


Abstract
Monitoring genetically altered T cells is an important component of adoptive T cell therapy in patients, and the ability to visualize their trafficking/targeting, proliferation/expansion, and retention/death using highly sensitive reporter systems that do not induce an immunologic response would provide useful information. Therefore, we focused on human reporter gene systems that have the potential for translation to clinical studies. The objective of the in vivo imaging studies was to determine the minimum number of T cells that could be visualized with the different nuclear reporter systems. We determined the imaging sensitivity (lower limit of T cell detection) of each reporter using appropriate radiolabeled probes for PET or SPECT imaging. Methods: Human T cells were transduced with retroviral vectors encoding for the human norepinephrine transporter (hNET), human sodium-iodide symporter (hNIS), a human deoxycytidine kinase double mutant (hdCKDM), and herpes simplex virus type 1 thymidine kinase (hsvTK) reporter genes. After viability and growth were assessed, 105 to 3 × 106 reporter T cells were injected subcutaneously on the shoulder area. The corresponding radiolabeled probe was injected intravenously 30 min later, followed by sequential PET or SPECT imaging. Radioactivity at the T cell injection sites and in the thigh (background) was measured. Results: The viability and growth of experimental cells were unaffected by transduction. The hNET/meta-18F-fluorobenzylguanidine (18F-MFBG) reporter system could detect less than 1 × 105 T cells because of its high uptake in the transduced T cells and low background activity. The hNIS/124I-iodide reporter system could detect approximately 1 × 106 T cells; 124I-iodide uptake at the T cell injection site was time-dependent and associated with high background. The hdCKDM/2′-18F-fluoro-5-ethyl-1-β-d-arabinofuranosyluracil (18F-FEAU) and hsvTK/18F-FEAU reporter systems detected approximately 3 × 105 T cells, respectively. 18F-FEAU was a more efficient probe (higher uptake, lower background) than 124I-1-(2-deoxy-2-fluoro-1-d-arabinofuranosyl)-5-iodouracil for both hdCKDM and hsvTK. Conclusion: A comparison of different reporter gene–reporter probe systems for imaging of T cell number was performed, and the hNET/18F-MFBG PET reporter system was found to be the most sensitive and capable of detecting approximately 35–40 × 103 T cells at the site of T cell injection in the animal model.
Adoptive immunotherapy using genetically engineered T cells is a promising and rapidly evolving field. Further advances in the field of adoptive therapy will require the ability to monitor the trafficking, targeting, and activation/proliferation of the administered cells. The application of novel reporter systems (gene–probe combinations) together with noninvasive imaging modalities, such as PET, has shown the potential for monitoring T cells in vivo (1–4). Herpes simplex virus type 1 thymidine kinase (hsvTK) and mutant hsv1sr39TK reporter genes have been used in most of these seminal studies. Radiolabeled reporter probes, including 1-(2-deoxy-2-fluoro-1-d-arabinofuranosyl)-5-iodouracil (FIAU) (labeled with 131I (5), 123I (5), 124I (6), and 125I (7)), 2′-18F-fluoro-5-ethyl-1-β-d-arabinofuranosyluracil (18F-FEAU) (8), and 9-(4-18F-fluoro-3-[hydroxymethyl]butyl)guanine (18F-FHBG) (9) have been used for imaging hsvTK in these studies, and comparisons of their efficacy have previously been performed (10–12). However, there has been a recent focus on human reporter systems, to avoid the potential risk of generating an immunologic reaction to viral (nonhuman) reporter systems, including hsvTK and hsvsr39TK reporter–transduced T cells.
We and others have tested different human-derived reporter systems for potential cell tracking in patients (12–21). These intrinsically nonimmunogenic reporter systems include the human sodium iodide symporter (hNIS)/124I-iodide (13), the human norepinephrine transporter (hNET)/123I/124I-metaiodobenzylguanidine (MIBG) (14) and 18F-meta-18F-fluorobenzylguanidine (18F-MFBG) (15), the human somatostatin receptor 2 (SSTR2/68Ga-DOTATOC) (16,17), the human ferritin/2+Fe reporter (18), and the human transferrin receptor (19). More recently described reporter genes include the truncated and mutated human mitochondrial thymidine kinase type 2 (h∆TK2, h∆TK2DM) genes (20,21) and the mutated human deoxycytidine kinase (hdCKDM) gene (22–24). Both human kinases were shown to exclusively phosphorylate pyrimidine-based PET probes, including 124I-FIAU, 18F-FEAU, and 2′-deoxy-2′-18F-fluoro-5-methyl-1-β-l-arabinofuranosyluracil.
The objective of this report was to compare the T cell imaging efficacy of 1 viral and 3 human reporter genes with different imaging probes (different reporter systems). We compared the hNET/18F-MFBG and 123I/124I-MIBG reporter systems with the previously described hNIS, hdCKDM, and hsvTK reporter systems using radiolabeled 124I-iodide, 18F-FEAU, and 124I-FIAU, respectively. We included 18F-MFBG as an alternative to 123I/124I-MIBG, for imaging hNET reporter–transduced T cells, because of the superior tumor-to-background images that can be obtained at earlier times after administration of MFBG compared with MIBG (15,25). Using a murine preclinical model, we determined the minimum number of reporter-transduced T cells that could be imaged with the different reporter systems. Our results demonstrate that the hNET/18F-MFBG reporter system is the most sensitive and allows for repetitive PET imaging of hNET-expressing T cells.
MATERIALS AND METHODS
Development of New Retroviral Vectors
Two new retroviral vectors, which carry the constitutively expressed hNET or hNIS separated by IRES (internal ribosomal entry site) with green fluorescent protein (GFP), were developed by recloning of the hNET (14) and hNIS (14) genes into the retroviral SFG backbone. The hsvTK/GFP and hdCKDM/GFP vectors were developed previously (Supplemental Fig. 1; supplemental materials are available at http://jnm.snmjournals.org) (13,26). The hsvTK-IRES-GFP and hdCKDM-IRES-GFP dual reporters were subsequently developed to compare the transduced-to-nontransduced T cell uptake ratio of the fusion and IRES-linked reporter systems for both hsvTK and hdCKDM (Supplemental Fig. 2).
