


Abstract
Acetylcholinesterase and butyrylcholinesterase accumulate with brain β-amyloid (Aβ) plaques in Alzheimer disease (AD). The overall activity of acetylcholinesterase is found to decline in AD, whereas butyrylcholinesterase has been found to either increase or remain the same. Although some cognitively normal older adults also have Aβ plaques within the brain, cholinesterase-associated plaques are generally less abundant in such individuals. Thus, brain imaging of cholinesterase activity associated with Aβ plaques has the potential to distinguish AD from cognitively normal older adults, with or without Aβ accumulation, during life. Current Aβ imaging agents are not able to provide this distinction. To address this unmet need, synthesis and evaluation of a cholinesterase-binding ligand, phenyl 4-123I-iodophenylcarbamate (123I-PIP), is described. Methods: Phenyl 4-iodophenylcarbamate was synthesized and evaluated for binding potency toward acetylcholinesterase and butyrylcholinesterase using enzyme kinetic analysis. This compound was subsequently rapidly radiolabeled with 123I and purified by high-performance liquid chromatography. Autoradiographic analyses were performed with 123I-PIP using postmortem orbitofrontal cortex from cognitively normal and AD human brains. Comparisons were made with an Aβ imaging agent, 2-(4′-dimethylaminophenyl)-6-123I-iodo-imidazo[1,2-a]pyridine (123I-IMPY), in adjacent brain sections. Tissues were also stained for Aβ and cholinesterase activity to visualize Aβ plaque load for comparison with radioligand uptake. Results: Synthesized and purified PIP exhibited binding to cholinesterases. 123I was successfully incorporated into this ligand. 123I-PIP autoradiography with human tissue revealed accumulation of radioactivity only in AD brain tissues in which Aβ plaques had cholinesterase activity. 123I-IMPY accumulated in brain tissues with Aβ plaques from both AD and cognitively normal individuals. Conclusion: Radiolabeled ligands specific for cholinesterases have potential for use in neuroimaging AD plaques during life. The compound herein described, 123I-PIP, can detect cholinesterases associated with Aβ plaques and can distinguish AD brain tissues from those of cognitively normal older adults with Aβ plaques. Imaging cholinesterase activity associated with Aβ plaques in the living brain may contribute to the definitive diagnosis of AD during life.
- Alzheimer's disease
- acetylcholinesterase
- autoradiography
- butyrylcholinesterase
- carbamate
- single photon emission computed tomography
Alzheimer disease (AD) is a neurodegenerative disorder and the most common cause of dementia. The prevalence of AD is predicted to dramatically rise over the next decades with concomitant increases in social and economic burdens (1). Efforts toward development of disease-modifying interventions for AD are hampered by a lack of early definitive diagnosis.
Currently, AD diagnosis is confirmed by postmortem brain visualization of β-amyloid (Aβ) plaques and tau neurofibrillary tangles (2). Brain imaging methods aimed at AD diagnosis have emerged involving visualization of Aβ plaques in the living brain. This approach has seen the development of agents such as 2-(4′-dimethylaminophenyl)-6-123I-iodo-imidazo[1,2-a]pyridine (123I-IMPY) (3), [N-methyl-11C-2-(4′-methylaminophenyl)-6-hydroxybenzothiazole (11C-PIB) (4), and (E)-4-(2-(6-(2-(2-(2-18F-fluoroethoxy)ethoxy)ethoxy)pyridin-3-yl)vinyl)-N-methyl benzenamine (18F-florbetapir) (5), among others, that bind to Aβ plaques in vivo and are visualized using SPECT or PET. However, with agents that target Aβ plaques, a major limitation is that a significant number of cognitively normal adults (10% at 50 y to 44% at 90 y) have such Aβ pathology (6,7). This limits the utility of Aβ plaque imaging agents alone for the premortem definitive diagnosis of AD. Consequently, Aβ imaging must be used in conjunction with clinical history and physical examinations as well as cognitive and neuropsychologic testing for the clinical diagnosis of AD (8). Therefore, the need is still great for biomarker development to aid in the diagnosis of AD.
Changes in the cholinergic system have been documented in AD (9). In particular there is loss of cholinergic neurons in the basal forebrain and other regions, thereby giving rise to many cognitive and behavioral symptoms (10). Two enzymes, acetylcholinesterase (AChE) and butyrylcholinesterase (BChE), hydrolyze acetylcholine and coregulate cholinergic neurotransmission in the brain (11). In the AD brain, AChE is significantly reduced (12); however, activity of BChE may increase (12) or remain the same (13,14). Additionally, these enzymes associate with Aβ plaques and neurofibrillary tangles in AD (14,15). The significance of this association remains unclear but may be related to Aβ plaque formation and maturation (16,17). In addition, cholinesterases may associate predominantly with AD plaques and not those found in cognitively normal brain (16,18,19). Therefore, plaque-associated AChE and BChE may represent specific markers for AD. Imaging of cholinesterases in brain Aβ plaques may aid in the diagnosis of AD.
