


Abstract
Serotonin receptor 6 (5-hydroxytrypamine-6, or 5-HT6) is a potential therapeutic target given its distribution in brain regions that are important in depression, anxiety, and cognition. This study sought to investigate the effects of age on 5-HT6 receptor availability using 11C-GSK215083, a PET ligand with affinity for 5-HT6 in the striatum and 5-HT2A in the cortex. Methods: Twenty-eight healthy male volunteers (age range, 23–52 y) were scanned with 11C-GSK215083 PET. Time–activity curves in regions of interest were fitted using a multilinear analysis method. Nondisplaceable binding potential (BPND) was calculated using the cerebellum as the reference region and corrected for partial-volume effects. Results: In 5-HT6–rich areas, regional 11C-GSK215083 showed a negative correlation between BPND and age in the caudate (r = −0.41, P = 0.03) (14% change per decade) and putamen (r = −0.30, P = 0.04) (11% change per decade) but not in the ventral striatum or pallidum. A negative correlation with age was also seen in cortical regions (r = −0.41, P = 0.03) (7% change per decade), consistent with the literature on 5-HT2A availability. Conclusion: To our knowledge, this was the first in vivo study on humans to examine the effect of age on 5-HT6 receptor availability. The study demonstrated a significant age-related decline in 5-HT6 availability (BPND) in the caudate and putamen.
The 5-hydroxytrypamine-6 (5-HT6) receptor is a 7-transmembrane serotonin receptor subtype and one of the 14 distinct mammalian serotonin receptors that are expressed almost exclusively in the central nervous system (1,2). In vitro studies demonstrated that 5-HT6 receptors are localized to the striatum, hippocampus, amygdala, hypothalamus, thalamus, and cerebral cortex (3). The localization to both basal ganglia and limbic structures suggests that this receptor may be involved in the serotonergic control of motor function, mood-dependent behavior, depression, and cognition (3). Subsequently, more attention has recently been given to 5-HT6 because it presents an attractive therapeutic target for neuropsychiatric disorders.
Functionally, 5-HT6 exhibits excitatory action, but it can also colocalize with γ-aminobutyric acid–ergic neurons and produce an inhibition of brain activity leading to complicated and discrepant results (2,4). Heterogeneous effects are also seen with other neurotransmitters in specific brain regions, with 5-HT6 antagonism resulting in increased extracellular glutamate, dopamine, and acetylcholine in the frontal cortex and hippocampus (5,6), whereas 5-HT6 agonists have been shown to produce increased extracellular γ-aminobutyric acid release in the dorsal hippocampus, striatum, and amygdala (4). Thus, with a multitude of interactions not fully elucidated, preclinical studies on both 5-HT6 agonists and 5-HT6 antagonists have shown efficacy against depression, anxiety, and cognitive impairment (1). In addition, 5-HT6 antagonists have shown promise in the treatment of obesity (7), major depressive disorder (8), age-related cognitive decline (9), and schizophrenia (10). Currently available antidepressants and antipsychotics have also been shown to have a strong affinity for the 5-HT6 receptor (11,12), adding speculation that these clinical effects might, at least in part, be mediated through this receptor.
Age has been associated with a decline in receptor density across several 5-HT receptor subtypes (13–15). Given that some of the potential indications, such as cognitive impairment in Alzheimer disease and age-related cognitive decline, are related to advancing age, it is important to understand the effects of age on 5-HT6 receptors and to ascertain whether they remain an important target in older age groups. This importance is further underscored by recent negative results from 2 phase III trials of the 5-HT6 antagonist idalopirdine for cognitive impairment in Alzheimer disease, despite promising results in preclinical studies (16).
Although age has been shown to be associated with a decline in 5-HT1B (14), 5-HT1A, 5 HT2A (15), and 5-HT4 receptors (13), the effects of age on 5-HT6 receptor availability in humans is not known. The availability of a PET ligand for the 5-HT6 receptor 11C-GSK215083 offers an opportunity to examine the in vivo receptor density in healthy humans and in disease states (17,18). In vitro GSK215083 exhibits high binding affinity for 5-HT6 (pKi = 9.8) and 5-HT2A (pKi = 9.1) receptors. Although 11C-GSK215083 has high affinity for 5-HT6 receptors in vivo, it has a nonnegligible affinity (albeit 5-fold lower) for 5-HT2A receptors in cortical regions (17). Therefore, whereas striatal 11C-GSK215083 signal is primarily reflective of 5-HT6 availability, its binding in the cortex is thought to reflect 5-HT2A availability (17,18).