T Cell Isolation, Transduction, Assessment, and Expansion
Buffy coat–containing T cells (60 mL) were obtained from the New York Blood Bank from healthy donors, who gave informed consent. T cells were isolated using Ficoll isolation medium as previously described (27). At 48 h after stimulation with phytohemagglutinin at a concentration of 2 μg/mL, T cells were transduced with a reporter gene–bearing retrovirus in the presence of interleukin-2 (20 U/mL), as described previously (4). During T cell expansion, the medium was supplemented with IL-15 at a concentration of 10 ng/mL every 3 d. T cell viability was measured using trypan blue exclusion. Analysis of GFP expression in all experimental cell populations was performed using a FACSCalibur analyzer (BD Biosciences), and the purity and fluorescence of cells were assessed using Cell Quest Software (Becton Dickinson). All assays were repeated at least 3 times.
Reporter T Cell Studies: In Vitro Uptake of Radiolabeled Probes
The radiolabeled imaging probes were prepared as previously described. 18F-MFBG (∼19 GBq/μmol) (15), 18F-FEAU (∼22 GBq/μmol) (28), carrier-free 124I-MIBG (>12 GBq/μmol), carrier-free 124I-iodide, and carrier-free 124I-FIAU were synthesized at Memorial Sloan Kettering Cancer Center (28). 123I-MIBG (clinical formulation) was obtained from Nuclear Diagnostics Products.
Triplicate samples containing 1.0 × 106 reporter-transduced or wild-type T cells in a total volume of 1.0 mL were incubated in RMPI medium containing fetal calf serum (10%) with 18.5 kBq/mL (0.5 μCi/mL) of each specific probe. The samples were gently shaken at 37°C for 2 h. After incubation, the cells were isolated by rapid filtration through glass microfiber filters and washed with 3 × 2 mL of ice-cold Tris-buffered saline (pH 7.4). The radioactivity in cells was measured with a γ-Packard γ counter (United Technologies).
Xenotransplantation of T Cells
All animal studies were performed under a Memorial Sloan Kettering Cancer Center protocol approved by the Institutional Animal Care and Use Committee. Athymic rnu/rnu male mice were obtained from the National Cancer Institute; the mice were 3 mo old at the time of the imaging. The mice were anesthetized by inhalation of isoflurane/oxygen and maintained on a heating pad. After completion of the imaging studies, all animals were sacrificed by CO2 inhalation.
T cells were prepared in fresh batches for each cohort of animals and assessed for viability before every set of injections. Every batch of T cells was normalized for transduction efficacy and percentage live cells and diluted in 50 μL of T cell medium. Subcutaneous T cell injections in both shoulders was performed over a 30-s period using a 28.5-gauge needle/syringe (2–3 mm lateral from the scapula).
Eight groups of mice were studied, 1 for each reporter system (8 reporter gene–radiolabeled probe combinations): hNET/18F-MFBG, hdCKDM/18F-FEAU, hsvTK/18F-FEAU (groups 1–3); hNET/124I-MIBG; hNIS/124I-iodide, hdCKDM/124I-FIAU, and hsvTK/124I-FIAU (groups 4–7); and hNET/123I-MIBG (group 8). Four cohorts of animals, injected with different numbers of T cells, were studied for each of the reporter gene–radiolabeled probe combinations, except for hNET/123I-MIBG (group 8) for which 4 cohorts were analyzed. Animals in cohort A of groups 1–7 (n = 8 animals/reporter system) received a subcutaneous injection of reporter-transduced T cells (105 and 106) in opposite shoulders. Animals in cohort B of groups 1–7 (n = 8 animals/reporter system) received a subcutaneous injection of reporter-transduced T cells (3 × 105 and 3 × 106) in opposite shoulders. Mice in group 8 (123I-MIBG/hNET; n = 17) were divided into 3 cohorts: cohort A (105 and 106 T cells), cohort B (3 × 105 and 3 × 106 T cells), and cohort C (107 and 3 × 107 T cells). Thirty minutes after T cell injection, animals received an intravenous injection of the appropriate/corresponding radiolabeled probe.
Nuclear Imaging of Primary T Cells
Animals from the hNET/18F-MFBG, hsvTK/18F-FEAU, and hdCKDM/18F-FEAU groups (1–3) were injected with 7.4 MBq (200 μCi) of radiolabeled probe, respectively, and imaged 2 and 4 h after tracer administration. Animals from the hNET/124I-MIBG, hNIS/124I-iodide, hdCKDM/124I-FIAU, and hsvTK/124I-FIAU groups (4–7) were injected with 14.8 MBq (400 μCi) of radiolabeled probe, respectively, and imaged 4 and 24 h after administration. Animals from the hNET/123I-MIBG group (8) were injected with 29.6 MBq (800 μCi) of 123I-MIBG and imaged 4 and 24 h after administration. PET imaging was performed using a Focus 120 microPET scanner (Siemens Medical Solutions USA, Inc.). A narrow energy window of 420–580 keV and a coincidence timing window of 6 ns were used for 124I-labeled probes, and a wide 350- to 750-keV window was used for 18F-labeled probe. The image data were corrected for nonuniformity of scanner response, dead time count losses, and physical decay to the time of injection. No correction was applied for attenuation, scatter, or partial-volume averaging.
Region-of-interest analysis of the acquired images was performed using ASIPro software (Siemens Medical Solutions USA, Inc.); total radioactivity and the maximum pixel values were recorded for each T cell injection site. Iodide uptake in the thyroid and stomach was not blocked, and radioactivity in these organs reflects radioactive iodide release after the slow catabolism of 124I-MIBG. Because of the rapid uptake and washout/clearance of free 124I-iodide, multiple imaging time points (0.5, 1, 2, 4, and 24 h) were obtained for the hNIS/124I-iodide reporter T cell–bearing animals.
