


Abstract
γ-aminobutyric acid (GABA)-A receptor–mediated neural transmission is important to promote practice-dependent plasticity after brain injury. This study investigated alterations in GABA-A receptor binding and functional and anatomic connectivity within the motor cortex in children with cerebral palsy (CP). Methods: We conducted 18F-fluoroflumazenil PET on children with hemiplegic CP to investigate whether in vivo GABA-A receptor binding is altered in the ipsilateral or contralateral hemisphere of the lesion site. To evaluate changes in the GABA-A receptor subunit after prenatal brain injury, we performed GABA-A receptor immunohistochemistry using rat pups with a diffuse hypoxic ischemic insult. We also performed diffusion tensor MR imaging and resting-state functional MR imaging on the same children with hemiplegic CP to investigate alterations in anatomic and functional connectivity at the motor cortex with increased GABA-A receptor binding. Results: In children with hemiplegic CP, the 18F-fluoroflumazenil binding potential was increased within the ipsilateral motor cortex. GABA-A receptors with the α1 subunit were highly expressed exclusively within cortical layers III, IV, and VI of the motor cortex in rat pups. The motor cortex with increased GABA-A receptor binding in children with hemiplegic CP had reduced thalamocortical and corticocortical connectivity, which might be linked to increased GABA-A receptor distribution in cortical layers in rats. Conclusion: Increased expression of the GABA-A receptor α1 subunit within the ipsilateral motor cortex may be an important adaptive mechanism after prenatal brain injury in children with CP but may be associated with improper functional connectivity after birth and have adverse effects on the development of motor plasticity.
Cerebral palsy (CP) refers to a broad range of diseases with nonprogressive permanent motor impairment that originate from insult to the immature brain. The overall prevalence of CP ranges from 1.5 to 2.5 per 1,000 children, and the number of children with CP has not fundamentally changed despite recent advances in prevention and therapeutic approaches (1). Among the various subtypes of CP, the spastic type is most common, occurring in 80% of cases (2). The cause of CP is multifactorial, including prenatal hypoxic ischemic injury, asphyxia, infections or inflammation, coagulopathies, and inheritance (3,4). Regardless of underlying etiologic factors, poor motor performance is a common manifestation in patients with CP.
During the developmental period, the brain has a great ability to compensate for neuronal injury (5,6). In particular, neural plasticity in early development allows recovery of motor performance even after hemispherectomy (7). Animal studies also show few deficits in motor function after neonatal cortical injury (8,9).
Despite this great capacity for plasticity in early development, motor disability in patients with spastic CP does not significantly improve even after constraint-induced movement therapy. Patients with congenital hemiparesis, even those who have a localized infarction in the white matter region, fail to develop sufficient motor functional recovery. Therefore, there might be unknown factors that hinder development of compensatory plasticity in CP.
γ-aminobutyric acid (GABA), an inhibitory neurotransmitter, acts as a neurotropic factor, and GABA receptor–mediated neural transmission plays an important role in cortical development (10). GABAergic neurons are most vulnerable to insult at 25 wk of gestation, a common period of premature brain injury (11). Therefore, we hypothesized that alterations in the expression of the synaptic or extrasynaptic GABA-A receptors could be a potential regulating factor in poor motor performance in patients with CP, as neuronal excitatory-inhibitory balance is important for proper motor performance.
Previously, Lee et al. (12,13), using 18F-fluoroflumazenil PET, demonstrated increased regional GABA-A receptor binding potential (BP) bilaterally in the motor areas of children with spastic diplegic CP who have bilateral periventricular leukomalacia. 18F-fluoroflumazenil was recently introduced for central benzodiazepine receptor imaging (14,15). However, it remains to be elucidated whether GABA-A receptor binding is increased within the ipsilateral motor cortex of the affected hemisphere or contralateral hemisphere after perinatal brain injury. It also is not yet clear which GABA-A receptor subunit among the diverse receptor subtypes is altered. Also unknown is whether the 18F-fluoroflumazenil increase in the motor area is specific to spastic diplegic CP, mostly occurring because of periventricular leukomalacia secondary to hypoxic ischemic insult, or is a common cortical adaptive process after various types of brain injury occurring during a critical period.