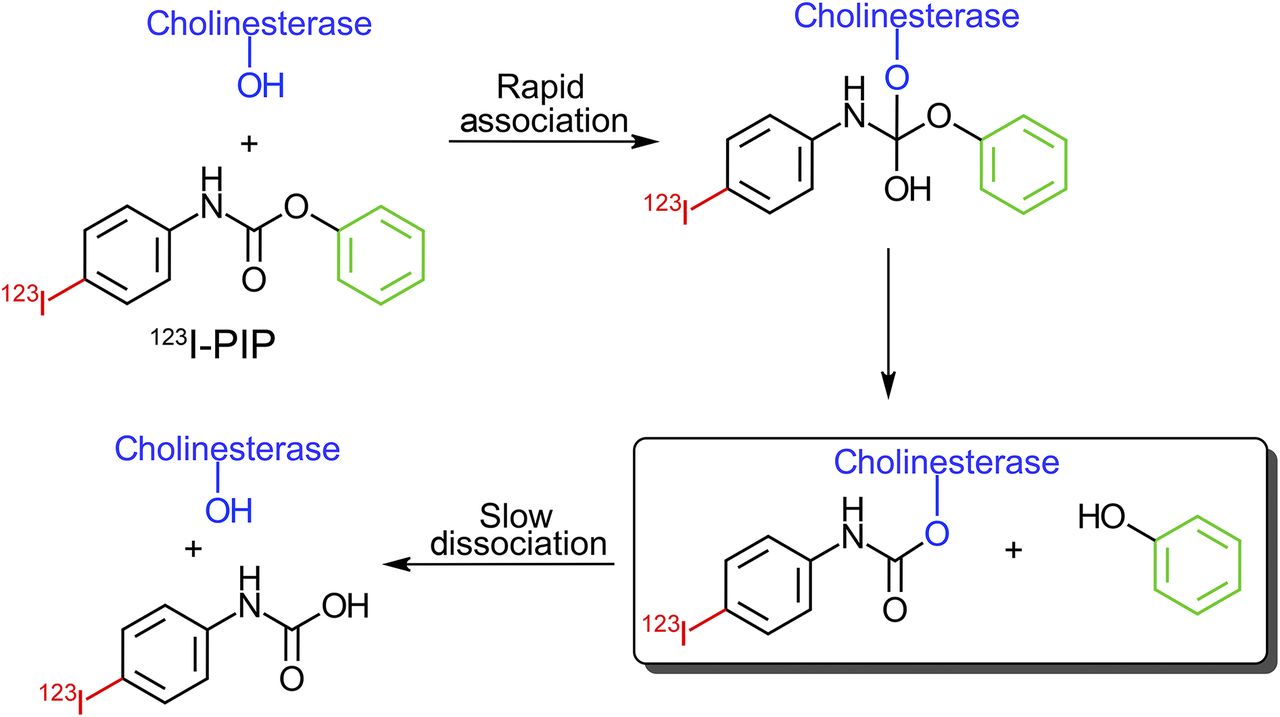
Several agents targeting AChE (20–23) and BChE (24–27) have been tested for human brain imaging of cholinesterase-associated AD pathology. However, focal visualization of cholinesterases associated with AD plaques has not been realized. Furthermore, an increase in radioligand uptake in regions typically associated with cholinesterase AD plaques has not been found in previous studies. For example, in vivo testing of 1-11C-methyl-4-piperidinyl n-butyrate (27), a BChE-specific ester substrate radioligand, demonstrated decreased uptake in AD brain, contrary to the known histochemical and enzymatic isolation studies of this enzyme (14,15). Cholinesterase ester substrates, such as 1-11C-methyl-4-piperidinyl n-butyrate, are rapidly metabolized by these enzymes. Thus, the ability to image cholinesterase activity with such agents relies on the metabolic trapping principle (20,28) in which a radiolabeled hydrophilic product of enzyme catalysis gets trapped within the brain after ester hydrolysis. In contrast, carbamates have dual cholinesterase substrate/inhibitor properties in that they form a covalent bond with the enzymes that is long-lived (Fig. 1). This enzymatic trapping of a radiolabeled imaging agent (29) containing a carbamate functionality is expected to have a longer binding duration to cholinesterases, compared with ester substrates (30,31). With the use of radiolabeled carbamate ligands, the temporal window for visualizing cholinesterase activity associated with AD pathology is potentially increased.

Schematic representation of interaction between cholinesterase catalytic serine and 123I-PIP showing slow dissociation of covalent intermediate containing 123I, a step occurring rapidly with ester substrates.
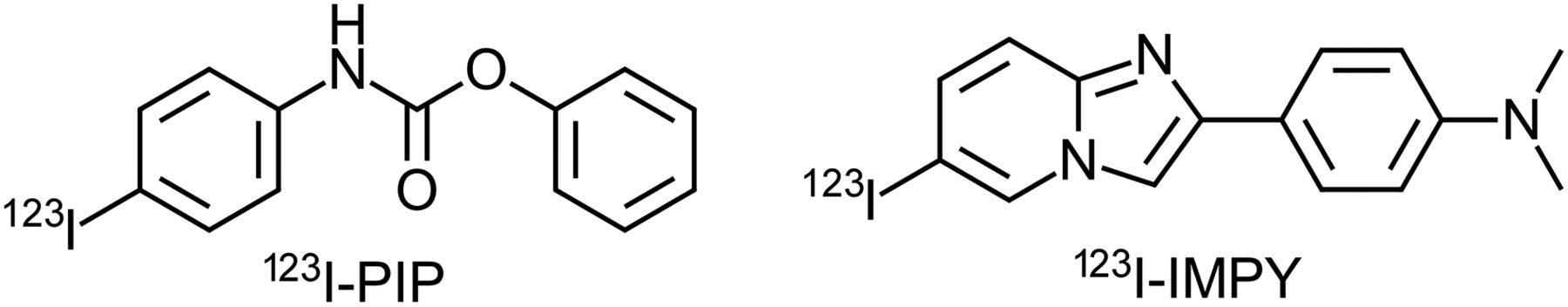
Herein are presented the synthesis, radiolabeling, and preliminary evaluation of phenyl 4-123I-iodophenylcarbamate (123I-PIP, 3), which exhibits affinity toward cholinesterases in kinetic analysis. To demonstrate specificity toward cholinesterases associated with Aβ plaques in AD brains, 3 categories of brain tissues were compared, based on prior histochemical and immunohistochemical analysis, as well as clinical and neuropathologic information: cognitively normal without Aβ plaques (normal/Aβ−), cognitively normal with Aβ plaques (normal/Aβ+), and AD orbitofrontal brain tissues. On the basis of previous observations (18), and confirmed in this preliminary study, cognitively normal individuals have less cholinesterase-associated Aβ plaques than those with AD. It was demonstrated, using autoradiography, that 123I-PIP (3) is able to visualize cholinesterases in Aβ plaques in AD brain tissues. In addition, 123I-PIP (3), in contrast to the Aβ imaging agent 123I-IMPY (Fig. 2), has the ability to distinguish Aβ plaques in AD brain tissues from those in cognitively normal brains.