SPECT imaging was performed on each animal receiving 123I-MIBG using a nano-SPECT/CT dedicated small-animal scanner (Bioscan Inc. USA). A photo peak energy window of 143–175 keV and a low-energy, high-resolution collimator were used to acquire the 123I images. Transverse images were reconstructed by filtered backprojection. Quantification was performed using InVivoScope Integrated proprietary Bioscan Analysis Software.
Statistics
The statistical significance of differences between mean values was estimated with Excel (Microsoft) using the independent t test for unequal variances. P values of less than 0.05 were considered to be statistically significant.
RESULTS
Characterization of Reporter Gene–Transduced Primary Human T Cells
After transduction, reporter-bearing primary human T cells were characterized for viability and reporter expression. Fluorescence-activated cell sorting profiles demonstrated a high fraction of viable and GFP-positive reporter cells. Each transduction yielded a high percentage of GFP-positive cells: 77.8% for hNET/GFP, 72.4% for hNIS/GFP, 83.4% for human hdCKDM/GFP, and 77.6% for hsvTK/GFP-transduced T cells, respectively, and high mean fluorescence levels corresponding to the respective vector design. All primary T cell groups demonstrated the same rate of proliferation as wild-type cells and high viability (>85%) (Supplemental Fig. 3).
In Vitro Reporter–Transduced Human T Cell Uptake Studies
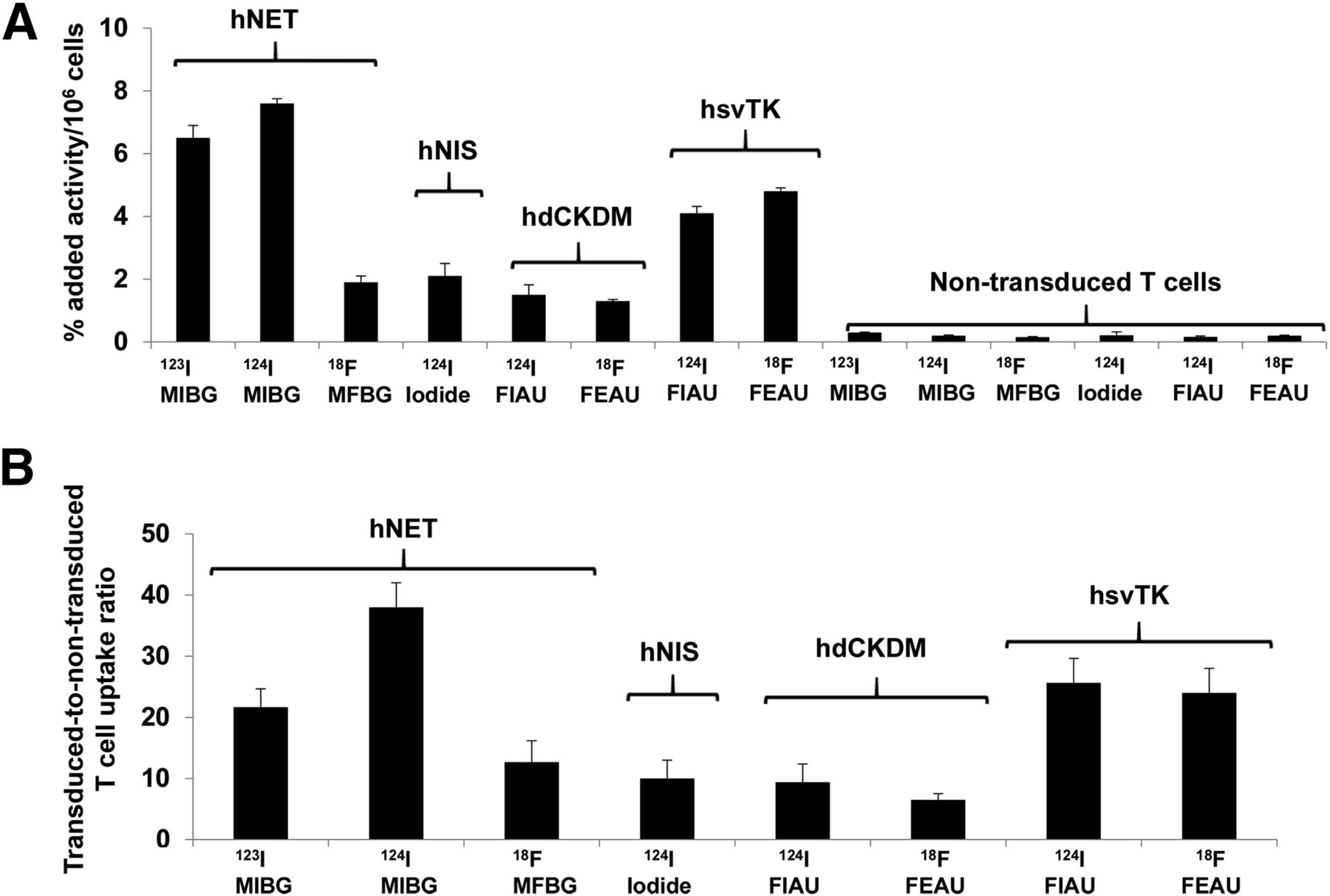
The initial assessment and comparison of the 4 reporter systems in human T cells was performed in vitro using a radiolabeled probe uptake assay (Fig. 1). The highest uptake levels were obtained with 123I-MIBG and 124I-MIBG in hNET reporter–bearing T cells after 2 h of incubation (6.5 ± 0.4 and 7.6% ± 0.1% of added radioactivity per 106 cells, respectively). Similarly, the hNET-transduced–to–nontransduced T cell ratios were also high. These values were significantly higher than those obtained with 18F-MFBG (1.9% ± 0.2% per 106 cells), which is consistent with prior in vitro uptake studies comparing MIBG and MFBG uptake in hNET-expressing tumor cells (15,25). Comparable values were measured in hNIS (2.2% ± 0.9% per 106 cells), hsvTK (4.1% ± 0.41% per 106 cells), and hdCKDM (1.5% ± 0.25% per 106 cells) reporter–bearing T cells with 124I-iodide, 18F-FEAU, and 124I-FIAU, respectively.