In this paper, we examine the effects of age on 5-HT6 receptor availability in healthy male volunteers using the PET ligand 11C-GSK215083.
MATERIALS AND METHODS
Participants
Twenty-eight healthy male volunteers (mean age, 36.0 ± 9.3 y; range, 23–52 y) were included in the study. All underwent a comprehensive screening assessment that included a clinical interview, complete physical examination with medical history, routine blood tests, electrocardiogram, and urine toxicology. Individuals were excluded if they self-reported or evaluation revealed a diagnosis of a current or lifetime psychiatric disorder; current or past serious medical or neurologic illness (including a history of head injury with loss of consciousness); metal in their body, which would result in MRI exclusion; a history of substance abuse or dependence; or illicit drug use in the previous 3 mo.
The study was performed under protocols approved by the Yale Human Investigation Committee, the Yale University Radiation Safety Committee, the Yale–New Haven Hospital Radioactive Drug Research Committee, and the Yale MRI Safety Committee. The study was performed in accordance with good clinical practice guidelines, regulatory requirements, and the code of ethics of the World Medical Association (Declaration of Helsinki). Subjects were recruited from Connecticut and surrounding states by paper and online advertisements, as well as personal referrals. Written informed consent was obtained from all participants at the beginning of screening after the study procedures had been fully explained.
Radiosynthesis of 11C-GSK215083 on the FXC Automated Module
11C-GSK215083 was radiolabeled with 11C-methyl triflate using an FXC automated module (GE Healthcare) by modifying a previously described procedure (18). 11C-methyl triflate was swept into a solution of desmethyl GSK215083 (0.8–0.9 mg) and 1 μL of 2,2,6,6,-tetramethylpiperidine in 150 μL of anhydrous N,N-dimethylformamide and was cooled at −30°C until radioactivity peaked. The resulting solution was then heated at 85°C for 4 min. The reaction mixture was cooled to 50°C, diluted with 1 ml of deionized water, and injected onto the semipreparative high-performance liquid chromatography column (Gemini C18, 5 μm, 10 × 250 mm; Phenomenex Inc.) eluting at a flow rate of 4 mL/min with a mixture of 40% acetonitrile and 60% 0.05 M ammonium formate adjusted to pH 8.1–8.4 using saturated ammonium hydroxide. The radioactivity fraction eluting between 18 and 19 min was collected, diluted with 50 mL of water, and loaded onto a C18 SepPak classic cartridge (Waters). The cartridge was rinsed with 10 mL of 1 mM HCl and dried. The product was then eluted off the SepPak with 1 mL of ethanol (U.S. Pharmacopeia [USP] grade), followed by 3 mL of saline (USP grade), into the product vessel precharged with a mixture of 7 mL of saline (USP grade) and 20 μL of 4.2% sodium bicarbonate (USP grade). The resulting solution was finally passed through a sterile 0.22-μm membrane filter (Millex-MP, 25 mm; Millipore) into a sterile empty dose-vial. Purity and molar activity were determined using an analytic reversed-phase high-performance liquid chromatography column (Luna C18(2), 5 μm, 250 × 4.6 mm; Phenomenex) eluting with a mixture of 40% acetonitrile and 60% 0.1 M ammonium formate at a flow rate of 1.5 mL/min.
Radiosynthesis on the Bioscan AutoLoop
11C-GSK215083 was also radiolabeled with 11C-methyl iodide using a loop method developed at the Yale PET Center. 11C-methyl iodide, produced by the gas-phase method, was swept through the sample loop preloaded with a solution of 0.5 mg of desmethyl GSK215083 and 1 μL of 2,2,6,6,-tetramethylpiperidine in 80 μL of anhydrous N,N-dimethylformamide until radioactivity peaked. The reaction mixture was allowed to stand for 5 min at ambient temperature, loaded onto the semipreparative high-performance liquid chromatography column, and purified as described as above. The product fraction was collected, diluted with 50 mL of water, and loaded onto a C18 Sep-Pak classic cartridge. The cartridge was rinsed with 10 mL of 1 mM HCl and dried. The product was then eluted off the SepPak with 1 mL of ethanol (USP grade), followed by 3 mL of saline (USP grade). The combined solution was finally passed through a sterile 0.22-μm membrane filter into a sterile dose-vial precharged with 7 mL of USP-grade saline and 20 μL of 4.2% sodium bicarbonate (USP grade). Purity and molar activity were determined using a Luna C18(2) column (5 μm, 250 × 4.6 mm) eluting with a mixture of 40% acetonitrile and 60% 0.1 M ammonium formate at a flow rate of 1.5 mL/min.