Structures of cholinesterase imaging agent 123I-PIP and β-amyloid imaging agent 123I-IMPY.
MATERIALS AND METHODS
Synthesis and Radiolabeling
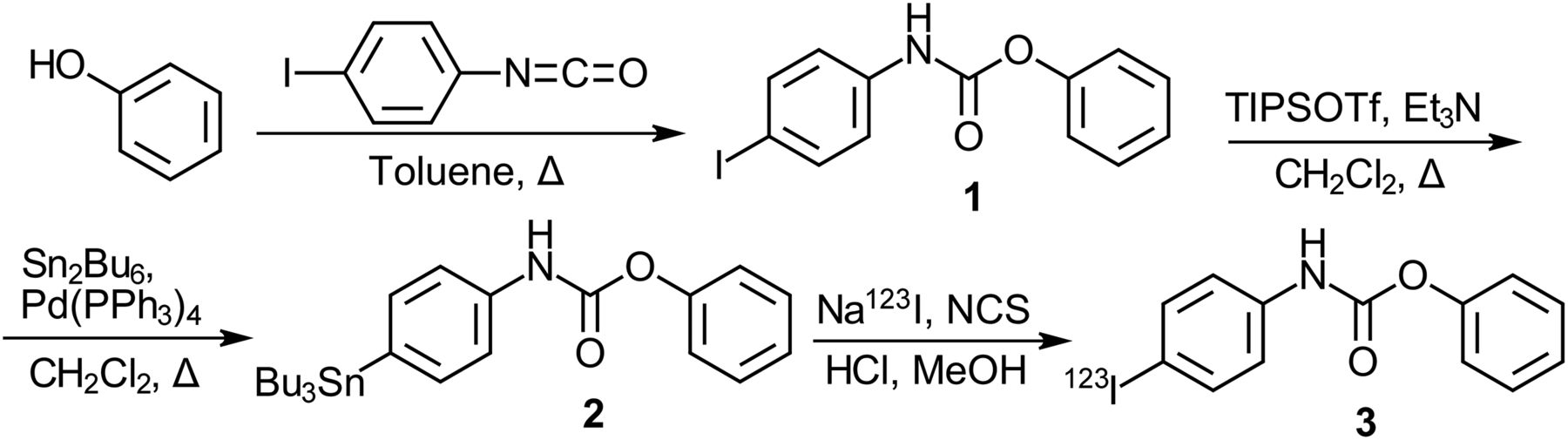
Synthesis of 123I-PIP (Fig. 3)
Phenol (1.021 g, 10.85 mmol) was dissolved in dry toluene (2 mL) under argon atmosphere to which was added iodophenylisocyanate (2.658 g, 10.85 mmol), dissolved in dry toluene (13 mL). This mixture was refluxed for 5 h. The resulting precipitate was hot gravity filtered to collect the resulting white crystalline product (1.983 g, 54%). Phenyl 4-iodophenylcarbamate (1, 0.200 g, 0.59 mmol) was suspended in dry CH2Cl2 (10 mL) under argon atmosphere. To this was added triethylamine (0.18 mL, 1.3 mmol) followed by triisopropylsilyl triflate (0.32 mL, 1.2 mmol). This solution was then added to tetrakis(triphenylphosphine)palladium(0) (0.026 g, 0.023 mmol) to which was added hexabutylditin (0.60 mL, 1.2 mmol). The reaction mixture was refluxed for 16 h, cooled to room temperature, and purified by silica gel flash chromatography (1:9 ethyl acetate/hexanes) to yield the desired compound as a white solid (0.147 g, 50%). To a solution (9 μL) of Na123I (64.42 MBq) in 0.1 M NaOH(aq) (9.0 × 10−7mol) (MDS Nordion) was added 0.1 M HCl (18 μL, 1.8 × 10−6mol) to neutralize the hydroxide. Phenyl 4-tributylstannylphenylcarbamate (2, 50 μL, 4.0 × 10−7mol) was added to the solution followed by N-chlorosuccinimide (28 μL, 8.4 × 10−8mol), both of which were dissolved in MeOH. The reaction was allowed to proceed for 15 min at room temperature; then 0.1 M NaHCO3 (27 μL, 2.7 × 10−3mol) was added. The reaction mixture was purified by high-performance liquid chromatography, using a SemiPrep system with a Zorbax Eclipse XDB-C18, 4.6 × 150 mm, 5-μm column (Agilent Technologies) and 1.0 mL/min of 80% MeOH:20% dH2O eluent. Fractions were collected every 20 s for 15 min with a RediFrac fraction collector (Pharmacia). Retention times were determined using the corresponding cold PIP (1) as a nonradioactive standard (3.9 min). Collected fractions that contained pure product were combined and acidified with HCl (0.1 M, 1 mL), and the solvent was removed under a stream of N2 gas with low heat to yield the desired radiolabeled compound as a residue (radiochemical yield, 87%; radiochemical purity, >96%; specific activity, >3,000 GBq/μmol). The residue was dissolved in 0.1 M maleate buffer, pH 7.4, for incubation with brain tissues. The supplemental data (Supplemental Figs. 1–8, available at http://tech.snmjournals.org) provide analytic data for compounds synthesized (Fig. 3).