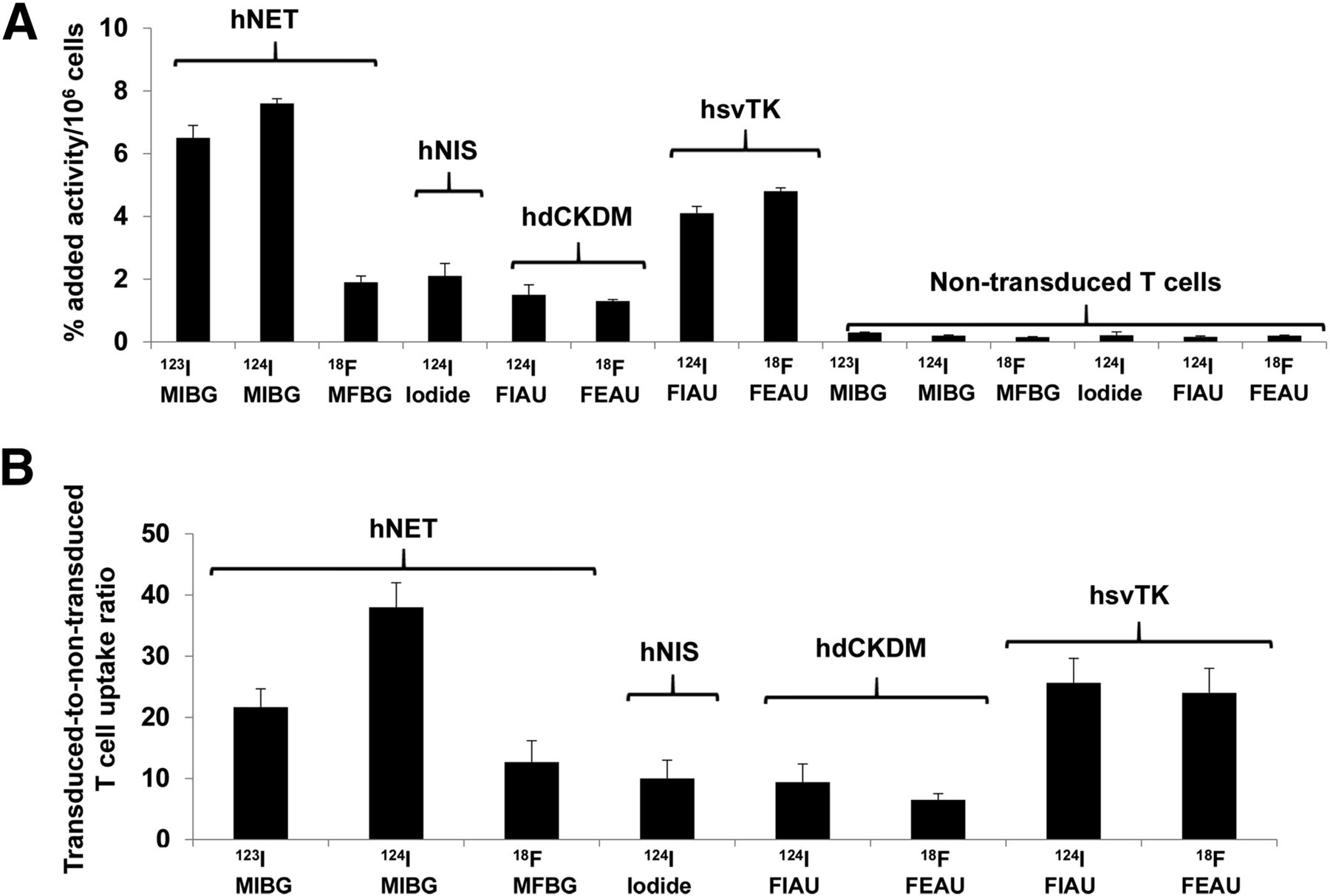
In vitro uptake of radiolabeled probes in human T cells transduced with different reporters. Data are presented as percentage added radioactivity/106 reporter-transduced T cells (A) and as transduced-to-nontransduced T cell uptake ratios (B). Data are presented from 3 independent experiments.
In Vivo Imaging of Reporter-Transduced Human T Cells in Athymic Mice
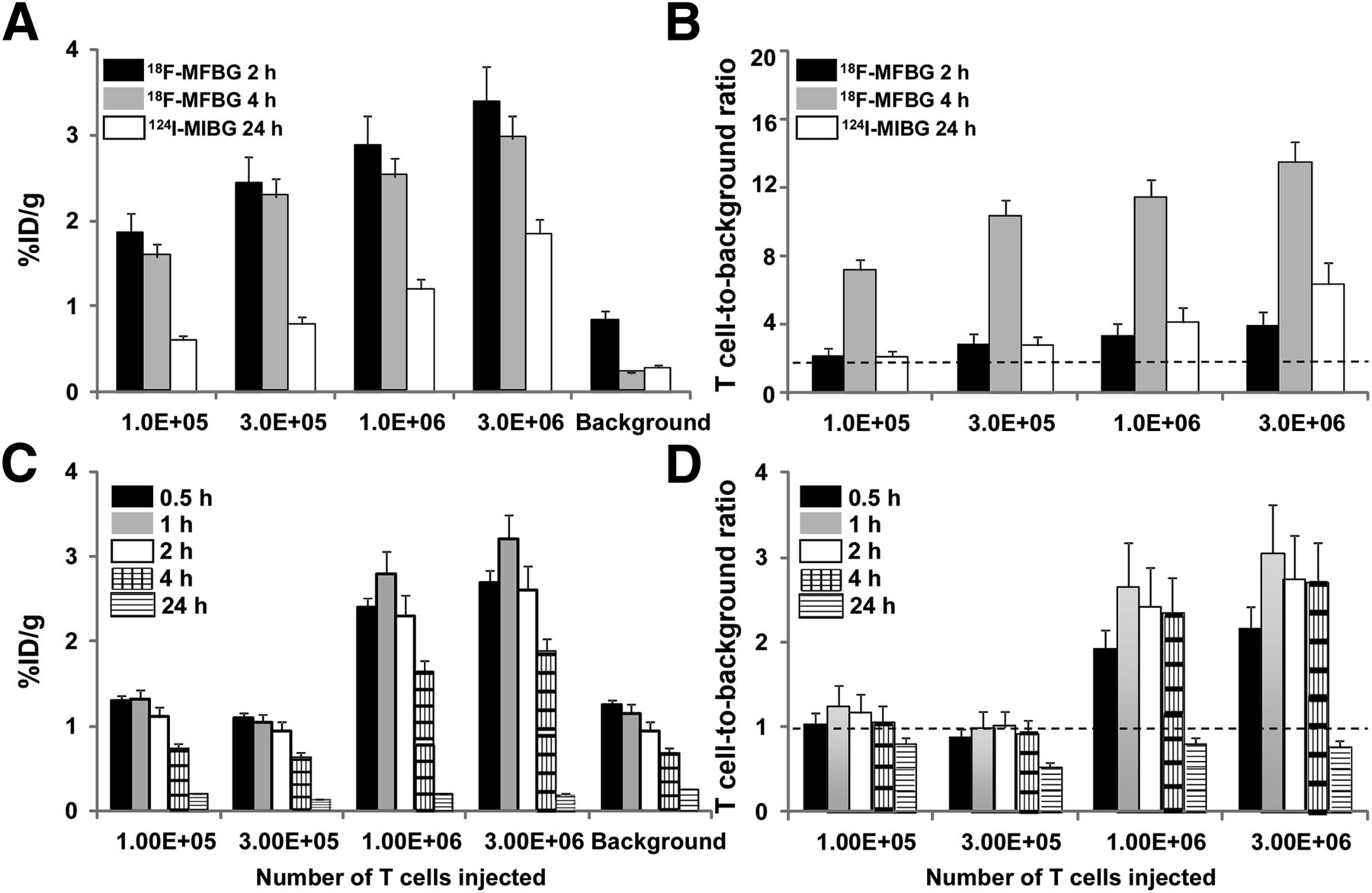
A comparison of the hNET and hNIS reporter systems is shown in Figures 2 and 3. Prior studies have shown that 18F-MFBG and 124I-MIBG accumulation in hNET-expressing tumor cells is stable between 2 and 4 h after injection (15,25), whereas 124I-iodide accumulation peaks at approximately 1 h and then washes out of hNIS-expressing cells and tumors (13,29,30). Thus, imaging was performed more frequently in the hNIS/124I-iodide experiments. Although the high background of 124I-MIBG at early (4 h) time points prevented the detection of individual signals from T cell injection sites, the long (∼4 d) half-life of 124I-MIBG allowed for late (24 h) imaging. Although the maximum accumulation of 18F-MFBG and 124I-iodide radioactivity by the subcutaneously injected reporter-bearing T cells was similar, the contrast (T cell–to–background radioactivity ratio) was significantly higher for the hNET/18F-MFBG reporter system (∼14/1 at 4 h) than for hNET/124I-MIBG reporter (∼7/1 at 24 h) and