Image Acquisition
Structural MRI was performed on a 3-T Trio system (Siemens Medical Solutions) with a circularly polarized head coil for purposes of excluding individuals with anatomic abnormalities and anatomically coregistering with PET scans. The dimensions and voxel size of MR images were 256 × 256 × 176 voxels and 0.98 × 0.98 × 1.0 mm, respectively.
Subjects underwent 90-min dynamic PET on an ECAT EXACT HR+ scanner (Siemens Medical Systems) in 3-dimensional mode. Before each PET scan, a transmission scan was acquired for attenuation correction. Up to 740 MBq (20 mCi) of radioactivity were administered by an infusion pump for each scan (mean, 585.39 ± 128.34 MBq). The maximum mass dose from the radioligand was limited to minimize any self-occupancy (≤10 μg) (mean, 3.03 ± 2.32 μg). For each PET scan, 11C-GSK215083 was administered by bolus intravenous infusion over 60 s, immediately followed by acquisition of emission data for up to 90 min.
Arterial input function was measured using an automated blood counter (PBS-101; Veenstra Instruments) for the first 7 min and 15 manual samples between 3 and 90 min after injection. Whole-blood samples were assayed for radioactivity using 0.2 mL, and the rest of the samples were then centrifuged at 2,930g for 5 min to obtain plasma. The radioactivity in all samples was assayed using cross-calibrated γ-counters (1480 Wizard; Perkin-Elmer). Plasma samples collected at 5, 15, 30, 60, and 90 min after injection were mixed with urea to a final concentration of 8 M and were subsequently filtered with 1.0-μm GE Whatman 13-mm CD/X filters. The filtrate was then analyzed with the automatic column-switching high-performance liquid chromatography method (19) to determine the parent fraction and 11C-GSK215083 metabolites.
Metabolite Analysis
Injected samples were first eluted through a capture column hand-packed with C18 sorbent (Strata-X; Phenomenex) using a mobile phase of 1:99 MeCN:H2O (v/v) at 2 mL/min for 4 min. The capture column was then backflushed onto the analytical column (Phenomenex Luna C18(2), 4.6 × 250 mm, 5 μm) with a mobile phase of 40:60 MeCN:0.1 M ammonium formate at a flow rate of 1.4 mL/min, giving a retention time of 8.5 min for parent 11C-GSK215083. Eluent was collected in separate vials every 2 min with a fraction collector (CF-1; Spectrum Chromatography) and counted with γ-counters. The time course of the unchanged (i.e., parent) fraction in the filtrate was fit with an integrated γ-function. This fraction curve was corrected for the time-varying extraction efficiency of radioactivity. The final plasma input function was calculated as the product of the total plasma time–activity curve and the parent fraction curve. Plasma free fraction was assessed by ultrafiltration (Centrifree; MilliporeSigma). The plasma free fraction was determined as the ratio of the radioactivity concentration in the filtrate to the total activity in plasma. Measurements of free fraction were performed in triplicate for each scan.
Image Processing
Dynamic scan data were reconstructed with corrections for attenuation, normalization, scatter, random events, and dead time using ordered-subsets expectation maximization (4 iterations, 16 subsets). PET images were reconstructed into 27 frames containing 63 axial slices of 128 × 128 voxels each (2.1 × 2.1 × 2.4 mm). Motion correction was applied to the dynamic images using a mutual-information algorithm (FSL-FLIRT, version 3.2; FMRIB Analysis Group) by frame-by-frame registration to a summed image (0–10 min after injection). A summed image was created from the motion-corrected data and registered to the subject’s 3-T MR anatomic image (6-parameter affine registration), which was then registered to an MRI template using a nonlinear transform with BioImage Suite. Regions of interest (ROIs) were based on anatomic automatic labeling for SPM2 (20) with the exception of a ventral striatum ROI that was drawn manually on each individual MR image (21,22)). Time–activity curves were then generated in all ROIs.