Therefore, we investigated 18F-fluoroflumazenil BPs in children with hemiplegic CP who had unilateral perinatal ischemic stroke without obvious overlying cortical injury, and we performed immunohistochemistry using the brain tissues of a rat pup model with diffuse hypoxic ischemic injury to test whether the alterations in GABA-A receptor binding observed in our human imaging studies can be replicated in an animal model and to evaluate GABA-A receptor subtypes and their layer-specific distribution within the overlying cerebral cortices.
We further evaluated GABA-A receptor binding with regard to brain connectivity by means of diffusion tensor imaging (16) for anatomic connectivity and task-independent resting-state functional MR imaging for brain functional connectivity (17).
MATERIALS AND METHODS
Human Subjects
Six children with congenital hemiplegic motor impairment (spastic hemiplegic CP, 2 male and 4 female patients; age range, 7–20 y; mean ± SD, 12.5 ± 4.8 y) without a history of epilepsy were included. The underlying etiology of congenital hemiplegia was proven to be perinatal ischemic stroke on conventional MR imaging, including venous infarction or small arterial infarction involved exclusively within the white mater as described previously (18). Patients with gray matter structural abnormalities such as cortical infarction, malformation, or any other type of congenital anomaly that may have influenced the cortical biodistribution of GABA-A receptors were excluded on the basis of visual assessment of conventional MR imaging (Fig. 1). Patient demographic data, including the Gross Motor Function Classification System, are in Table 1.

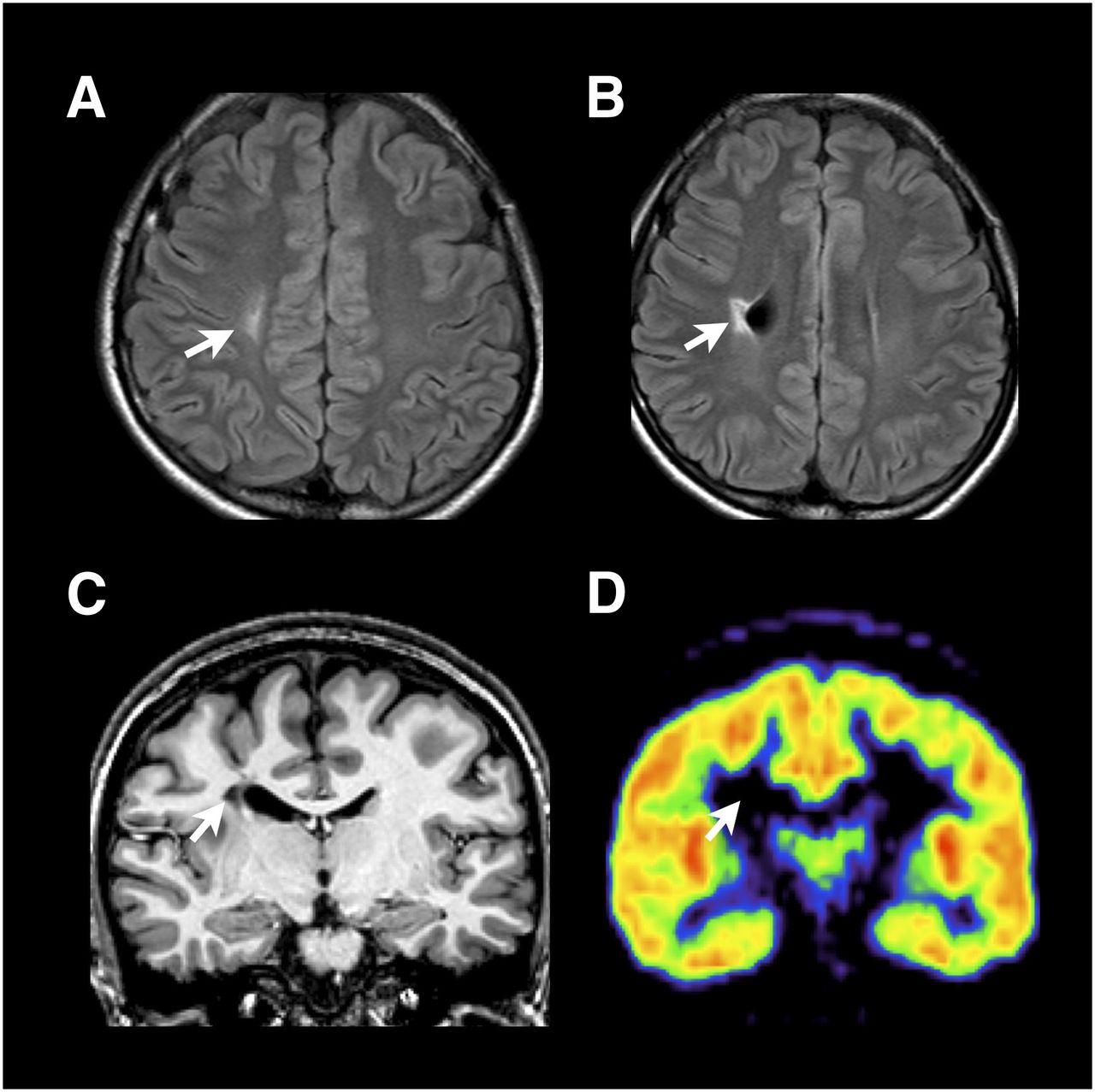