hNIS/124I-iodide reporter (∼3/1 at 1–4 h) for the highest T cell–injected number (3 × 106). Because of the lower background, the hNET/18F-MFBG reporter was able to detect less than 105 T cells, compared with approximately 106 T cells for the hNIS/124I-iodide and hNET/124I-MIBG reporter (Table 1). The SPECT studies of hNET/123I-MIBG T cell imaging failed to visualize 3 × 106 cells. Therefore, ultrahigh numbers (107 and 3 × 107) of hNET reporter T cells were injected, followed by 29.6 MBq (800 μCi) of 123I-MIBG and SPECT imaging at 4 and 24 h. The results of this additional study demonstrated a clear signal at the injection site of 3 × 107 hNET reporter T cells but not at the 107 T cell injection site (Supplemental Fig. 4).

PET imaging of human primary T cells transduced with hNET (A) or hNIS (B) reporters. Different numbers of T cells were injected subcutaneously, followed by systemic administration of corresponding radiopharmaceuticals and PET imaging at respective time points. Number of T cells injected is shown in right upper and lower panels. %ID/g = percentage injected dose per gram.
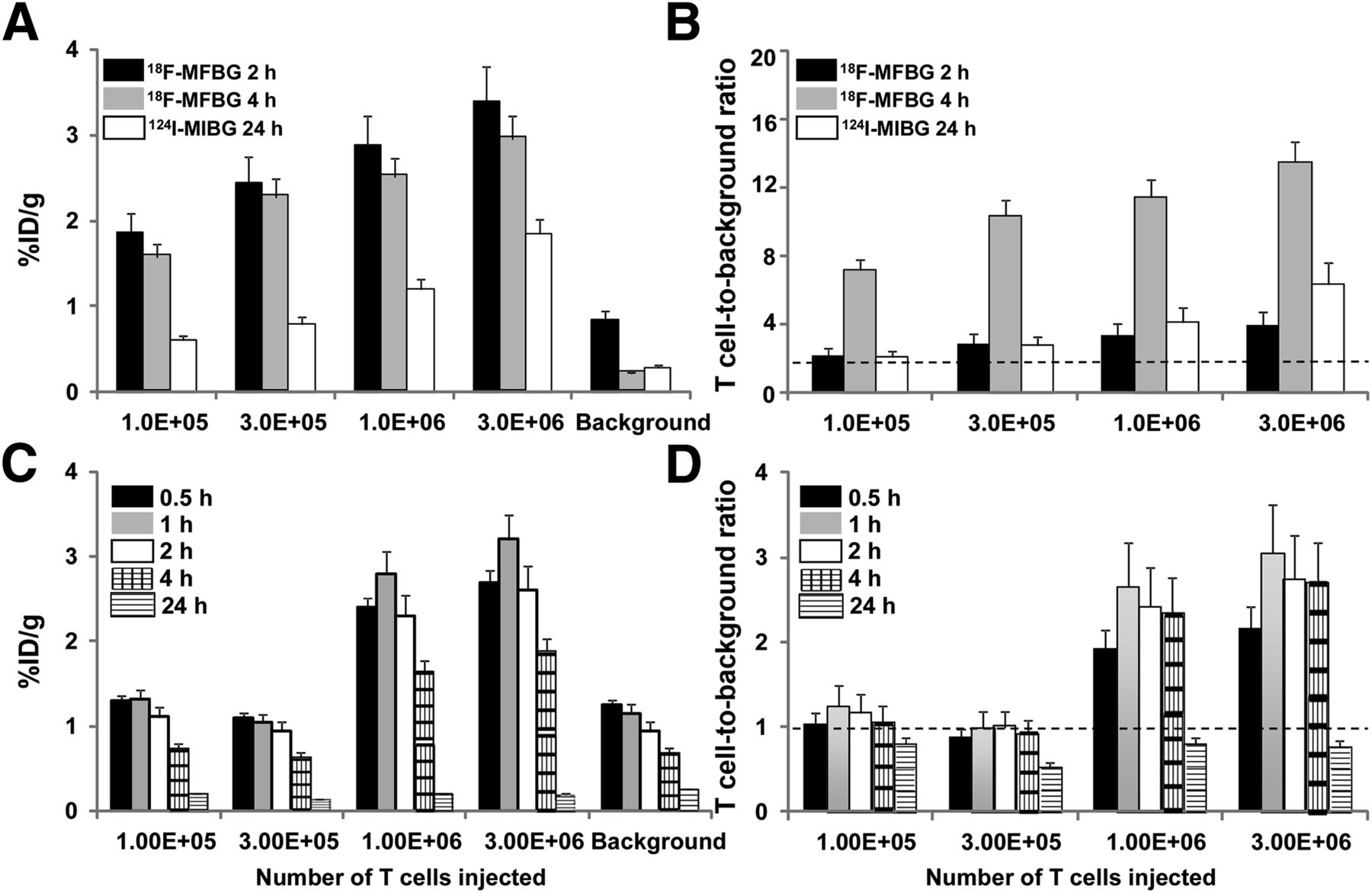
Analysis of PET images of human T cells transduced with hNET (A and B) and hNIS (C and D) reporters, as shown in Figure 2. Data are %ID/g ± SD (A and C) and T cell–to–background ratios ± SD (B and D). Data are from 2 independent experiments. %ID/g = percentage injected dose per gram.