Synthesis of PIP (1), corresponding tributyltin precursor (2), and 123I-radiolabeled product (3, 123I-PIP). Et3N = triethylamine; NCS = N-chlorosuccinimide; Pd(PPh3)4 = tetrakis(triphenylphosphine)palladium(0); Sn2Bu6 = hexabutylditin; TIPSOTf = triisopropylsilyl trifluoromethanesulfonate.
Synthesis of 123I-IMPY
123I-IMPY was synthesized according to a previously published procedure, and the analytic data were consistent with the structure of the compound as described (3).
Esterase Activity Assay
Cholinesterase activity was determined using a modification (32) of Ellman’s spectrophotometric method (33). To test for interaction of enzyme with PIP (1; Fig. 2), enzyme activity was determined after various time intervals of incubation with 0.033 mM PIP, a concentration that avoided complete deactivation of enzyme before steady-state conditions were attained. Second-order association constants (ka values) were determined as previously described (34).
Tissue Autoradiography
For this initial evaluation of 123I-PIP (3), the human orbitofrontal cortex was studied, supported by corresponding clinical and neuropathologic information for each brain (Supplemental Table 1). Brain tissue and related information were provided by the Maritime Brain Tissue Bank, after approval from the Nova Scotia Health Research Ethics Board. Cases were designated as AD (n = 10, 83 ± 12 y old), normal/Aβ+ (n = 2; age, 63 and 86 y), and normal/Aβ− (n = 3; age, 55, 63, and 71 y) based on neuropathologic diagnosis and the presence or absence of Aβ plaques. For analysis of 123I-PIP, AD, normal/Aβ+, and normal/Aβ− sections (n = 74, 11, and 30, respectively) were radiolabeled. Brains were immersion-fixed in 10% formalin in 0.1 M phosphate buffer (PB) (pH 7.4) and cut in 1- to 2-cm coronal slabs. These slabs were cryoprotected by immersion in increasing concentrations of sucrose, ranging from 10% to 40% in 0.1 M PB (pH 7.4) and stored in 40% sucrose in 0.1 M PB (pH 7.4) with 0.6% sodium azide until used. Blocks of tissue from the orbitofrontal cortex were cut on a Leica SM2000R microtome with Physitemp freezing stage and BFS-30TC controller in 50-μm serial sections. Sections were stored in 40% sucrose in 0.1 M PB (pH 7.4) at −20°C until used. Before use, sections were thawed and rinsed in 0.1 M maleate buffer (pH 7.4). To each section in 900 μL of 0.1 M maleate buffer pH 7.4 was added 100 μL of maleate buffer (pH 7.4) containing 123I-PIP (3, 1.1 MBq) and the tissue incubated for up to 3 h with gentle agitation. The tissue was then rinsed twice for 1 min each in distilled water, mounted, dried on a slide warmer, and exposed to a high-resolution phosphor imaging screen (GE Healthcare). The screen was scanned with a Typhoon 9400 imager (GE Healthcare) to produce the autoradiogram. Images were saved using ImageQuant (GE Healthcare) and contrast-adjusted so the background from different images matched with Adobe Photoshop CS5.
Autoradiography with 123I-IMPY was performed using a modification of a published procedure (3). Sections to be incubated with 123I-IMPY were thawed, washed in maleate buffer (pH 7.4), mounted, and dried. The sections were rehydrated in maleate buffer (pH 7.4) for 3 min and incubated in 35 mL of maleate buffer (pH 7.4), with 560 μL of 0.1% bovine serum albumin in maleate buffer (pH 7.4) containing 123I-IMPY (1.7 MBq/per section). The tissue was incubated for 1 h with gentle agitation and subsequently rinsed for 2 min in saturated Li2CO3 in 50% EtOH(aq), 2 min in 50% EtOH(aq), and 30 s in distilled water. The sections were dried and exposed to the phosphor imaging screen and images generated as described above.
RESULTS
Synthesis and Radiolabeling
Phenyl 4-iodophenylcarbamate (PIP, 1) was synthesized from phenol and 4-iodophenylisocyanate (Fig. 3). Radiolabeling was performed in 2 steps; the initial step involved protection of the carbamate functionality with a triisopropylsilyl group followed by generation of a tributyltin intermediate, deprotection, and subsequent introduction of 123I.
Synthesis of IMPY followed a previously published procedure (3). The corresponding tributyl tin intermediate was generated and 123I introduced.
123I-PIP (3) and 123I-IMPY were both purified by high-performance liquid chromatography and recovered in high yield, 87% and 71%, and purity, >96% and >99%, respectively. Radiolabeling of 123I-PIP (3) and 123I-IMPY could be performed efficiently and quickly within 1 h.
Cholinesterase Kinetics
PIP (1; Fig. 3) interacted with both AChE and BChE at a concentration (0.033 mM) that avoided complete deactivation of the enzymes. PIP (1) demonstrated time-dependent association with both AChE and BChE, quantified as the second-order association constants (ka). Interaction of AChE with PIP (1; Supplemental Fig. 1) was more rapid (ka: 1.56 ± 0.18 × 104 M−1min−1) than BChE (ka: 3.47 ± 0.82 × 102 M−1min−1).
