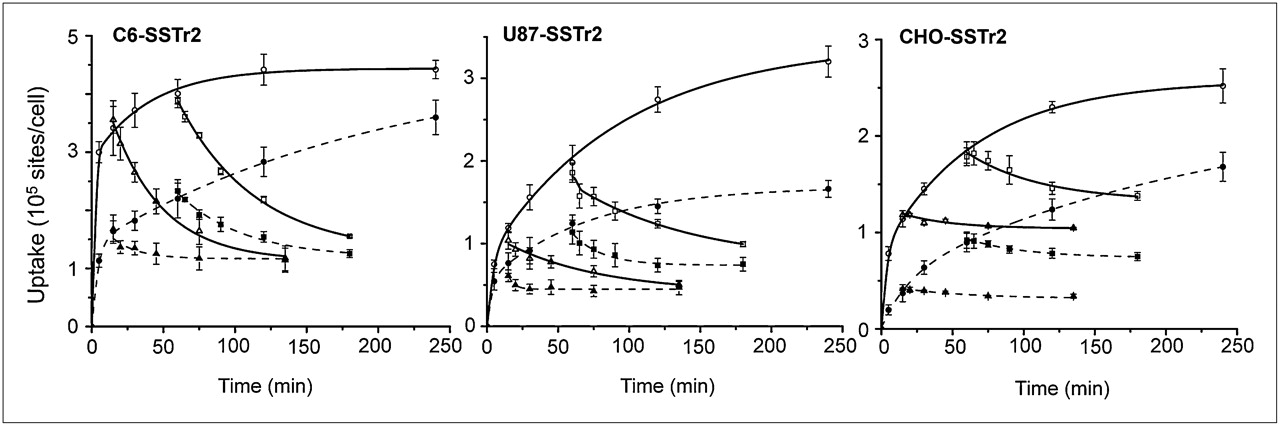
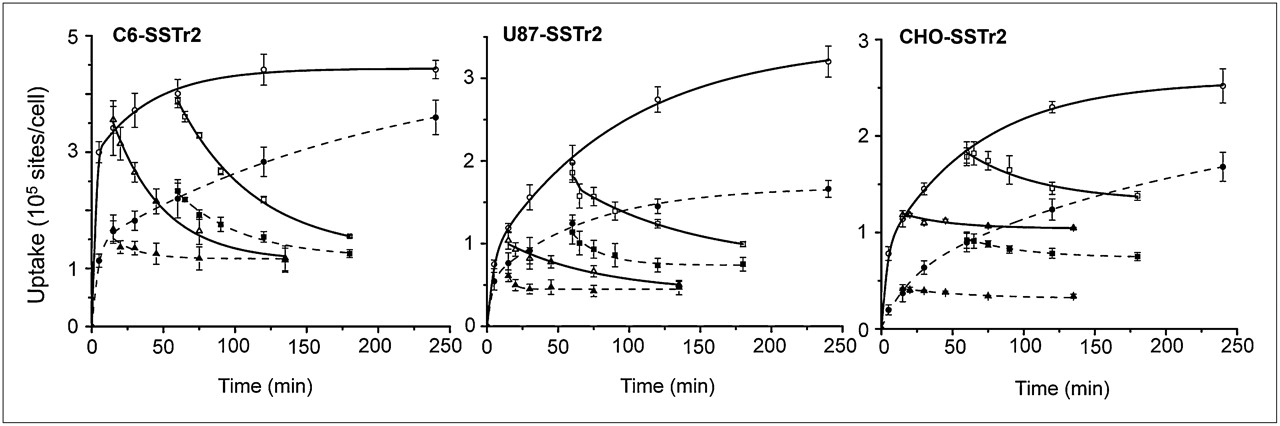
Kinetics of 67Ga-DOTATOC binding to and dissociation from hSSTr2 expressed by transduced cells. Dissociation of ligand–receptor complex was initiated by addition of excess natGa-DOTATOC after 0.25 and 1 h. Results (mean ± SD) are from 2–4 independent studies. Solid line = 37°C; dotted line = 24°C.
68/67Ga-DOTATOC Radiosynthesis
The 68Ga-DOTATOC was synthesized within 30 min with a specific activity of approximately 19 GBq/μmol (60% radiochemical yield, uncorrected). 67Ga-DOTATOC was prepared using a similar method, with radiochemical yields of 98% and a specific activity of more than 9 GBq/μmol.