A 20-y-old woman with left hemiplegia. (A and B) Fluid-attenuated inversion recovery MR axial images show focal ischemic infarction within right centrum semiovale with focal dilatation of lateral ventricle. (C) T1-weighted coronal image shows periventricular focal infarction extending to corticospinal tract without overlying gray matter injury. These findings are compatible with focal venous infarction. (D) Fluoroflumazenil binding-potential map corresponding to slice in C shows increased BP at affected-side motor cortices. Arrows indicate lesion site.
Demographic Data of Patients with Hemiplegic CP
Data from 18 healthy young subjects (9 male and 9 female) who participated in the previous 18F-fluoroflumazenil PET study were used as a control group (13). Their ages ranged from 19 to 24 y, with a mean of 21.3 ± 1.3 y. For structural MR imaging, diffusion tensor imaging, and functional MR imaging, data from 18 age- and sex-matched healthy control subjects (8 male and 10 female; age range, 7–20 y; mean, 13.5 ± 4.7 y) from a database (13) were used. The Institutional Review Board approved this study, and all patients or their legal representatives signed a written informed consent form.
18F-Fluoroflumazenil PET Imaging
The image acquisition and analysis methods for 18F-fluoroflumazenil PET were the same as described in our previous paper (13). In brief, dynamic PET scans were given to 6 patients and 18 control subjects using a DSTe scanner (GE Healthcare) in a sequence of 150 frames. Voxelwise BP was calculated using a multilinear reference tissue method (19) with mean activity within the manually delineated pons as reference tissue activity using PMOD software (PMOD Technologies Ltd.). We assumed that the density of GABA-A receptors in the pons is negligible. When the brain had a left-sided lesion, we flipped brain images to locate the lesion side to the right hemisphere.
