


Abstract
18F-BCPP-EF was applied to assess mitochondrial complex I (MC-I) activity in the brains of Parkinson disease model monkeys prepared by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and also presynaptic dopamine parameters. Methods: 11C-β-CFT for the dopamine transporter; 11C-3,4-dihydroxy-phenyl-L-alanine (β-11C-L-DOPA), L-6-18F-fluorodopa (18F-FDOPA), or 6-11C-methyl-m-tyrosine (11C-6MemTyr) for dopamine synthesis; or 2-tert-butyl-4-chrolo-5-{6-[2-(2-18F-fluoroethoxy)-ethoxy]-pyridin-3-ylmethoxy}-2H-pyridazin-3-one (18F-BCPP-EF) for MC-I was intravenously injected into normal and MPTP monkeys in order to analyze their uptake in the striatum. Results: Significant reductions in presynaptic dopamine parameters and MC-I activity were detected in the striatum of MPTP monkeys. Correlations were observed between MC-I activity and dopamine transporter as well as between MC-I activity and dopamine synthesis in the striatum. The order of detectability of impaired MC-I activity was 11C-6MemTyr >> β-11C-L-DOPA > 18F-FDOPA. Conclusion: 18F-BCPP-EF has potential as a PET probe for the quantitative imaging of MC-I damage in the living brains of Parkinson disease model monkeys using PET.
Parkinson disease (PD) is characterized by the progressive degradation of nigrostriatal pathways with the selective loss of dopaminergic neurons in the substantia nigra pars compacta, resulting in movement disorder such as resting tremor, akinesia, bradykinesia, rigidity, and postural instability. These clinical symptoms of PD were previously reported to be induced after 40%–50% loss of the neurons in the substantia nigra pars compacta; there was also a reduction of dopamine to about 20% of normal levels in the striatum (1). Although the direct cause of the selective neurodegeneration in PD has not yet been identified, mitochondrial dysfunction has emerged as a common aspect of its pathogenesis (2,3). In mammalian cells, the electron transport chain in mitochondria consists of 5 complexes (from I to V) that produce adenosine triphosphate, with complex I (MC-I) being the first and rate-limiting step of overall respiratory activity and oxidative phosphorylation under physiologic conditions. Impaired electron transport and oxidative phosphorylation due to MC-I deficiency may account for the neuronal cell death in PD. The loss of MC-I catalytic activity has been detected in multiple tissues from sporadic PD patients with increased oxidative stress (3).
The exposure of humans to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) induces a syndrome that mimics the core neurologic symptoms of PD and relatively selective dopamine neurodegeneration in the substantia nigra pars compacta (4). MPTP is metabolized by monoamine oxidase B located in glia cells, to 1-methyl-4-phenylpyridinium ions, which are selectively taken up through the dopamine transporter (DAT) and stored in vesicles via uptake by vesicular monoamine transporter 2. These ions then induce their toxic effects in dopaminergic neurons by inhibiting adenosine triphosphate synthesis and generating free radicals.
PET has been applied to the quantitative and noninvasive imaging of biologic functions, and several abnormalities in the nigrostriatal dopamine pathway have been imaged in PD patients using PET (5). In addition to dopamine, we recently designed a PET probe, 2-tert-butyl-4-chrolo-5-{6-[2-(2-18F-fluoroethoxy)-ethoxy]-pyridin-3-ylmethoxy}-2H-pyridazin-3-one (18F-BCPP-EF), for the quantitative imaging of MC-I in vivo and confirmed that it was suitable for imaging MC-I function in the living brains of rats (6) and monkeys (7,8).
In the present study, we prepared an MPTP-induced PD model of cynomolgus monkeys (Macaca fascicularis) (9,10), and 18F-BCPP-EF was evaluated based on its ability to detect changes in MC-I activity in the living brain after a treatment with MPTP. Presynaptic dopamine damage was monitored using 11C-β-CFT for DAT and 11C-3,4-dihydroxy-phenyl-L-alanine (β-11C-L-DOPA), L-6-18F-fluorodopa (18F-FDOPA), and 6-11C-methyl-m-tyrosine (11C-6MemTyr) for dopamine synthesis, and the relationship between presynaptic dopaminergic damage and impaired MC-I activity was also assessed.