Time–activity curves were fitted with the MA1 (multilinear analysis 1) method of Ichise et al. (23) (t* = 40) using the metabolite-corrected arterial input function over 90 min after injection. Volumes of distribution were estimated for the amygdala, cerebellum, caudate, hippocampus, putamen, pallidum, and ventral striatum and for a global cortical region consisting of the frontal, parietal, temporal, and occipital cortices. Regional nondisplaceable binding potential (BPND) was calculated from distribution volumes using cerebellum as the reference region because it is virtually devoid of 5-HT2A (24) and 5-HT6 receptors (25).
Partial-volume correction based on the Mueller-Gartner algorithm (26) was applied after segmentation of MR images using SPM12. The uncorrected and corrected data were analyzed separately.
Because striatal 11C-GSK215083 BPND primarily reflects 5-HT6 availability, a linear mixed model (SPSS Statistics for Macintosh, version 22.0; IBM) was used to model the independent and joint effects of age (continuous) and striatal ROIs (i.e., caudate, putamen, and ventral striatum) (within-subject factor) on BPND, with intercept as a random effect, a scaled identity covariance structure, and relevant covariates of body mass index (BMI) and mass dose (μg). The interaction between region and age was modeled, and slopes for regions were estimated post hoc. Correlations between age and BPND were calculated separately for each region, and P values unadjusted for multiple comparisons are reported.
RESULTS
Table 1 shows the demographics of the study sample.
Demographics of Sample
The results of the primary mixed-model analysis on striatal ROIs (uncorrected BPND), with BMI and mass dose as covariates, showed an overall significant effect of age (F(1,24) = 7.39, P = 0.012), ROIs (F(2,52) = 5.35, P = 0.008) and an age × ROI interaction (F(2,52) = 5.07, P = 0.010). There was no main effect of BMI or mass dose. Similarly, after partial-volume correction, analysis with BMI and mass dose as covariates showed a significant effect of age (F(1,24) = 5.93, P = 0.02) and ROI (F(2,52) = 10.67, P < 0.001) but no age × ROI interaction (F(2,52) = 2.83, P = 0.07). There was also no main effect of either BMI or mass dose with partial-volume correction.
To verify that age effect on BPND was not confounded by age-related changes in the reference region, we examined the correlation between age and cerebellar distribution volume and found no significant correlation (r = 0.04, P = 0.85).
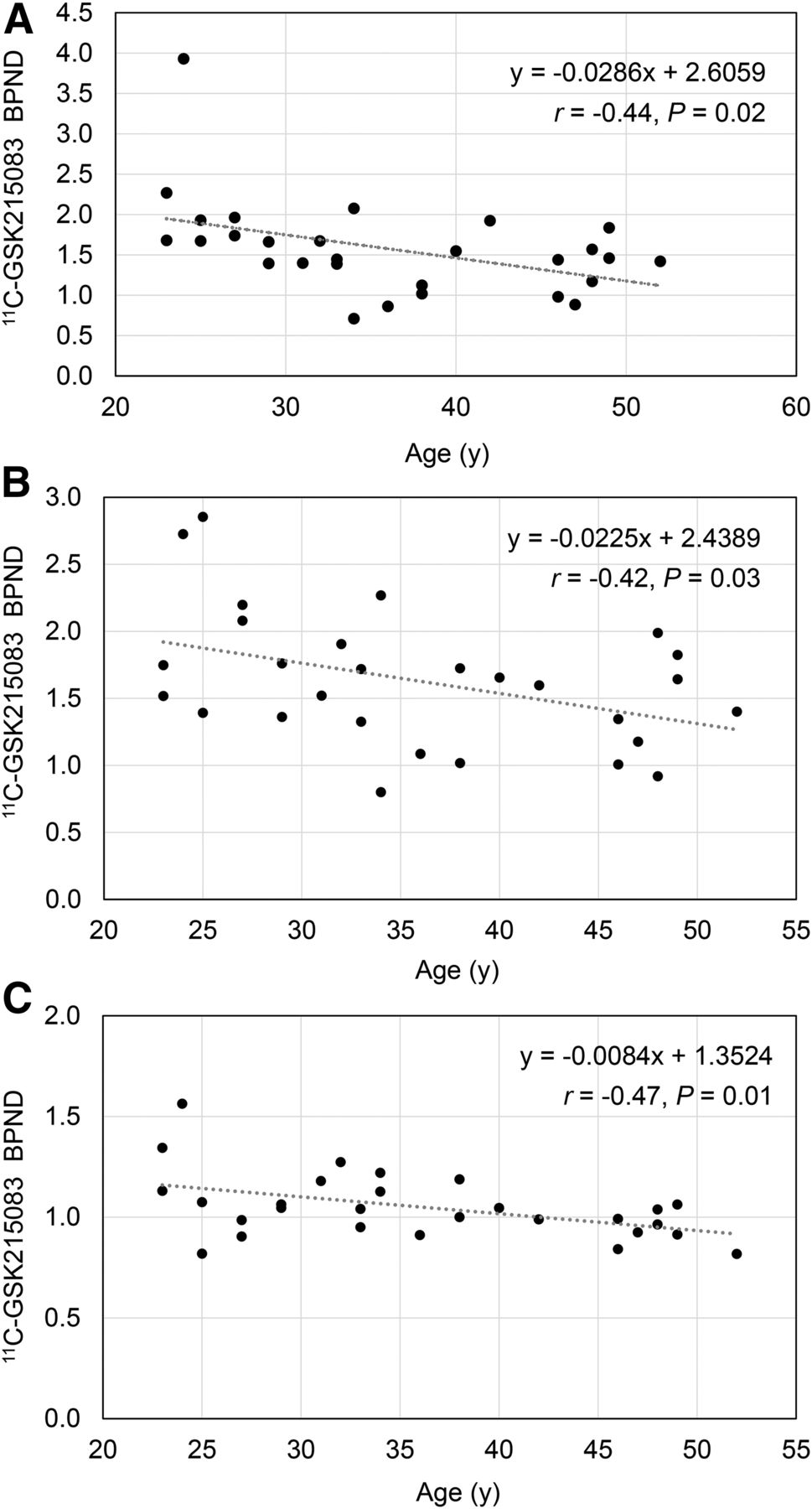