Spatial preprocessing and statistical analysis of BP maps were performed using SPM8 software (Institute of Neurology, University College London). A BP map of each individual was transformed to the group template in Montreal Neurologic Institute (MNI) space using a nonlinear transform function of the individual T1-weighted image in BP space to the template. These maps were convolved with an isotropic gaussian kernel of 8 mm in full width at half maximum. To evaluate regional alterations in BP, we remove the effects of global differences in BP by scaling the BP of each voxel with the mean BP from the entire brain. After voxelwise application of a 2-sample t test between groups, clusters composed of a minimum of 65 contiguous voxels with a threshold of P < 0.001 (uncorrected) were considered significant, corresponding to P < 0.05 corrected for multiple comparisons estimated by 10,000× Monte Carlo simulations (20).
We also compared the 18F-fluoroflumazenil BP in the affected and intact sides to compensate for age effects in the group comparison study (supplemental data, available online at http://jnm.snmjournals.org).
Animal Hypoxic Model
Five rat pups (7 d old, equivalent to 23–36 wk of gestation in humans (21)) were subjected to 2 h of hypoxia in a humidified hypoxia chamber with ambient O2 levels continuously regulated at around 5%. We performed these experiments in accordance with the requirements of the Institutional Animal Care and Use Committee of Yonsei University College of Medicine. The chamber was flushed with 95% N2 and 5% CO2 for 2 h to keep the rats under hypoxic conditions. Hypoxia was chosen as an insult because prenatal hypoxia is implicated in most cases of CP. Control rats were kept under normoxic conditions. The ambient temperature of the chamber was controlled at 37.5°C by water circulation from a temperature-controlled bath. This hypoxic ischemia method was chosen as an animal model of CP because this method preserves the cortices relatively well without severe cerebral ischemia, which is generally entailed in the model of middle cerebral artery occlusion. Among fluoroflumazenil binding sites, α1, α2, and α3 subunits are critical during development; we examined expression patterns of these 3 subunits using immunohistochemistry.
GABA-A Receptor Immunohistochemistry in Animal Model
Rat pups that were kept under normoxic conditions (n = 5) or exposed to hypoxia (n = 5) at postnatal day 7 were anesthetized by an intraperitoneal injection of ketamine and xylazine and perfused intracardially with 4% paraformaldehyde in phosphate-buffered saline at 10 d after birth (corresponding to 36–40 wk in humans). The brains were removed and fixed in 4% paraformaldehyde for 24 h at 4°C and then were immersed in 30% sucrose in phosphate-buffered saline until they sank. They then were cryosectioned into 40-μm sagittal sections (1.2–1.4 mm lateral to the midline) in a cryostat. After permeabilization with 0.2% Triton X-100 (The Dow Chemical Co.), sections were incubated with 3% bovine serum albumin in phosphate-buffered saline for 1 h.
Rabbit polyclonal GABA-A receptor α1 subunit antibody (Alomone), α2 subunit antibody (Sigma), and α3 subunit antibody (Alomone) were applied at 1:250 at 4°C overnight followed by horseradish peroxidase–conjugated antirabbit secondary antibody at 1:250 for 2 h. After being washed 3 times, the sections were treated with diaminobenzidine tetrahydrochloride and H2O2. Brain sections were placed on glass slides, mounted using Permount medium (Fisher Scientific), and qualitatively analyzed by an experienced specialist. Densitometric analysis was then performed to confirm the visual evaluation results.
Tract-Based Spatial Statistics of the White Matter Anisotropy in Patients with Hemiplegic CP
All structural MR imaging, diffusion tensor imaging, and resting-state functional MR imaging were performed using a 3.0-T scanner (Intera; Philips Medical Systems) and analyzed using the same method as described in our previous paper (13). In brief, high-resolution T1-weighted MR imaging volumes were acquired with a 0.859 × 0.859 × 1.2 mm voxel size. We performed diffusion tensor imaging using a single-shot echo-planar acquisition from 45 diffusion-encoded gradient directions with 1.72 × 1.72 × 2 mm voxels, an echo time of 60 ms, and a b-factor of 600 s/mm2.