Uptake and Dissociation of 67Ga-DOTATOC in hSSTr2-Transduced Cells
67Ga-DOTATOC showed a specific time- and temperature-dependent uptake in the hSSTr2-tranduced cells (C6, U87, and CHO) (Fig. 1). The uptake in all 3 cell lines at 37°C was significantly higher than that at 24°C (P < 0.022) after a 4-h incubation. 67Ga-DOTATOC displayed a rapid uptake in the C6-SSTr2 cells at 37°C and reached an apparent equilibrium within 30 min. The kinetic displacement studies showed a similar displacement of radioactivity from cells preincubated for 0.25 and 1 h, indicating that radiolabeled DOTATOC was not internalized at 37°C. The same study performed at 24°C showed that equilibrium binding was not reached within 4 h of incubation and showed similar low levels of undisplaced cell-bound radioactivity.
Interestingly, the kinetic binding studies with U87-SSTr2 cells showed more rapid equilibrium when performed at 24°C than at 37°C, consistent with the displacement studies performed at 37°C. This finding suggests that a large fraction of radiolabeled DOTATOC was being internalized.
The kinetic binding studies with CHO-SSTr2 cells showed similar rates of equilibrium binding whether performed at 24°C or 37°C. However, the displacement studies showed considerably higher levels of DOTATOC internalization at 37°C.
hSSTr2 Expression Level and Binding Affinity of natGa-DOTATOC
On the basis of the kinetic profiles (Fig. 1), the experimental conditions were optimized to determine the Kd value and SSTr2 expression level (Bmax) in each of the transduced cell lines. These data are listed in Table 1. A typical saturation binding curve and Scatchard transformation plot for each transduced cell line is shown in Supplemental Figure 2. The Kd values for all 4 SSTr2-transduced cell lines were similar, approximately 5–7 nM. The Scatchard transformations were all linear, indicating a single class of high-affinity binding sites for 67Ga-DOTATOC. The rank order for hSSTr2 expression was C6-SSTr2 > > U87-SSTr2 ≈ CHO-SSTr2 > > Jurkat-SSTr2 cells (Table 1). Repeated SSTr2 binding site assays and FACS analysis in hSSTr2/GFP-transduced CHO cells showed no significant change in the number of binding sites per cell or FACS profile over a 20-mo period.

In vitro uptake of 67Ga-DOTATOC by hSSTr2-transduced cells. (A) Accumulation of 67Ga-DOTATOC in WT and hSSTr-transduced cells after 2 h incubation at 37°C. natGa-DOTATOC or somatostatin-14 (1 μM) was used in blocking experiments. (B) Correlation of 67Ga-DOTATOC uptake vs. Bmax for different hSSTr2-transduced cell lines. NCA = no natGa-DOTATOC or somatostatin-14 added.
Kd and Bmax Values for natGa-DOTATOC Binding to hSSTr2 Expressed by Transduced Cells
67Ga-DOTATOC Uptake in hSSTr-Transduced and WT Cells
The uptake of 67Ga-DOTATOC in hSSTr2-transduced cells was significantly higher than that in corresponding WT (nontransduced) cells (Fig. 2A). The calculated transduced-to-WT ratios ranged from 8.1 ± 1.7 (Jurkat) to 108 ± 15 (C6). These results are consistent with the Bmax estimates (Table 1); a good correlation between the cell uptake values and Bmax estimates was obtained (Fig. 2B). Little or no 67Ga-DOTATOC was accumulated in CHO cells transduced with the hSSTr1, hSSTr3, or hSSTr4 genes (Fig. 2A). In addition, the uptake of 67Ga-DOTATOC in SSTr2-transduced cell lines was substantially blocked (>90%) with a 1,000-fold excess of gallium-DOTATOC or somatostatin-14 (Fig. 2A). These results demonstrated the high specificity of 67Ga-DOTATOC binding to the human SSTr2 receptor and the low natural expression of SSTr2 or SSTr5 in the WT cells.
68Ga-DOTATOC Biodistribution and Imaging
Small-animal PET images of 68Ga-DOTATOC uptake in hSSTr2-transduced and WT xenografts were obtained at 1 and 2 h after injection of the tracer (Fig. 3). The hSSTr2-transduced xenografts were clearly visualized and distinct from both adjacent background radioactivity and WT xenografts in the projection images (Figs. 3A and 3B). Coronal and sagittal slices distinctly show the hSSTr2 xenografts, kidneys, and bladder. GFP–fluorescence imaging also visualized the SSTr2-IRES-GFP–transduced xenografts but not WT tumors (Fig. 3B), consistent with the radiotracer-based in vivo imaging. As expected, the kidneys and bladder were not seen in the fluorescence images, whereas the small-animal PET images reflect the dominant renal clearance of 68Ga-DOTATOC from the body.