Human Tissue Autoradiography
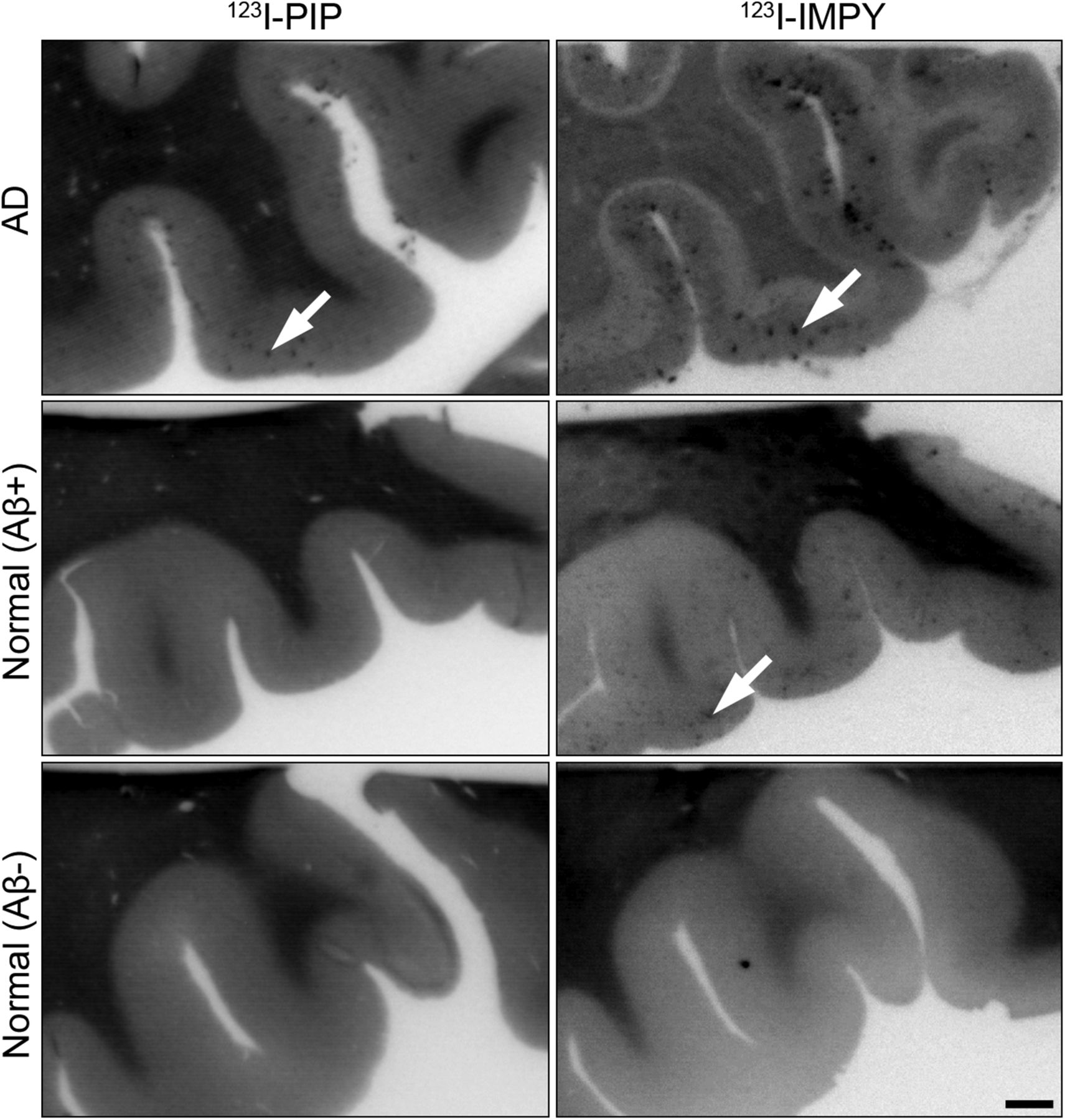
In an overview analysis (Fig. 4) and in all AD cases (74 sections total) radioactivity was observed to accumulate in the orbitofrontal cortex when tissue was exposed to 123I-PIP (3) or 123I-IMPY (Fig. 4, top). Brain tissue from normal/Aβ+ cases revealed radiolabel deposition from 123I-IMPY but no comparable accumulation from 123I-PIP (3) in 11 sections of the orbitofrontal cortex of these brains studied (Fig. 4, middle). No radiolabel accumulation from either agent occurred in the orbitofrontal cortex of 30 sections from normal/Aβ− cases (Fig. 4, bottom).