We flipped all MR images to locate the lesion in the right hemisphere when the brain had a left-sided lesion. The fractional anisotropy (FA) values were analyzed using a tract-based spatial statistics technique (22) including eddy current correction, generation of a mean FA skeleton, projection of each subject’s FA map onto the mean FA skeleton, and voxelwise statistical evaluation of FA in the skeleton. To correct for multiple comparisons using the threshold-free cluster enhancement approach in tract-based spatial statistics, regional FA differences were given a threshold at the level of threshold-free cluster enhancement-corrected P < 0.05. We also evaluated the hemispheric difference in FA in patients by comparing initial FA data used for the group comparison and flipped data using the method described above.
Statistical Analysis of Resting-State Motor Functional Connectivity in Patients with Hemiplegic CP
We acquired 165 functional MR imaging scans from each subject using an echo-planar sequence with a voxel size of 2.75 × 2.75 × 4.8 mm, a repetition time of 2,000 ms, and an echo time of 30 ms. During functional MR imaging, the subjects were instructed to stay awake with their eyes closed, without focusing on a specific thought and without moving. After removal of the first 5 scans, the remaining 160 scans underwent acquisition time delay correction and head motion correction, followed by spatial normalization by application of nonlinear transformation from the coregistered T1-weighted image in the functional MR imaging space to the MNI template using SPM8. Motion effects were further corrected using independent component analysis by reducing components highly correlated (>0.7) with 1 of 6 motion parameters. Finally, normalized data were spatially smoothed using a gaussian filter of 4 mm in full width at half maximum.
To evaluate functional connectivity, correlation coefficients between the time series of the 18F-fluoroflumazenil–increased region in the primary motor cortex in CP and those of entire brain voxels were derived after band-pass filtering (0.01–0.08 Hz) and regressing out motion effects and global signal changes at white matter, cerebrospinal fluid, and whole brain. To examine group differences, a 2-sample t test was applied to z values of the correlation coefficients after the Fisher r-to-z transformation for each voxel. Clusters with a minimum of 39 contiguous voxels with a threshold of P < 0.005 (uncorrected) were considered significant, equivalent to P < 0.05 corrected by the cluster size threshold as described above.