Sensitivity of T Cell Number–Dependent Reporter Imaging Using PET
The T cell imaging patterns observed with the hdCKDM/GFP and herpes simplex virus hsvTK/GFP fusion reporters with either 18F-FEAU or 124I-FIAU were similar and between those observed with the hNET/18F-MFBG and hNIS/124I-iodide reporter systems (Figs. 4 and 5). Because both reporter systems have been shown previously to result in fairly stable radioactivity accumulation in tumor cells between 1 and 4 h after injection of tracer (11,12,24), we studied only the 4-h time point to allow for optimal clearance of 18F-FEAU and 124I-FIAU background radioactivity. 18F-FEAU yielded significantly higher T cell accumulation values than 124I-FIAU at 4 h for both reporters, and the T cell–to–background ratios with 18F-FEAU were higher than with 124I-FIAU, partly due to the lower background values observed with 18F-FEAU. Both hdCKDM/18F-FEAU and the hsvTK/18F-FEAU reporter systems were able to detect approximately 3 × 105 T cells at 4 h (Fig. 5; Table 1).
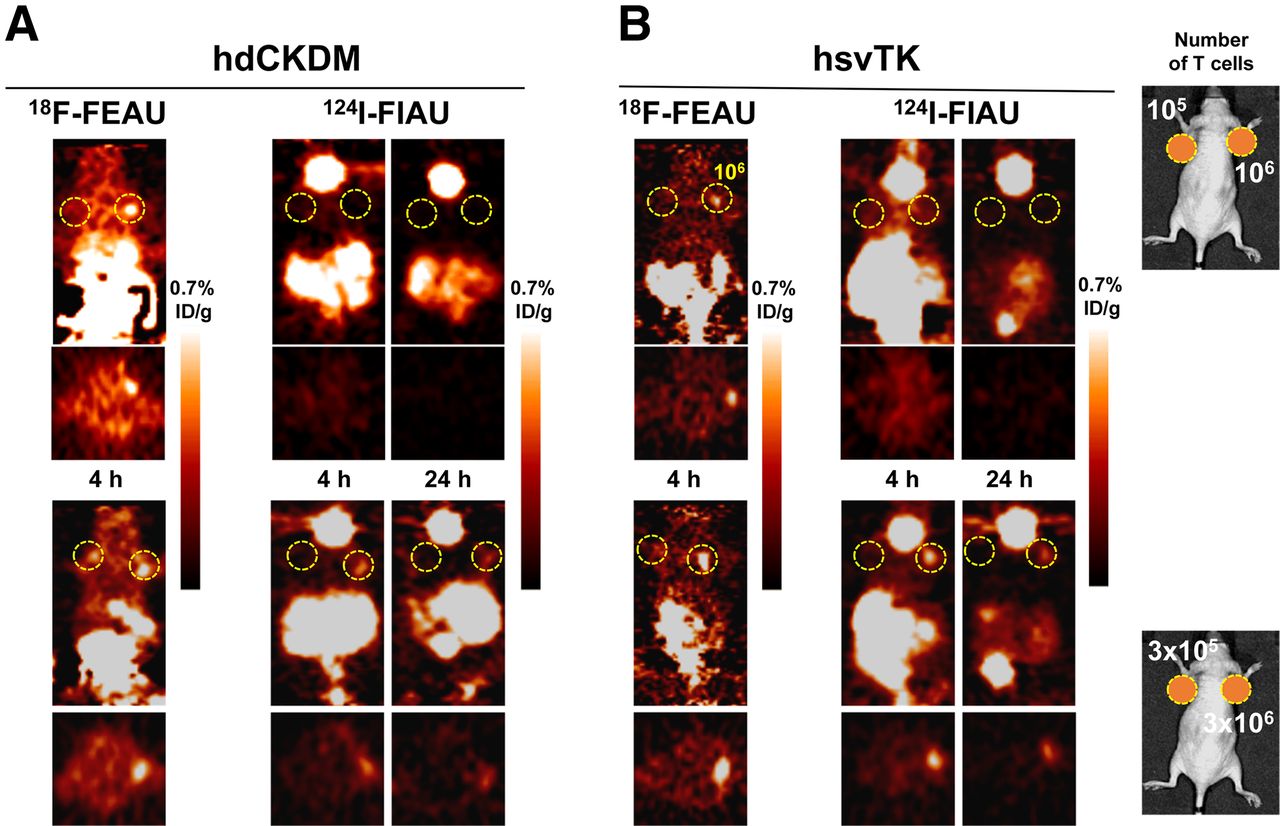
PET imaging of human primary T cells transduced with hdCKDM (A) or hsvTK (B) reporters. Different numbers of T cells were injected subcutaneously, followed by systemic administration of corresponding radiopharmaceuticals and PET imaging at respective time points. Number of T cells injected is shown in right upper and lower panels. %ID/g = percentage injected dose per gram.