Small-animal PET and fluorescence imaging of hSSTr2-transduced xenografts. (A) Optical small-animal PET projection and transaxial images of mouse with 3 transduced xenografts at 2 h after injection of 68Ga-DOTATOC. (B) GFP–fluorescence, small-animal PET projection, and transaxial images of mouse with 2 WT and 2 transduced xenografts at 2 h after injection of 68Ga-DOTATOC. Radioactivity in kidneys and bladder is visualized in projection images. CT = C6-SSTr2; Cw = C6-WT; JT = Jurkat-SSTr2; UT = U87-SSTr2; Uw = U87-WT.
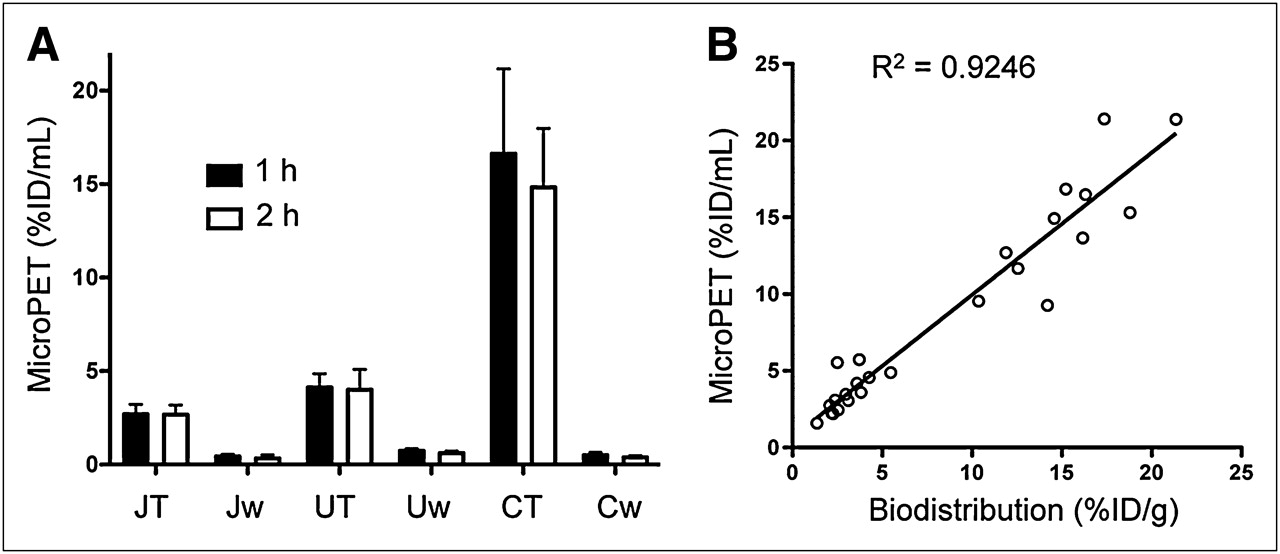
The 68Ga-DOTATOC organ uptake levels (%ID/mL) were obtained by region-of-interest analysis of the small-animal PET images and corrected for partial-volume effects. Xenograft uptake was similar at 1 and 2 h after injection, demonstrating rapid in vivo binding of 68Ga-DOTATOC to hSSTr2 receptors in these animal xenografts (Fig. 4A). The rank order of 68Ga-DOTATOC accumulation was C6-SSTr2 > U87-SSTr2 > Jurkat-SSTr2, which is consistent with the biodistribution results based on the ex vivo radioactivity measurements (Figs. 5A and 4B), Bmax values (Table 1), and in vitro uptake studies (Fig. 2A). The imaging values corrected for partial-volume effects correlated well with the tissue-sampling values (Fig. 4B). These results are consistent with SSTr2-specific small-animal PET with 68Ga-DOTATOC.