MATERIALS AND METHODS
Animals
Animals were maintained and handled in accordance with the recommendations of the U.S. National Institutes of Health and the guidelines of the Central Research Laboratory, Hamamatsu Photonics. The following experiments were approved by the Ethical Committee of the Central Research Laboratory, Hamamatsu Photonics.
Five male normal and 5 MPTP-treated cynomolgus monkeys (Macaca fascicularis; age range, 2.8–3.3 y; weight range, 2.0–3.5 kg) were subjected to PET scans (9,10). MPTP at doses ranging from 0.2 to 0.4 mg/kg of free base in phosphate-buffered saline was injected intravenously once per week over a 4-mo period until a stable parkinsonian syndrome was observed. The total doses of MPTP administered ranged from 1.5 to 3.0 mg/kg. To avoid the possibilities of spontaneous recovery and direct inhibition of 18F-BCPP-EF binding to MC-I by MPTP, MPTP administration was halted for 2 mo, from the last MPTP treatment to the first PET measurements.
MR images of the monkeys were obtained with a 3.0-T MR system (Signa Excite HDxt 3.0T; GE Healthcare) under pentobarbital anesthesia.
PET Ligand Syntheses
11C-β-CFT was labeled by the 11C-N-methylation of its nor-compound with 11C-methyl iodide (11). Radioactive purity was greater than 98%, and specific radioactivity was 42.0 ± 7.9 GBq/μmol. β-11C-L-DOPA was synthesized using a combination of organic synthesis and multienzymatic procedures (12). Radiochemical purity was better than 98%, and specific radioactivity was 25.4 ± 8.5 GBq/μmol. 18F-FDOPA was labeled using chiral phase-transfer alkylation (13). Radioactive and enantiomeric purities were greater than 99% and 92%, respectively. Specific radioactivity was 50.5 ± 13.5 GBq/μmol. 11C-6MemTyr was labeled using the rapid Pd(0)-mediated cross-coupling reaction of 11C-methyl iodide and corresponding alkenyl boronate precursor (10,14). Radiochemical purity was greater than 99%, and specific radioactivity was 40.0 ± 10.1 GBq/μmol. 18F-BCPP-EF was radiolabeled by the nucleophilic 18F-fluorination of the corresponding precursor (6–8). Radiochemical purity was greater than 98%, and specific radioactivity was 139.6 ± 37.0 GBq/μmol.
PET Measurements
After overnight fasting, monkeys were tracheostomized, immobilized with pancuronium bromide (0.05 mg/kg intramuscularly), and artificially ventilated (Cato; Drager). Anesthesia was continued with 0.4–0.8 vol% isoflurane in a N2O/O2/N2 (1:1:1) gas mixture. A venous cannula for PET probe injection and an arterial cannula for blood sampling (only for 18F-BCPP-EF measurements) were inserted in both inferior limbs. The animal’s head was set in a supine position on a bed with stereotactic coordinates aligned parallel to the orbitomeatal line plane. In PET measurements using β-11C-L-DOPA, 18F-FDOPA, and 11C-6MemTyr, carbidopa, an inhibitor of peripheral aromatic L-amino acid decarboxylase, was administered at a dose of 5 mg/kg, by mouth, 1 h before each PET probe injection. After a transmission scan, emission scans were obtained for 91 min after the injection of each PET probe using an animal PET scanner (SHR-7700; Hamamatsu Photonics) (15). PET data were reconstructed by the filtered backprojection method, and individual PET and MR images were coregistered. Volumes of interest in brain regions were drawn manually on MR images by referring to regional information from BrainMaps.org (16).