RESULTS
Statistical parametric maps of intergroup differences in 18F-fluoroflumazenil BP revealed that, in comparison with the control group, the patients with hemispheric CT had increased GABA-A receptor BP in the primary motor cortices (paracentral gyrus and precentral gyrus) and the occipital cortex of the affected hemisphere (Fig. 2; Table 2). When we compared the 18F-fluoroflumazenil BP of the affected hemisphere with that of the intact hemisphere, a similar increase was found in the affected side specifically in the paracentral gyrus, precentral gyrus, anterior cingulate gyrus, and postcentral gyrus (Supplemental Fig. 1).
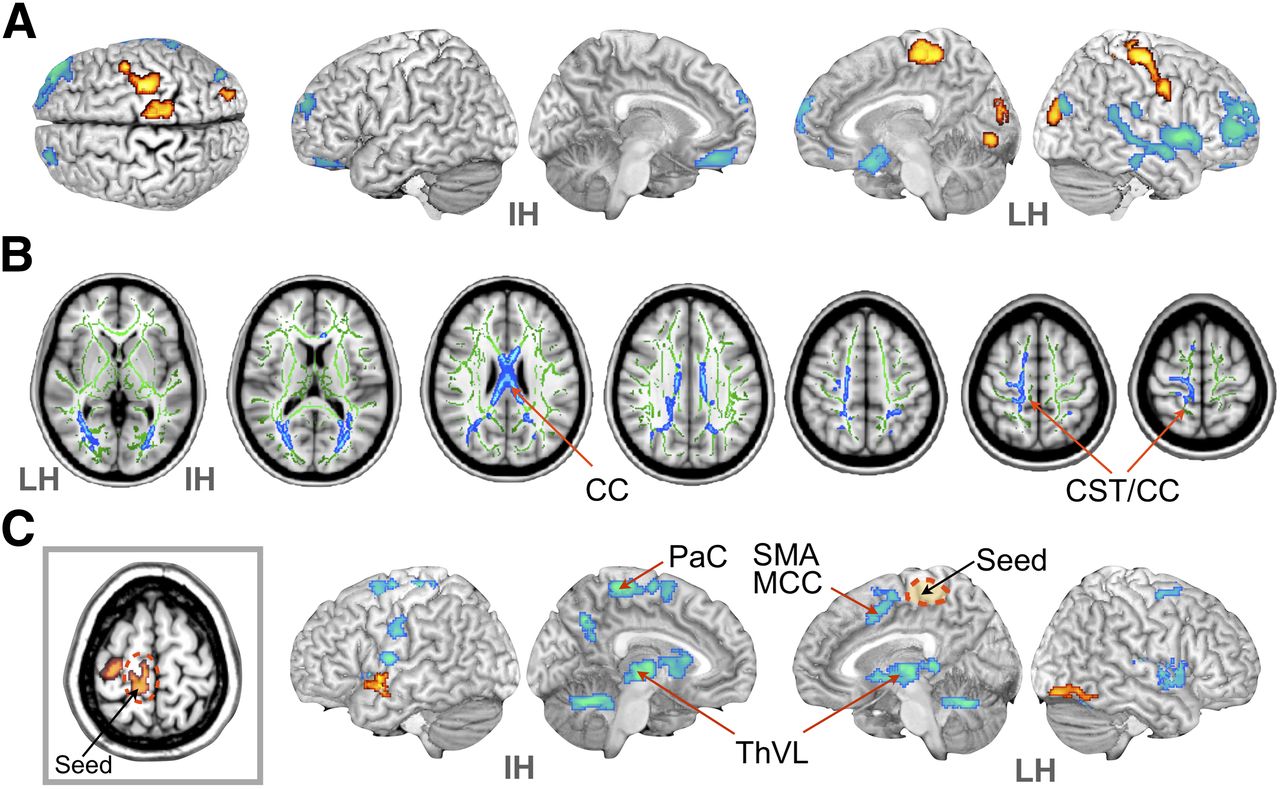
Statistical maps of fluoroflumazenil binding potential, diffusion anisotropy, and functional connectivity in patients with hemiplegic CP. (A) Map of regional differences in fluoroflumazenil binding potential. Red indicates increased binding potential in patients with hemiplegic CP, compared with healthy controls; blue indicates reduced binding potential in hemiplegic CP, compared with controls (P < 0.05, corrected cluster size). (B) Regions of decreased FA in hemiplegic CP, compared with controls (P < 0.005, threshold-free cluster enhancement-corrected [blue] on white matter skeleton [green]). (C) Group difference in functional connectivity between controls and hemiplegic CP from seed region detected using 2-sample t test (P < 0.05, corrected cluster size). A seed region (dashed circle) was chosen in paracentral gyrus of affected hemisphere (rectangle), where hemiplegic CP had increased fluoroflumazenil binding potential. Red indicates increased connectivity from seed region in hemiplegic CP; blue indicates reduced connectivity from seed region, compared with controls. CC = corpus callosum; CST = corticospinal tract; IH = intact hemisphere; LH = lesioned hemisphere; MCC = middle cingulate gyrus; PaC = paracentral lobule; SMA = supplementary motor area; ThVL = ventrolateral thalamus.
Results of Regional 18F-Fluoroflumazenil Binding-Potential Analysis
GABA-A receptor α1 immunoreactivity increased exclusively in motor areas in rats exposed to hypoxic conditions, compared with wild-type normal controls (Fig. 3A, top). Preincubation of GABA-A receptor α1 subunit antibody with an α1 blocking peptide blocked α1 immunoreactivity in cortical slices (Fig. 3A, bottom). α1 immunoreactivity increased selectively in cortical layers III, IV, and VI (Fig. 3B). In contrast, the expression of GABA-A receptor α2 and α3 subunits was not significantly altered in this study (Fig. 3C).