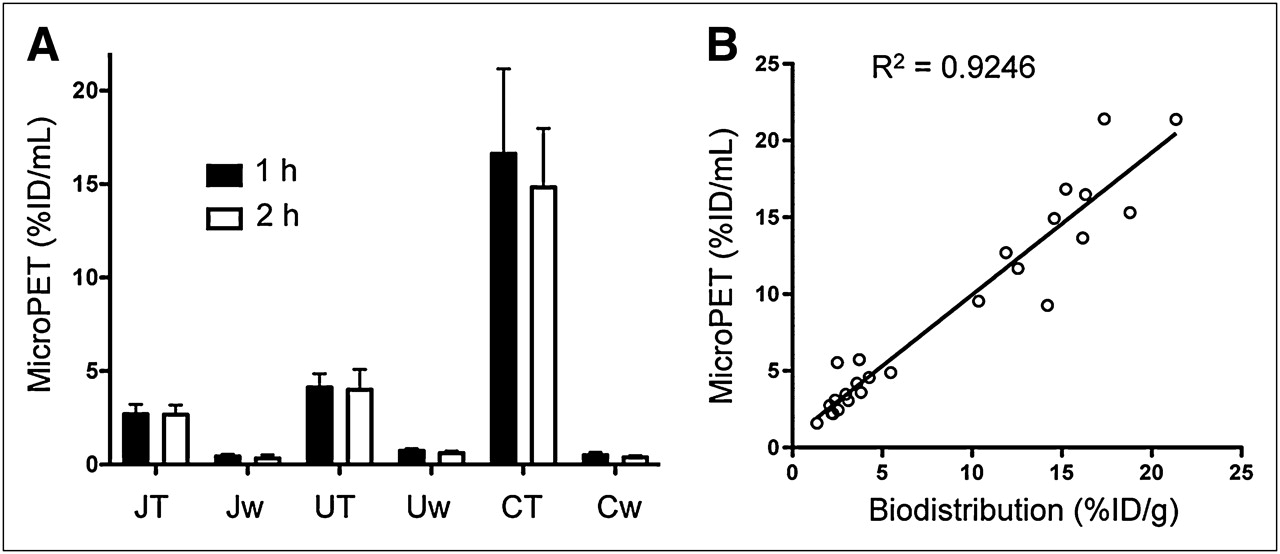
Small-animal PET tumor uptake of 68Ga-DOTATOC. (A) Uptake (%ID/mL) derived from small-animal PET at 1 and 2 h after injection of 68Ga-DOTATOC. (B) Correlation of uptake values derived from small-animal PET and biodistribution for 68Ga-DOTATOC. CT = C6-SSTr2; Cw = C6-WT; JT = Jurkat-SSTr2; Jw = Jurkat-WT; UT = U87-SSTr2; Uw = U87-WT.
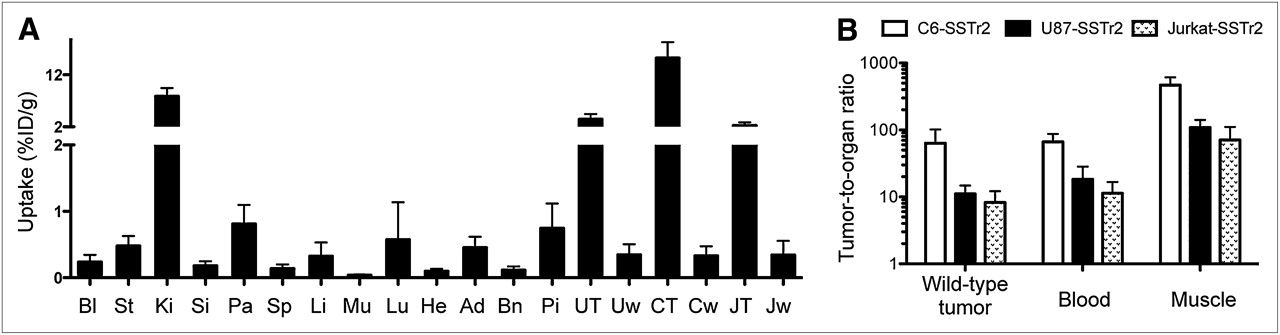
68Ga-DOTATOC biodistribution studies. (A) Biodistribution of 68Ga-DOTATOC (%ID/g ± SD) in hSSTr2-transduced tumor-bearing nude mice at 2 h after injection. (B) 68Ga-DOTATOC uptake ratios for hSSTr2-transduced xenograft and normal organs. Ad = adrenal; Bl = blood; Bn = bone; CT = C6-SSTr2; Cw = C6-WT; He = heart; JT = Jurkat-SSTr2; Jw = Jurkat-WT; Ki = kidney; Li = liver; Mu = muscle; Pa = pancreas; Pi = pituitary; Si = small intestines; Sp = spleen; St = stomach; UT = U87-SSTr2; Uw = U87-WT.