PET Data Analysis
In the quantitative analysis of 18F-BCPP-EF, arterial blood samples were obtained, centrifuged to separate plasma, and weighed, and their radioactivity was measured. In the metabolite analysis, methanol was added to some plasma samples (sample/methanol = 1/1), which were centrifuged, and the supernatants obtained were developed using thin-layer chromatography plates (AL SIL G/UV; Whatman) with a mobile phase of ethyl acetate. The ratio of unmetabolized fraction (Rf = 0.66) was determined using a phosphoimaging plate (FLA-7000; Fuji Film). The input function of unmetabolized 18F-BCPP-EF was calculated using the data obtained by correction of the ratio of the unmetabolized fraction to total radioactivity, which was used as the arterial input function. A kinetic analysis of 18F-BCPP-EF was performed using the Logan graphical analysis (7,8).
The quantitative analysis of 11C-β-CFT was performed with a simplified reference tissue model to calculate the nondisplaceable binding potential (BPND) (17) using the time–activity curve in the cerebellum as an input function. Quantitative analyses of β-11C-L-DOPA, 18F-FDOPA, and 11C-6MemTyr were conducted by a multitime graphical analysis (MTGA) (10,18,19) applying the time–activity curve in the cerebellum as an input function.
The correlation analyses against MC-I activity were conducted using right and left striatal regions separately from 1 monkey. Because some PET measurements and quantitative analyses failed, the final number of monkeys used in the analyses was 4 for control and 4 for MPTP-treated.
Statistical Analysis
Results are expressed as mean ± SD. Comparisons between conditions were performed using the paired, 2-tailed Student t test, and correlations were tested by a single regression analysis using the Kaleidagraph program (Synergy Software). A probability level of less than 5% (P < 0.05) was considered to indicate statistical significance.
RESULTS
Figure 1 represents the typical PET images of a normal monkey and an MPTP-treated monkey. As shown in the top panels, PET measurements of DAT availability (11C-β-CFT [Fig. 1A]) and dopamine synthesis (β-11C-L-DOPA [Fig. 1B], 18F-FDOPA [Fig. 1C], and 11C-6MemTyr [Fig. 1D]) clearly imaged the striatum of the normal monkey brain. The cerebral uptake of 18F-BCPP-EF in normal monkeys showed homogeneous and symmetric patterns in the 2 hemispheres and enabled discrimination of the cortical and basal ganglion regions (Fig. 1E).
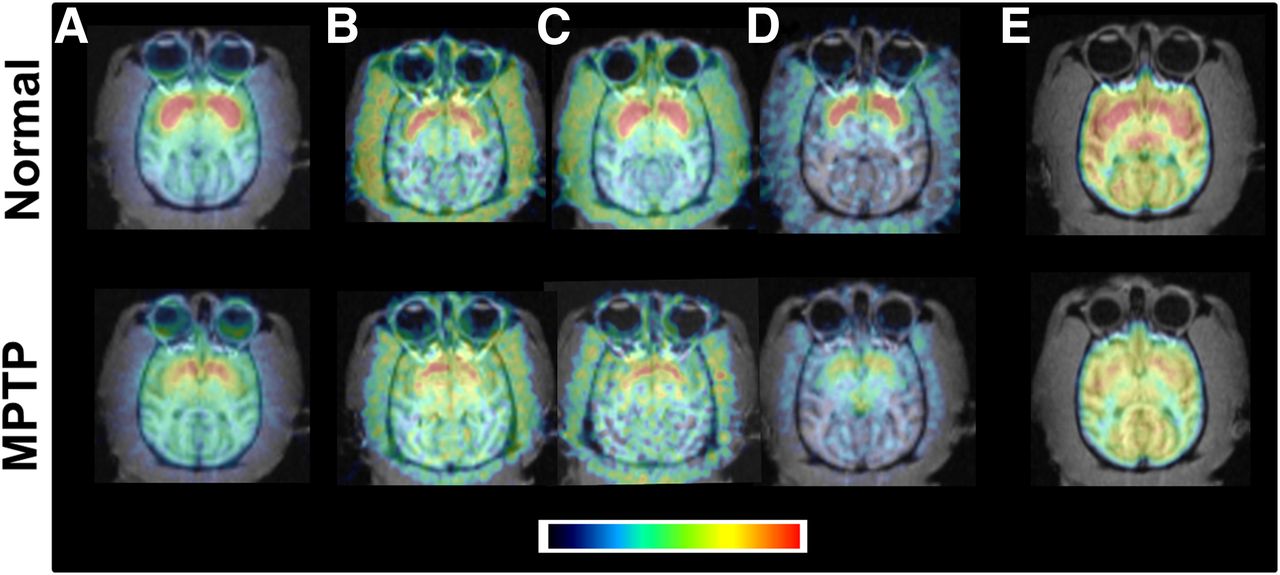
Effects of treatment with MPTP on postsynaptic dopamine parameters and MC-I activity in striatum of monkey brain. Typical PET images of normal monkey (top) and MPTP-treated monkey (bottom) are presented. PET scans for DAT (A; 11C-β-CFT) and dopamine synthesis (β-11C-L-DOPA [B], 18F-FDOPA [C], and 11C-6MemTyr [D]) were acquired for 91 min without arterial blood sampling. PET scans for MC-I (E; 18F-BCPP-EF) were obtained for 91 min with sequential arterial blood sampling.