Table 2 presents average 11C-GSK215083 BPND (with SD) for ROIs, along with the correlation with age, the percentage change per decade, slope (change in BPND per year), and the unadjusted P value. BPND was comparable to that estimated using MA2 by Parker et al. (18). 11C-GSK215083 BPND correlated negatively with age in the caudate (corrected: r = −0.44, P = 0.02; uncorrected: r = −0.62, P = 0.001), putamen (corrected: r = −0.42, P = 0.03; uncorrected: r = −0.44, P = 0.02), and cortical regions (corrected: r = −0.47, P = 0.01; uncorrected: r = −0.62, P = <0.001) but not in the amygdala, hippocampus, ventral striatum, or pallidum (Table 2; Fig 1). The reduction per decade was 18% in the caudate, 10% in the putamen, and 12% in cortical regions with the uncorrected BPND data. Likewise, with the corrected BPND data, similar reductions were found: 14% in the caudate, 11% in the putamen, and 7% in cortical regions.
Regional 11C-GSK215083 BPND and Correlation with Age

Correlation between age and 11C-GSK215083 BPND in caudate (A), putamen (B), and cortical (C) regions.
DISCUSSION
To our knowledge, this was the first in vivo study on humans to examine the effects of age on 5-HT6 receptor availability (BPND) using 11C-GSK215083 PET imaging. The study demonstrated the presence of a significant age-related decline in 5-HT6 availability in the caudate and putamen but not in the amygdala, hippocampus, ventral striatum, or pallidum. The reduction per decade was greatest in the caudate (14%) followed by the putamen (11%) in corrected data. 11C-GSK215083 binding in cortical regions, reflective of 5-HT2A availability, also showed an age-related decline (7% per decade).
Consistent with a prior autoradiography study on humans (11) and a PET imaging study using 11C-GSK215083 (18), 5-HT6 availability was found to be higher in striatal than nonstriatal regions. 11C-GSK215083 has significant affinity for 5-HT2A receptors in cortical regions (17). In a series of displacement studies using a 5-HT2A–selective antagonist (ketanserin) and a dual 5-HT6/5-HT2A antagonist (SB742457), Parker et al. (18) demonstrated that cortical 11C-GSK215083 BPND was primarily reflective of 5-HT2A availability.
The finding of an age-related decline in 5-HT6 receptor availability is largely consistent with other 5-HT receptors, namely 5-HT1A, 5-HT1B, 5-HT2A, and 5-HT4 (13,14,27,28), although there are regional differences. The age-related decline in cortical 11C-GSK215083 BPND (reflective of 5-HT2A receptor availability) is consistent with previous PET imaging studies using other selective or nonselective radiotracers (15,28). Moses-Kolko et al. (15) found an 8% decline per decade in 5-HT2A availability using 18F-altanserin, a finding that is strikingly similar to our finding of a 7% decline per decade in corrected data. A similar pattern of age-related decline in cortical areas has also been seen with 5-HT1A (15) and 5-HT1B receptors (14).