68Ga-DOTATOC levels were low in blood and other non–SSTr2-expressing organs, except the kidneys. Radioactivity in the muscle and blood was low at 0.03 ± 0.01 and 0.24 ± 0.08 %ID/g, respectively. Conversely, organs known to express moderate levels of SSTr2 showed higher uptake of radiotracer and included the pancreas (0.81 ± 0.22 %ID/g), adrenal gland (0.45 ± 0.14 %ID/g), and pituitary gland (0.74 ± 0.31 %ID/g). Except for the kidneys, the highest accumulation of radioactivity was measured in hSSTr2-transduced xenografts (Fig. 5A). These values ranged from 2.27 ± 0.44 %ID/g for Jurkat-SSTr2, to 3.46 ± 0.76 %ID/g for U87-SSTr2, to 15.2 ± 2.3 %ID/g for C6-SSTr2. In contrast, low uptake was measured in all corresponding WT xenografts (Fig. 5A). The uptake ratio between hSSTr2-transduced and nontransduced xenografts was 64 ± 31 for C6, 11.1 ± 2.9 for U87, and 8.3 ± 3.2 for Jurkat; comparisons to blood and muscle are also shown (Fig. 5B). These results were also consistent with the ex vivo radioautography (Supplemental Fig. 3).
There was no significant difference between the growth rates of the C6-WT or C6-SSTr2 xenografts at 9–15 d after implantation of tumor cells (Table 2). However, there was a highly significant growth delay for Jurkat-SSTR2 cells, compared with Jurkat-WT cells (P = 0.029), with the mean tumor weight being around 25-fold lower. A similar but less dramatic effect was seen for the U87-SSTr2 xenografts (3-fold lower at 8 d, P = 0.022). Despite this, there was no correlation between 68Ga-DOTATOC uptake and xenograft size or age.
Effect of SSTr2 Expression on Xenograft Growth in Athymic Mice
Linear regression analysis of SSTr2 tumor uptake (%ID/g) versus the expression level of hSSTr2 (Bmax) is shown in Figure 6. These results are consistent with specific, receptor-mediated uptake of 68Ga-DOTATOC and indicate that the expression level of hSSTr2 determines the magnitude of in vivo uptake by the transduced xenografts.
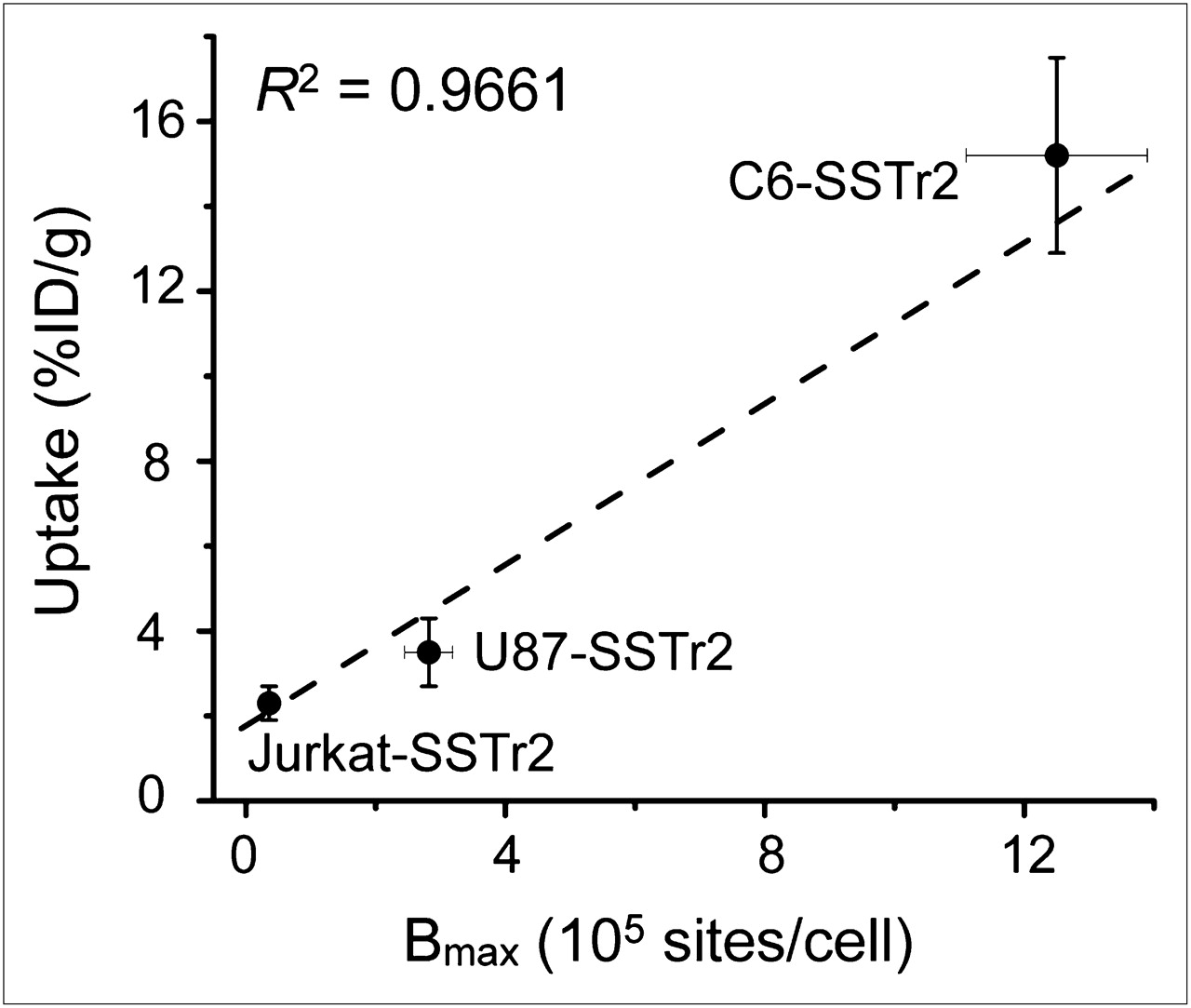
Correlation of in vivo tumor uptake of 68Ga-DOTATOC (%ID/g) and in vitro hSSTr2 expression Bmax.
DISCUSSION
The hSSTr2 gene–68GaDOTATOC reporter system has distinct imaging advantages over other hSSTr2–radioligand systems, other human reporter gene systems, and the HSV1-tk and HSV1-tksr reporter systems (13). Compared with 3 other categories of reporter genes (fluorescent proteins, intracellular enzymes, and cell membrane transporters), hSSTr2 belongs to the category of cell membrane receptors. hSSTr2 is a transmembrane