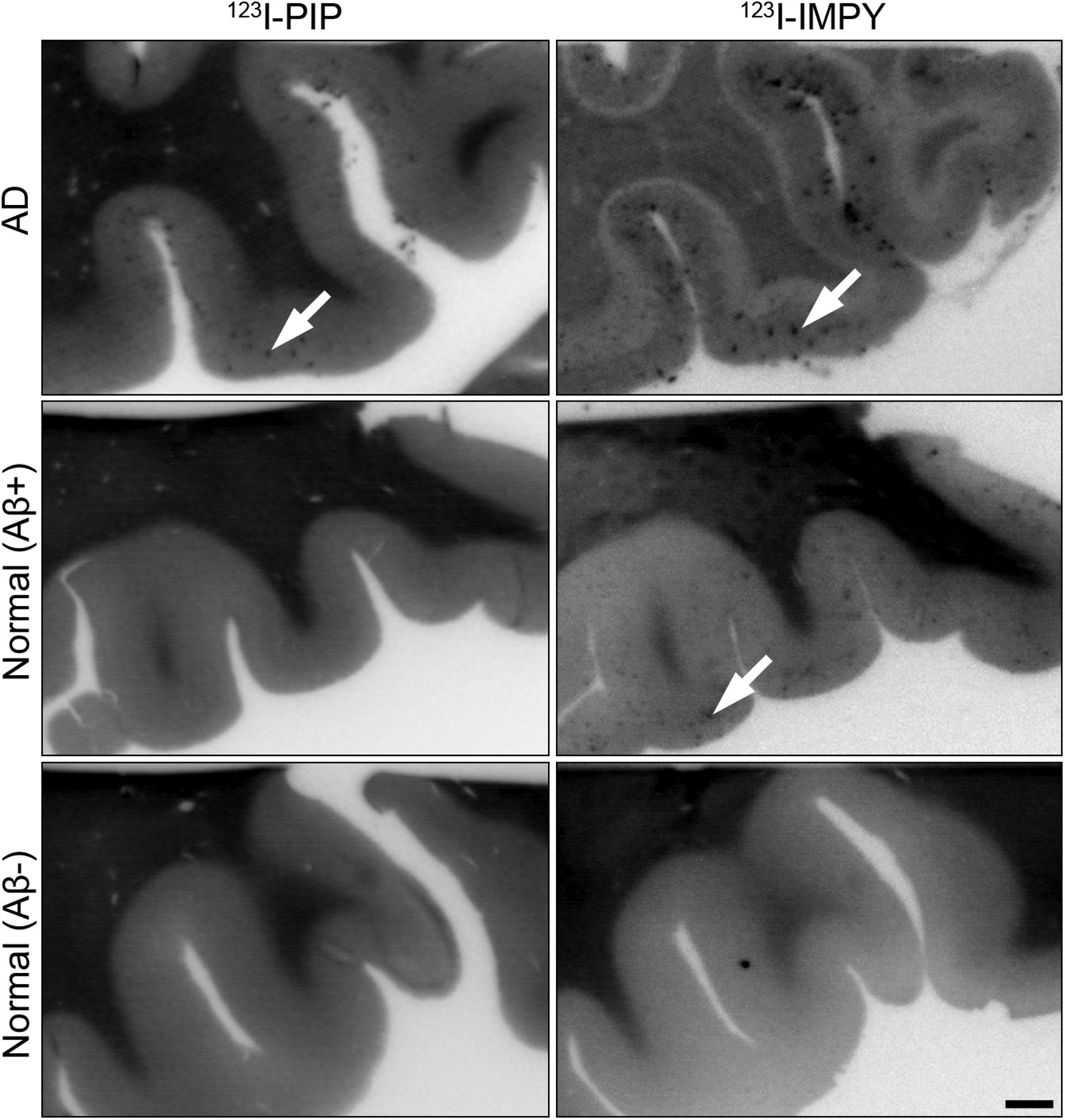
Photomicrographs of representative human orbitofrontal cortex sections from normal/Aβ−, normal/Aβ+, and AD human orbitofrontal cortex showing in vitro autoradiography with 123I-PIP (3) and 123I-IMPY. In all AD brain tissue, sections labeled with 123I-IMPY and 123I-PIP (n = 74) visualized Aβ plaques. Aβ plaques in normal/Aβ+ sections were visualized with 123I-IMPY but were not labeled with 123I-PIP (n = 11). Sections of cortex from normal/Aβ− did not show accumulation of either 123I-PIP (n = 30) or 123I-IMPY. 123I-PIP (3) demonstrates selectivity for detection of Aβ plaques in AD brain and not those in cognitively normal brain. Arrows indicate Aβ plaques. Scale bar = 1 mm.
A more detailed analysis of Aβ plaque visualization compared autoradiography with histochemical techniques for detecting fibrillar Aβ plaques using thioflavin-S and cholinesterase activity in the orbitofrontal cortex region of the brain (Fig. 5). In AD brain tissue, distinct Aβ plaques were visualized with all methods used for analysis. Normal/Aβ+ brain tissue provided Aβ plaque visualization only with thioflavin-S and 123I-IMPY. With normal/Aβ− brain tissue, no Aβ plaques were detected by any of the methods used. Thus, visualization of cholinesterase activity in association with Aβ plaques, whether by histochemical or radiochemical means, is able to distinguish AD pathology from Aβ plaques in normal brain tissue.