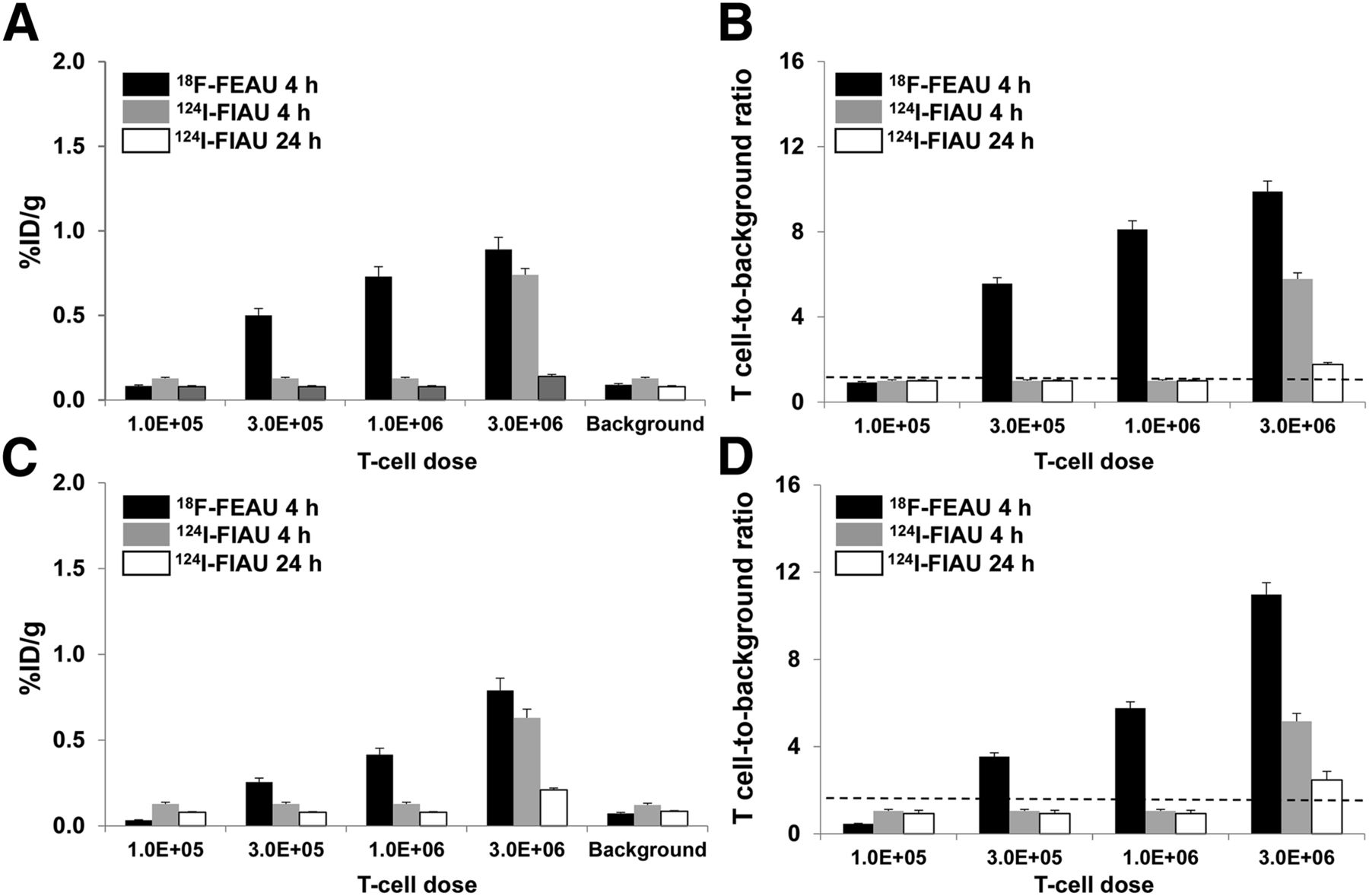
Analysis of PET images of human T cells transduced with hdCKDM (A and B) and hsvTK (C and D), as shown in Figure 4. Data are %ID/g ± SD (A and C) and T cell–to–background ratios to ± SD (B and D). Data are from 2 independent experiments. %ID/g = percentage injected dose per gram.
DISCUSSION
The objective of this report was to determine the minimum number of T cells that can be imaged by different reporter systems (reporter gene + radiolabeled probe) using PET in a murine model. Three human genes (hNET, hNIS, and hdCKDM) and an established viral reporter gene (hsvTK), coupled with their corresponding radiolabeled substrates (comprising different reporter systems), were studied. We compared the sensitivity of the reporter systems to image different numbers of hNET-, hNIS-, hdCKDM-, and hsvTK-transduced T cells at subcutaneous injection sites. All PET reporter systems were effective, but there was a greater than 10-fold difference in threshold sensitivity for T cell detection (Table 1). Not surprisingly, the SPECT reporter system (hNET/123I-MIBG) had more than 100-fold-lower sensitivity than the most sensitive PET reporter system (hNET/18F-MFBG). The hNET/18F-MFBG reporter system could detect less than 1 × 105 T cells, and we were able to visualize all 4 sets of T cells (ranging in number between 1 × 105 and 3 × 106) in all experimental animals (n = 8 per group). The equation describing the relationship between T cell number and measured radioactivity above background levels is T cell number at the injection site = 31,515 × e(1.03 × [measured percentage injected radioactivity/g – background]) (R = 0.80). Thus, approximately 35,000–40,000 hNET reporter T cells can be detected using 18F-MFBG and small-animal PET, 4 h after their subcutaneous injection and intravenous administration of the radiotracer.