protein, and its binding domain is located on the external plasma membrane (13). The hSSTr2 gene encodes a transmembrane protein that does not require radioligand permeation of the cell membrane to bind to the receptor. We showed that this process was rapid; the half-lives for binding and release from the receptor was on the order of several minutes. Interestingly, internalization of the DOTATOC ligand may be cell type–specific, because kinetic differences were observed between C6-, U87-, and CHO-transduced cells. The intracellular domain is associated with G-protein–linked signaling. Whenever an agonistic ligand binds to SSTr2, rapid internalization of the receptor–ligand complex will occur. The SSTr2 receptor will be reexpressed on the cell membrane and be ready for binding with new ligand within 24 h (21).
The hSSTr2 reporter gene can be compared with other human reporter genes, including the mitochondrial thymidine kinase 2 (22), sodium iodide symporter (23), and norepinephrine transporter (24) genes. A comparison of the hSSTr2–radioligand systems published in the literature is provided in Table 3, and a summary of other published reporter systems studied in our laboratory is provided in Supplemental Table 1. The comparisons in Table 3 and Supplemental Table 1 demonstrate the potential advantages of the hSSTr2 gene–68Ga-DOTATOC reporter system. A significant time dependence for optimal transduced–to–nontransduced tumor uptake ratios has been demonstrated for other reporter–radioligand systems, and optimal images are obtained at late imaging times (22,24). However, radiolabeled somatostatin analogs (small peptide) generally display rapid accumulation and can be excreted rapidly as well—especially true for 68Ga-DOTATOC. Rapid uptake in the reporter-transduced xenografts and rapid excretion of nonbound ligand were observed. As a result, 68Ga-DOTATOC showed high transduced–to–nontransduced tumor uptake ratios at early imaging times (1 h after injection), and image quality at 2 h after injection did not improve significantly, reflecting rapid binding and high affinity in the setting of rapid renal clearance. 68Ga-DOTATOC uptake in reporter-transduced xenografts was directly proportional to hSSTr2 expression levels in the xenografts, indicating that early PET with 68Ga-DOTATOC could serve as a basis for the noninvasive quantification of hSSTr2 gene expression. Thus, the 68Ga-DOTATOC–hSSTr2 system has potential for being an ideal human reporter gene system for translation to clinical studies.