Presynaptic DAT availability in the striatum, which was assessed using 11C-β-CFT, was markedly lower in MPTP monkeys than in normal animals (bottom panels of Fig. 1A). In addition, dopamine synthesis in the striatum, as evaluated using β-11C-L-DOPA (Fig. 1B), 18F-FDOPA (Fig. 1C), and 11C-6MemTyr (Fig. 1D), was also significantly lower in MPTP monkeys than in normal animals (66.7%, 83.1%, and 46.6%, respectively, of the corresponding normal).
The metabolic rates of 18F-BCPP-EF in plasma were rapid; only 52%, 19%, and 10% of nonmetabolites were detected 10, 30, and 60 min, respectively, after the injection without formation of any lipophilic metabolites. The striatal total distribution volume (Vt) of 18F-BCPP-EF was significantly lower in MPTP monkeys than in normal animals (71.7% that of the corresponding normal) (Fig. 1E). The plot of the Vt of 18F-BCPP-EF against the BPND of 11C-β-CFT revealed a positive correlation (R2 = 0.673, P < 0.001) (Fig. 2A). Although the plot of the Vt of 18F-BCPP-EF against the MTGA metabolic constant (Ki) values of β-11C-L-DOPA, 18F-FDOPA, and 11C-6MemTyr showed positive correlations in the striatum, 11C-6MemTyr (R2 = 0.834, P < 0.001) provided greater correlation coefficients than β-11C-L-DOPA (R2 = 0.424, P < 0.05) and 18F-FDOPA (R2 = 0.550, P < 0.01) (Fig. 2B).
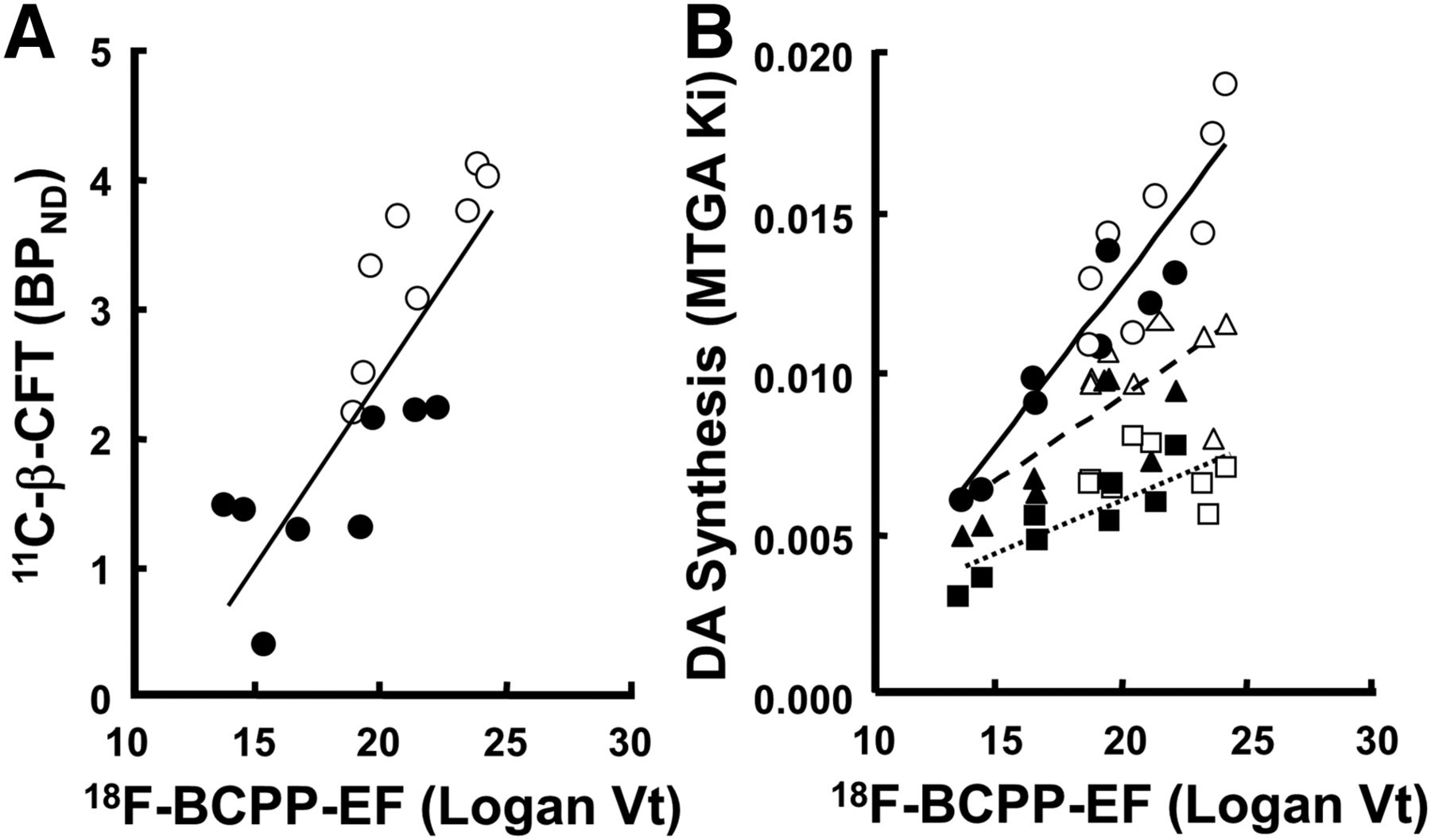
Relationships between Vt of 18F-BCPP-EF and BPND of 11C-β-CFT (○,●) (A) and between MTGA Ki values of 18F-FDOPA (□,▪▪▪), β-11C-L-DOPA (△,), and 11C-6MemTyr (○,●) (B) in striatum of living monkey brain. These correlation analyses involved normal (open symbols) and PD animals (closed symbols). Correlation analyses against MC-I activity were conducted using imaging results from 1 of 2 striata from single animal (8 monkeys total; 4 controls and 4 MPTP-treated). DA = dopamine synthesis.