The region-specific age-related decline in the caudate and putamen, but not in the amygdala, hippocampus, ventral striatum, or pallidum, is a unique finding. This result persisted despite controlling for potential confounders such as BMI and mass dose and correcting for partial-volume effects. We also performed a secondary analysis excluding outliers (defined as BPND > 3 SDs from mean). The BPND of 3.9 in the caudate was more than 3 SDs from the mean, and when this value was excluded, the overall results in the caudate were similar, demonstrating a significant age-related decline (y = −0.0170 + 2.1106; r = −0.39, P = 0.041).
Among the 5-HT receptor subtypes, age-related decline in the caudate and putamen has been observed with 5-HT2A (28) and 5-HT4 (exclusively in the striatum; not in cortical areas) (13), whereas 5-HT1B showed an increase in availability with age in the pallidum and putamen (14).
An age-related decline in 5-HT2A receptor availability has been shown in the hippocampus and amygdala (15). Because both 5-HT2A and 5-HT6 receptors are expressed in the amygdala (29,30) and hippocampus (31,32), the absence of an age effect in these ROIs in our study could be confounded by the relative proportion of 5-HT6 and 5-HT2A receptors, low BPND, or high variability in these small ROIs. Therefore, specific conclusions on changes in 5-HT6 receptor availability with age in these regions cannot be drawn from our present results.
The findings of the study must be viewed within the context of its limitations. Our sample comprised male subjects who were younger than 60 y, with a mean of 36.0 ± 9.3 y. The sample thus does not capture age effects in subjects older than 60 y, and it would be important to include older subjects in future studies.
Another limitation is that the ligand, GSK215083, has high affinity for both 5-HT2A and 5-HT6 receptors. It is therefore difficult to ascribe a change in tracer binding to one specific receptor type. To more accurately estimate the effect of age on 5-HT6 receptor density with 11C-GSK215083, a blocking study could be done with a selective 5-HT2A antagonist. A previous study showed that ketanserin, a 5-HT2A antagonist, reduced only 3% of 11C-GSK215083 signal in the putamen and 16% in the caudate (18). On the basis of this finding, the signal in the putamen and caudate in the current work is thought to reflect relatively specific binding for the 5-HT6 receptor.
Additionally, whereas the MA2 reference model is a validated method for the ligand (18), the MA1 reference model was found to be a good fit for time–activity curves in our sample, in part, by only fitting data for times longer than 40 min. There were no significant age effects on cerebellar distribution volume, and therefore the use of cerebellum as a reference region does not confound the results. The age-related decline in 5-HT6 receptor availability in caudate and putamen is an important consideration for drug development aimed at targeting this receptor in the treatment of geriatric depression, age-related cognitive decline, and movement disorders.
CONCLUSION
This in vivo study on humans examined the effect of age on 5-HT6 receptor availability and found a significant age-related decline in 5-HT6 availability (BPND) in the caudate and putamen.
DISCLOSURE
This work was supported by Abbvie and Pfizer. Rajiv Radhakrishnan is supported by the David Mahoney grant program of the Dana Foundation and by CTSA grant UL1 TR001863 from the National Center for Advancing Translational Science (NCATS), components of the National Institutes of Health (NIH), and the NIH Roadmap for Medical Research. No other potential conflict of interest relevant to this article was reported. The contents of this publication are solely the responsibility of the authors and do not necessarily represent the official view of NIH.
Acknowledgments
We thank the staff of the Yale PET Center, the Clinical Neuroscience Research Unit (CNRU) at the Connecticut Mental Health Center (CMHC) of the Connecticut Department of Mental Health and Addiction Services (DMHAS), the Hospital Research Unit (HRU) at Yale–New Haven Hospital (YNHH), and the Yale Magnetic Resonance Research Center (MRRC).
Footnotes
Published online Apr. 6, 2018.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication December 4, 2017.
- Accepted for publication March 21, 2018.





{kind=link}
Jump to section
Related Articles
Cited By...
- Brain maps of general cognitive function and spatial correlations with neurobiological cortical profiles
- A replicable and generalizable neuroimaging-based indicator of pain sensitivity across individuals
- Resting state changes in aging and Parkinsons disease are shaped by underlying neurotransmission - a normative modeling study
- Transitions between cognitive topographies: contributions of network structure, neuromodulation, and disease