Comparison of Radioligands for Imaging Human SSTr Reporter Gene
Although the hSSTr2 gene was originally described as a promising reporter gene in 1999 (14) and again in 2005 (17) (Table 2), image quality remained suboptimal using 111In-DTPA-octreotide, 99mTc-P829, 188Re-P829, 99mTc-P2045, or 94mTc-demotate-1 radioligands (Table 3). This has now changed significantly with the availability of 68Ga-DOTATOC. DOTATOC is a peptide–chelate conjugate originally designed for radiotherapy of SSTr2-expressing tumors (18). It has been classified as a receptor agonist and has undergone extensive clinical testing in both Europe (3) and the United States (25) and is being developed as a clinical imaging and targeted radiotherapeutic agent. Gallium-DOTATOC has been reported as an SSTr2- or SSTr5-selective ligand; it displaces the binding of the panspecific ligand 125I-Tyr11-SST14 to SSTr1–SSTr5 with an inhibitory concentration of 50% (IC50) of >1,000 for SSTr1, 2.5 for SSTr2, 613 for SSTr3, >1,000 for SSTr4, and 73 nM for SSTr5 (26). Although direct Kd values have not been determined for these analogs, our calculated binding affinity of 5–7 nM for gallium-DOTATOC is consistent with the 2.5 nM IC50 value for 125I-Tyr11-SST14 binding. Other somatostatin analogs such as vapreotide and lanreotide are SSTr2-, SSTr3-, and SSTr5-specific, with low nanomolar binding affinities to these receptor subtypes (27). The FDA-approved P829 ligand has a similar SSTr2-, SSTr3-, and SSTr5-binding profile, with IC50 values of 2.5, 1.5, and 2.0, respectively (28). On the basis of SSTr binding affinities, gallium-DOTATOC is the more optimal agent for imaging SSTr2 in vivo. Consistent with these findings, 68Ga-DOTATOC has been used successful in imaging neuroendocrine tumors in patients.
Apart from being SSTr2-specific, 68Ga-DOTATOC can be rapidly prepared with the generator-produced PET radionuclide 68Ga (half-life, 68.3 min; maximum energy of the positron electron, 1.89 MeV, 89%). This isotope has a high abundance of β+ particles, which can be quantified by clinical PET cameras. The short half-life is ideally suited to the rapid blood clearance of the ligand and reduces patient radiation dose. Because 68Ga is a generator-produced radionuclide, it can be produced and used at nuclear medicine facilities with no onsite cyclotron, using a simple kit preparation procedure.
Several issues not addressed in this study relate to the potential biologic effects of transducing a cell membrane signaling receptor on the transduced cells. In vitro, activation of SSTr2 on tumor cells results in cell growth arrest, whereas activation of SSTr5 results in cell proliferation. Moreover, selective SSTr2 and selective SSTr5 agonists counteract and antagonize each other (29,30). For carcinoid tumors, this counteraction seems to explain why SSTr2 agonists can reduce symptomatic effects but only some tumors show growth responses. Moreover, for somatotropinomas, Taboada et al. (31) have shown that patients who have high SSTr2–SSTr5 messenger RNA tumor levels respond better to octreotide therapy than those with low ratios. In vivo animal studies have also shown that SSTr2-transfected xenografts are more sensitive to somatostatin treatment (32,33), and the SSTr2-transfected tumors grow more slowly than WT tumors. For example, Vernejoul et al. observed that endogenous somatostatin was able to suppress the growth after in vivo SSTr2 gene transfer to orthotopically implanted pancreatic tumors (32).
We observed variable effects of SSTr2 transduction on xenograft growth rates. C6-SSTr2 xenografts, which had high SSTr2 expression in vitro and high 68Ga-DOTATOC uptake in vivo, did not significantly differ in the tumor size from C6-WT xenografts. Compared with U87-WT xenografts, U87-SSTr2 xenografts, which had moderate SSTr2 expression in vitro and moderate 68Ga-DOTATOC uptake in vivo, showed a significant delay in tumor growth at 8 d after implantation. Finally, compared with nontransduced, WT Jurkat xenografts, Jurket-SSTr2 xenografts, which had the lowest SSTr2 expression in vitro and moderate 68Ga-DOTATOC uptake in vivo, demonstrated a large and significant growth delay (tumor size) at 35 d after implantation. These results suggest considerable intertumor variability with respect to the ability of endogenous somatostatin to suppress tumor growth.