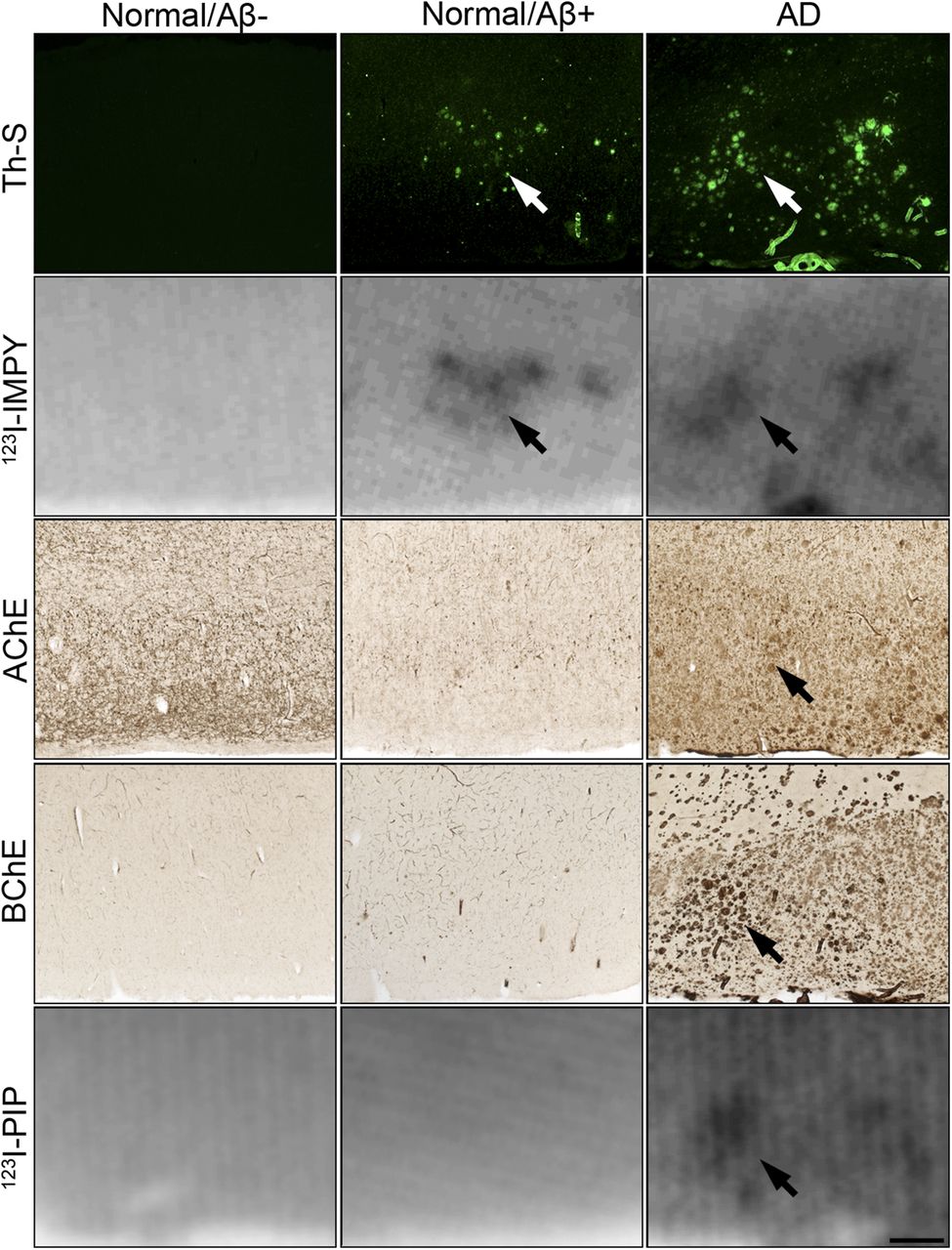
Photomicrographs of representative human orbitofrontal cortex sections from normal/Aβ−; normal/Aβ+; and AD cases showing histofluorescent staining with thioflavin-S (Th-S), autoradiography with 123I-IMPY, histochemical staining for AChE and BChE, and autoradiography with 123I-PIP (3). Accumulation of 123I-IMPY in cortex corresponded to Th-S–positive Aβ deposits in brain tissues from both AD and normal/Aβ+ individuals. Brain tissues from normal/Aβ− individuals did not show cortical accumulation of 123I-IMPY. Autoradiography of Aβ deposition with 123I-IMPY is not able to distinguish AD from normal/Aβ+. Accumulation of 123I-PIP (3) in sections of orbitofrontal cortex (n = 74) corresponded to BChE, AChE, and Th-S–positive Aβ deposits in AD only. Normal/Aβ+ tissues were positive for Th-S; however, BChE and AChE activity was not associated with these structures and 123I-PIP (3) also did not accumulate in these sections (n = 11). Similarly, normal/Aβ− tissues were devoid of plaque-associated cholinesterase activity, and 123I-PIP (3) did not accumulate in these sections (n = 30). Autoradiography of cholinesterase-associated Aβ deposition with 123I-PIP (3) is able to distinguish AD from normal/Aβ+. Arrows indicate plaques visualized using described methods. Scale bar = 500 μm.
DISCUSSION
PIP synthesis (1; Fig. 3) was achieved in a 1-step reaction. 123I-PIP radiolabeling (3; Fig. 3) involved protection of PIP (1) with a triisopropylsilyl group and replacement of iodine by a tributyltin by reaction with hexabutylditin in the presence of palladium catalyst. In contrast to a comparable ester-tributyltin intermediate synthesis (35), attempts at conversion of PIP (1) to the tributyltin intermediate without silyl protection were unsuccessful, suggesting the unprotected carbamate functionality interferes with stannylation of the iodophenyl moiety of PIP (1). The silyl carbamate protecting group was readily removed during the workup. Triisopropylsilyl protection has been reported to protect the nitrogen of carbamates, normally requiring concentrated acid for removal (36). Facile removal of the silyl protecting group from PIP (1) may be due to steric effects of neighboring phenyl rings. The stannylated product was purified from reagents and starting material by high-performance liquid chromatography. After stannylation, incorporation of 123I occurred readily at room temperature in the presence of N-chlorosuccinimide, and the product was purified by high-performance liquid chromatography.