This study builds on a recent comparison between 18F-MFBG and clinically approved 123/124I-MIBG for imaging of hNET-expressing cells and tumors (15,25). These studies showed that 18F-MFBG can be synthesized in reasonable radiochemical yields under milder conditions and that it is a more hydrophilic benzylguanidine analog than MIBG, has a lower binding to plasma proteins, and is cleared more rapidly from nontarget tissues and from the body. These characteristics result in superior tumor-to-background images at earlier times after administration of MFBG, compared with MIBG. Although the in vitro experimental data (Fig. 1) and the previously published binding data (15) suggest that MIBG would be a more effective probe than MFBG, the higher logD and serum protein binding and slower body clearance of MIBG, compared with that of MFBG, account for the discordance between the in vitro and in vivo uptake results. These results also corroborate previously published studies using the hNET reporter system in a model of intratumorally injected Epstein-Barr virus–specific T cells (30).
hNIS imaging, after administration of 124I-iodide, demonstrated rapid accumulation and rapid clearance of the probe from the sites of T cell injection, which is similar to the results described by us and others (29,31) in stably transduced xenografts. However, the T cell–to–background ratio was fairly stable from 1 to 4 h due to the rapid clearance of iodide from most tissues (background) as well as from the T cell injection site. The advantage of rapid initial accumulation was negated by the high background level of radioactivity and near-parallel clearance from the T cell injection site and surrounding tissues (background).
PET imaging of hsvTK/GFP and hdCKDM/GFP with 18F-FEAU also yielded positive results for T cell numbers between 3 × 105 and 3 × 106. Similar PET T cell imaging threshold values were obtained with hsvTKsr39/18F-FHBG by Su et al. and Yaghoubi et al. (32,33). HsvTK and human dCKDM reporter imaging with 124I-FIAU and PET at 24 h after injection was only able to reliably visualize the highest number of T cells injected (3 × 106). Only 20% of the animals showed a positive T cell signal after injection of 1 × 106 cells, and no visualization of T cells was observed below 1 × 106 T cells by either hsvTK or hdCKDM imaged with 124I-FIAU.
We and others have previously demonstrated that FEAU has more rapid body clearance, lower background, and higher affinity to and accumulation by HsvTK and hdCKDM enzymes than FIAU (12). These preferential pharmacokinetic and pharmacodynamic parameters resulted in a lower T cell detection threshold with 18F-FEAU than with 124I-FIAU.
The in vivo experiments demonstrated an advantage of short-half-life 18F-based probes that clear nontarget tissue and the body rapidly, including 18F-MFBG and 18F-FEAU for imaging early (i.e., 4 h). In contrast, the more slowly clearing iodine-based substrates, such as 124I-MIBG, 123I-MIBG, and 124I-FIAU, require a longer time to achieve a good target-to-background ratio because the bulky electron-donor iodine is more hydrophobic than the small electron-acceptor fluorine. The hNET/123I-MIBG SPECT study showed the lowest sensitivity, with no visualization of 3 × 106 T cells at both early (2–4 h) and late (8–24 h) time points after subcutaneous injection. An additional hNET/123I-MIBG study, with injection of high numbers of T cells (1 × 107 and 3 × 107), showed reliable detection of T cells only at the highest number injected, demonstrating the limitation of 123I-MIBG SPECT imaging.
CONCLUSION
A comparison of different constitutively expressed radionuclide-based reporter genes (3 human and 1 viral) and their corresponding reporter probes for imaging of T cell number was performed. The hNET/18F-MFBG PET reporter system was the most sensitive, capable of detecting 3–4 × 104 T cells at the site of T cell injection in the animal model. Fast-clearing short-half-life 18F-based probes demonstrate high specificity and allow for repetitive of T cell imaging.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This work was supported by the NIH P50 CA86438-11, R01 CA163980, and R01 CA161138 grants and Mr. William H. Goodwin and Mrs. Alice Goodwin and the Commonwealth Foundation for Cancer Research and The Experimental Therapeutics Center of Memorial Sloan Kettering Cancer Center. Technical services were provided by the MSKCC Small-Animal Imaging Core Facility, supported in part by NIH Small-Animal Imaging Research Program (SAIRP), NIH Shared Instrumentation grant no. 1 S10 RR020892-01, NIH Shared Instrumentation grant no. 1 S10 RR028889-01, and NIH Center grant P30 CA08748. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Dr. Jason Lewis and the Radiochemistry Core for their technical assistance and expertise.
Footnotes
Published online May 29, 2015.
- © 2015 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication April 23, 2015.
- Accepted for publication April 25, 2015.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Imaging cellular immunotherapies and immune cell biomarkers: from preclinical studies to patients
- Development of a Clinically Relevant Reporter for Chimeric Antigen Receptor T-cell Expansion, Trafficking, and Toxicity
- Imaging of T-cell Responses in the Context of Cancer Immunotherapy
- Molecular Imaging with Reporter Genes: Has Its Promise Been Delivered?
- 89Zr-oxine labelling and PET imaging shows lung delivery of a cell/gene cancer therapy
- New Developments in Imaging Cell-Based Therapy
- Reporter Genes for PET Imaging of CAR T Cells Offers Insight into Adoptive Cell Transfer
- Antibody with Infinite Affinity for In Vivo Tracking of Genetically Engineered Lymphocytes
- Norepinephrine Transporter as a Target for Imaging and Therapy
- Molecular Imaging of Inflammation: Current Status
- Dynamic imaging for CAR-T-cell therapy