Because SSTr2 gene therapy can dramatically affect tumor growth rate, a constitutively expressed SSTr2 reporter gene may not be widely applicable for monitoring tumor trafficking, growth, and metastases. For similar reasons, the use of an inducible SSTr2 reporter gene to monitor cell signaling and function is likely to be compromised by endogenous somatostatin ligands. For example, the SSTr2 reporter gene is unlikely to be useful for monitoring T cells in vivo, because T cells are known to express SSTr2 and activation of this receptor leads to the suppression of γ-interferon (34) secretion. In activated T cells, there is an upregulation of SSTr5 (35), which when activated in tumor cells abrogates the effects of SSTr2 activation (30). Because we and others have shown that SSTr2 transfection of tumor cells can slow their growth in vivo, it is highly plausible that SSTr2-transfected T cells will demonstrate phenotypic alterations associated with activation of hSSTr2-mediated signaling by endogenous somatostatin.
To avoid endogenous SSTr2 signaling in SSTr2 reporter–transduced cells, modifications of the hSSTr2 gene are necessary to abrogate its signal transduction function without compromising its ligand-binding receptor function. Such a strategy has been shown to be successful for the dopamine 2 receptor (36). Recently, Han et al. showed that hSSTr2-activated signaling can be eliminated by selective truncation of the intracellular domain of the receptor (37). They developed a mutated hSSTr2 gene, HA-SSTr2Δ314, and showed that this truncation of the intracellular domain eliminated SSTr2-mediated intracellular signaling, as measured by the absence of effects on cAMP, cGMP, and proliferation, but does not alter 111In-DTPA-octreotide binding to the receptor. Their in vivo SPECT study of xenografts transduced with the truncated receptor (HA-SSTr2Δ314) yielded results similar to those of xenografts transduced with the WT receptor (37).
CONCLUSION
Our studies show that high specificity and sensitivity for imaging hSSTr2 reporter gene expression is obtained with 68Ga-DOTATOC and that, compared with other SSTr2-binding radioligands, it provides high transduced–to–nontransduced (target-to-background) radioactivity ratios at early imaging times. The hSSTr2–68Ga-DOTATOC reporter system is a promising combination for translation to clinical studies, providing that endogenous SSTr2 signaling does not significantly affect the biology of the transduced cells. Alternatively, the application of a truncated hSSTr2 gene, such as HA-SSTr2Δ314, may be more desirable.
Acknowledgments
We thank Dr. Pat Zanzonico and Valerie Longo for imaging support, Dr. Ekaterina Moroz for technical support, and Dr. Friedrich Raulf of Novartis (Basle, Switzerland) for kindly supplying SSTr-transfected CHO cells. We also appreciate Dr. Graeme Bell (University of Chicago, Chicago, IL) for kindly supplying the hSSTr2 gene. This work was supported by NIH grant P50-CA84638. The Memorial Sloan-Kettering Cancer Center Small Animal Imaging Core Facility was supported by NIH Small-Animal Imaging Research Program grant R24 CA83084 and NIH Center grant P30 CA08748.
- © 2011 by Society of Nuclear Medicine
REFERENCES
- Received for publication May 12, 2010.
- Accepted for publication September 23, 2010.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Imaging of T-cell Responses in the Context of Cancer Immunotherapy
- Molecular Imaging with Reporter Genes: Has Its Promise Been Delivered?
- Specific Targeting of Somatostatin Receptor Subtype-2 for Fluorescence-Guided Surgery
- Bicyclic Peptides as a New Modality for Imaging and Targeting of Proteins Overexpressed by Tumors
- Antibody with Infinite Affinity for In Vivo Tracking of Genetically Engineered Lymphocytes
- 68Ga-DOTATOC and FDG PET Imaging of Preclinical Neuroblastoma Models
- Dynamic imaging for CAR-T-cell therapy
- Comparative Analysis of T Cell Imaging with Human Nuclear Reporter Genes
- Imaging Mouse Cancer Models In Vivo Using Reporter Transgenes
- Noninvasive Molecular Imaging Using Reporter Genes
- Radiopeptide Imaging and Therapy in the United States