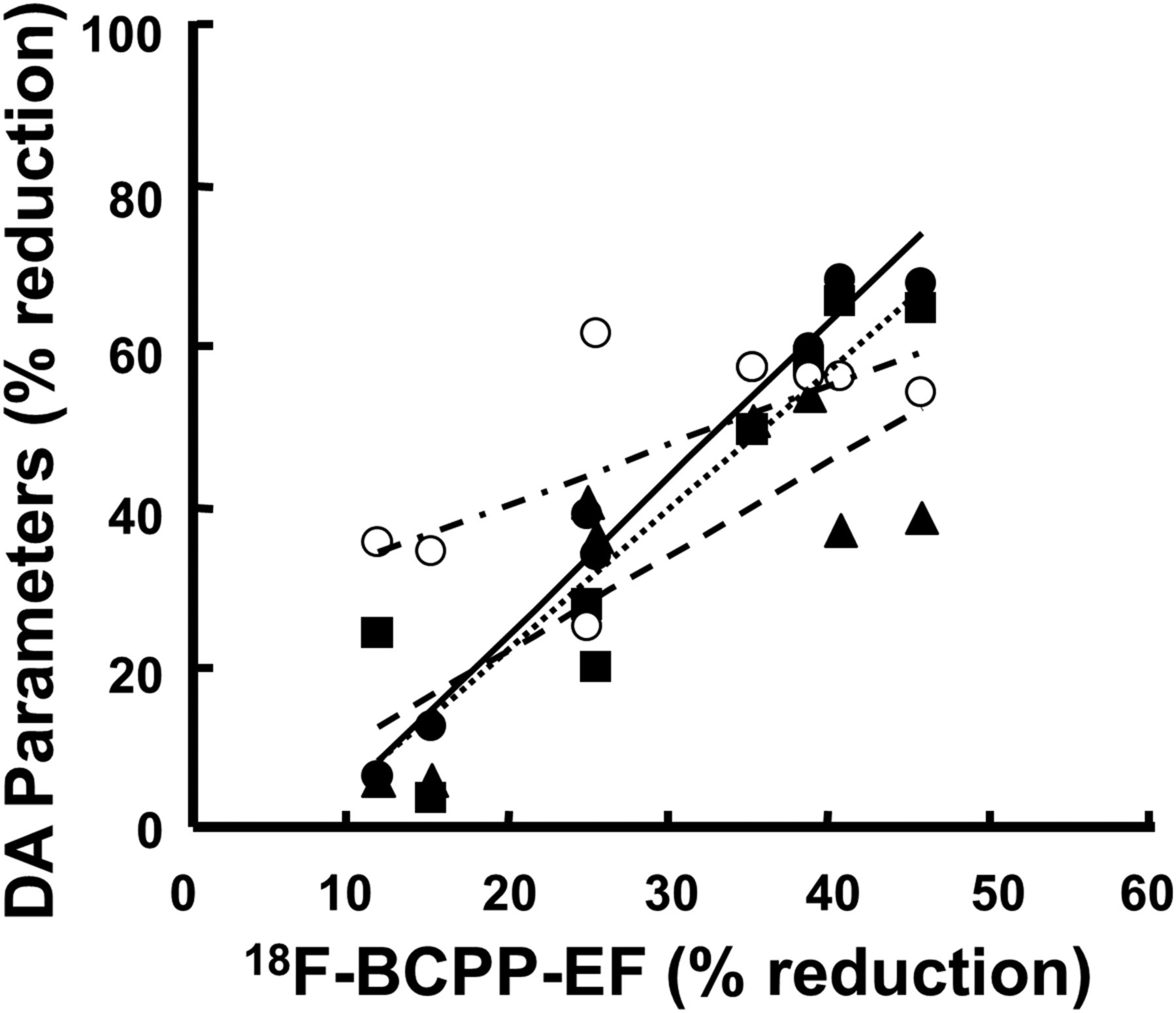
As shown in Figure 3, the correlation analyses between MPTP-induced percentage reductions of dopamine parameters and percentage reduction of 18F-BCPP-EF in the striatum of the living monkey brain revealed that dopamine synthesis rates assessed using 11C-6MemTyr (R2 = 0.974, P < 0.001), 11C-L-DOPA (R2 = 0.634, P < 0.01), and 18F-FDOPA (R2 = 0.844, P < 0.001) were a much better correlation that that of DAT measured using 11C-β-CFT (R2 = 0.451, P < 0.05).

Relationships between MPTP-induced percentage reduction of 11C-6MemTyr (●), 11C-L-DOPA (▪▪▪), 18F-FDOPA (▪▪▪), or 11C-β-CFT (○) against percentage reduction of 18F-BCPP-EF in striatum of living monkey brain. Correlation analyses against MC-I activity were conducted using imaging results from 1 of 2 striata from single animal (8 monkeys total; 4 controls and 4 MPTP-treated). Degrees of each percentage reduction were calculated using mean values of normal animals for each PET probe. DA = dopamine synthesis.