Conversion of IMPY to the tributyltin intermediate was achieved using a procedure similar to that used for PIP (1) and previous ester molecules (35). However, subsequent attempts to incorporate 123I into IMPY using N-chlorosuccinimide as a free radical initiator were unsuccessful. This may be due to the lone pair of electrons on nitrogen atoms in IMPY that are not involved in active π-bonding, thus are available to interfere with free radical generation. The use of hydrogen peroxide to initiate the reaction, as previously described for 123I-IMPY synthesis (3), yielded the desired radiolabeled product.
PIP (1) interacted with both AChE and BChE and, similar to most other carbamates (34,37), produced a time-dependent association with these enzymes, suggesting the formation of a covalently bonded carbamoylated enzyme intermediate. Carbamates are known to form covalent intermediates with the catalytic group of serine hydrolases (30,31). Several factors can influence selectivity for one cholinesterase over another, including the size of the carbamate-containing molecule relative to each enzyme’s active site gorge (30) and interactions with functional groups within the gorge (37). Most previously tested cholinesterase imaging agents are esters and hydrolyzed rapidly by AChE and BChE. Such rapid hydrolysis may be detrimental for in vivo imaging of this enzyme activity by permitting diffusion of the product containing the radiolabel away from the target enzyme. In contrast, time-dependent association of cholinesterases with PIP (1) indicates that radioactivity is bound with enzyme significantly longer than with ester substrates. This longer-lived association of PIP (1) components with cholinesterases is expected to afford an extended window for visualization of this enzyme activity.
Kuhl et al. reported decreased AChE and BChE activity in the brain of AD patients (27). However, this result may reflect synaptic cholinesterase levels as opposed to the plaque-associated cholinesterases thought to contribute to the in vitro and isolation measurements of these enzymes (14,15). Other potential imaging agents evaluated for cholinesterases have also had difficulty in reflecting histochemical and isolation enzymatic levels, possibly due to complex kinetics and low specificity (38). The present study specifically aimed to visualize plaque-associated cholinesterases in AD brain tissue, a result that has not been realized with previously tested cholinesterase imaging agents.
This study used postmortem brain tissue from 3 clinical and neuropathologic classifications, AD, normal/Aβ+, and normal/Aβ− (Fig. 4). Autoradiography of these tissues demonstrated that 123I-IMPY detects, without discrimination, thioflavin-S–positive deposits of Aβ in both AD and normal/Aβ+ brains (Figs. 4 and 5). In contrast, 123I-PIP (3) accumulated only in regions of thioflavin-S–positive Aβ plaques in AD and not normal/Aβ+ brain tissues (Figs. 4 and 5). Furthermore, these focal 123I-PIP (3) accumulations correspond to areas of histochemically detected cholinesterase activity associated with Aβ plaque pathology (Fig. 5). Although the sample of brains used in this preliminary study is small, the observations are consistent with other published data showing that BChE can distinguish plaques of aging from those of dementia (18). Several additional studies are required to further validate plaque-associated cholinesterase imaging for the diagnosis of AD. Because of the limited number of brain tissues analyzed in the current study, a larger postmortem cohort with examination of various brain regions will be required to further assess the capability of such cholinesterase carbamate imaging agents to distinguish between cognitively normal and AD brains. In addition, cholinesterases are found in high concentrations in the plasma, thus, potentially presenting a challenge for the intravenous administration of such cholinesterase imaging agents to reach the brain for in vivo assessment. However, in this respect, studies of BChE imaging agents to date (27,35) suggest that cholinesterase radiopharmaceuticals can be delivered to the brain in sufficient quantity for imaging enzyme activity. SPECT imaging of transgenic AD mice may further elucidate the in vivo characteristics of this cholinesterase imaging agent.
In the present study, 123I-PIP (3) recapitulated the histochemical association of cholinesterases with Aβ plaques and, in a small postmortem sample, distinguished AD brain pathology from cognitively normal brains containing Aβ plaques, based on the presence or absence of cholinesterase activity.
CONCLUSION
The cholinesterases AChE and BChE are associated with Aβ plaques in the AD brain and are potential imaging targets for the diagnosis of this disease. We have described the synthesis and radiolabeling of the cholinesterase ligand 123I-PIP, which binds to both enzymes. This radioimaging agent visualized cholinesterase activity associated with Aβ plaques in human AD brain tissues. In contrast, this imaging agent does not detect Aβ plaques in cognitively normal brain tissue in which associated cholinesterase activity is minimal. Thus, in this preliminary study 123I-PIP was able to differentiate AD brain from normal/Aβ+. Imaging cholinesterase-associated Aβ pathology is a significant advance for this biomarker and is a promising approach toward the definitive diagnosis of AD during life.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This research was supported by the Canadian Institutes of Health Research (MOP-82798, RNS-117795, MOP-119343), Capital Health Research Fund, Nova Scotia Health Research Foundation (MED-MAT-2011-7512), Faculty and Department of Medicine of Dalhousie University, Natural Sciences and Engineering Research Council of Canada, Killam Trusts, Innovacorp, Dalhousie Medical Research Foundation, Gunn Family Research Prize, the Dalhousie Medical Research Foundation Irene MacDonald Sobey Endowed Chair in Curative Approaches to Alzheimer's Disease, and Ms. Sadie MacLeod. Sultan Darvesh is a scientific cofounder and stockholder in Treventis Corporation, a biotech company focused on development of diagnostic and therapeutic agents for Alzheimer's disease. Sultan Darvesh, Ian R. Macdonald, Ian R. Pottie, and Earl Martin are listed as inventors on patents related to this work, and these patents are assigned to Treventis Corporation. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Sean Jay Kelly for his technical assistance.
Footnotes
Published online Nov. 5, 2015.
- © 2016 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication June 11, 2015.
- Accepted for publication October 21, 2015.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.