DISCUSSION
The present study demonstrated the ability of 18F-BCPP-EF as a PET probe to detect MPTP-induced changes in MC-I activity in parallel with assessments of presynaptic dopamine parameters. The most remarkable results of the present study was that 18F-BCPP-EF detected MPTP-induced impairments in MC-I activity in the living brain of PD model monkeys using PET. MC-I is the first and rate-limiting step of overall respiratory activity and oxidative phosphorylation from MC-I to MC-V, and impaired electron transport and oxidative phosphorylation due to MC-I deficiency may account for the neuronal cell death in PD. A small reduction in MC-I activity of approximately 25% is sufficient to decrease adenosine triphosphate synthesis and mitochondrial respiration in brain mitochondria. MC-I dysfunction in PD patients has been identified in mitochondria obtained from postmortem brains and biopsy tissue samples (2,3); however, MC-I dysfunctions in the living brain had not been demonstrated noninvasively before the present study.
As we have reported previously, 18F-BMS-747158-02 developed for myocardial imaging (20) is not suitable for MC-I imaging in the living brain because of its high affinity and high lipophilicity (6). 18F-fluorobenzyl triphenylphosphonium cation was reported to bind to mitochondria, reflecting their membrane potential with voltage-dependency (21); however, this PET probe has not been applied for imaging of mitochondrial function in the living brain. In contrast, our previous reports demonstrated the first PET probe as 18F-BCPP-EF for MC-I imaging in the living brain (6–8).
Furthermore, correlations were observed between MC-I activity and DAT availability and also between MC-I activity and dopamine synthesis in the striatum. We previously confirmed that the degeneration of nigrostriatal dopamine neurons was induced by the intravenous administration of MPTP to monkeys over a period of several months and resulted in impaired presynaptic dopamine parameters, as determined by DAT availability, dopamine synthesis, and tyrosine hydroxylase immunoreactivity in the brain (9,10). A previous study reported that although plotting the striatal MTGA Ki values of 11C-6MemTyr, β-11C-L-DOPA, and 18F-FDOPA against the BPND of 11C-β-CFT in the striatum of monkeys revealed positive correlations, the Ki value of 11C-6MemTyr provided greater correlation coefficients than β-11C-L-DOPA and 18F-FDOPA (10). The present study also demonstrated that the most sensitive PET probe for the detection of impaired MC-I activity was 11C-6MemTyr. The different abilities of β-11C-L-DOPA, 18F-FDOPA, and 11C-6MemTyr to detect presynaptic dopamine damage may be attributed to their different metabolic profiles on the basis of catechol- (β-11C-L-DOPA and 18F-FDOPA) and noncatechol-type (11C-6MemTyr) structures (10).
In the nigrostriatal regions containing high dopaminergic neurons, PET imaging using dopamine-specific probes is useful for diagnostic and treatment efficacy assessment of PD patients (5). In contrast, several recent studies suggested that other monoaminergic neurons, such as serotonin and noradrenalin, may be affected in nondopaminergic brain regions of PD patients (22,23), indicating that multiparametric PET imaging with PET probes for other kinds of monoamines or with PET probes for a more general neurophysiologic index is required. Our preliminary PET study using 18F-BCPP-EF has revealed that MC-I activity outside nigrostriatal regions is impaired in the brain of MPTP-treated monkey (H. Tsukada and M. Kanazawa, unpublished data, March 2012).
CONCLUSION
The preliminary results of the present study suggest that 18F-BCPP-EF has potential as a PET probe for the quantitative imaging of neuronal degeneration as impaired MC-I activity in the living brains of PD model monkeys.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. No potential conflict of interest relevant to this article was reported.
Acknowledgments
We gratefully acknowledge the technical assistance of Dai Fukumoto and Aiko Iwazaki.
Footnotes
Published online Feb. 11, 2016.
- © 2016 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication November 11, 2015.
- Accepted for publication February 3, 2016.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The relationship between brain activation and mitochondrial complex I protein levels during cognitive function in healthy humans: an [18F]BCPP-EF PET and functional MRI study of task switching
- In vivo mitochondrial and glycolytic impairments in patients with Alzheimer disease
- Effect of MPTP on Serotonergic Neuronal Systems and Mitochondrial Complex I Activity in the Living Brain: A PET Study on Conscious Rhesus Monkeys