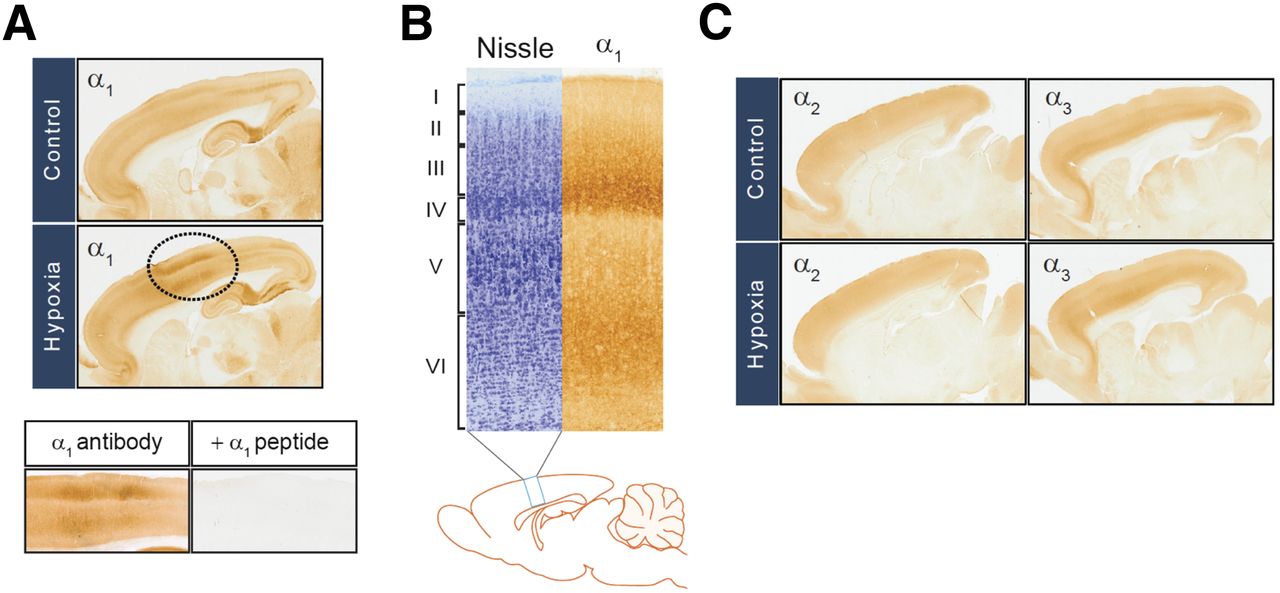
Hypoxia-increased expression of GABA-A receptor α1 subunit in rat motor cortex. (A, top) Immunohistochemistry was performed using GABA-A α1 receptor subunit antibody in sagittal sections of rat brains removed from 10-d-old rat pups preexposed to 5% hypoxia for 2 h at 7 d after birth. (A, bottom) Blocking peptide for α1 receptor subunit antibody was used to show specificity of α1 immunosignals. (B) Motor cortical layers were defined using Nissl stain and GABA-A α1 receptor immunohistochemistry. (C) Immunohistochemical analysis was performed as in A for GABA-A α2 and α3 receptor subunits.
Tract-based spatial statistical analysis showed reduced FA over a wide area, primarily in the affected hemisphere. FA reduction within patients with hemispheric CP, compared with controls, was observed in the white matter around the motor area and corpus callosal fibers (Fig. 2B). Compared with the contralateral hemisphere, the lesion side showed similarly reduced anisotropy in the thalamus, corpus callosum, and corona radiata (Supplemental Fig. 2).
The paracentral gyrus with increased 18F-fluoroflumazenil in the affected hemisphere of patients with CP had reduced functional connectivity with the motor-related regions such as the ipsilateral middle cingulate gyrus, supplementary motor area, right and left thalamus, and contralateral motor areas (Fig. 2C).
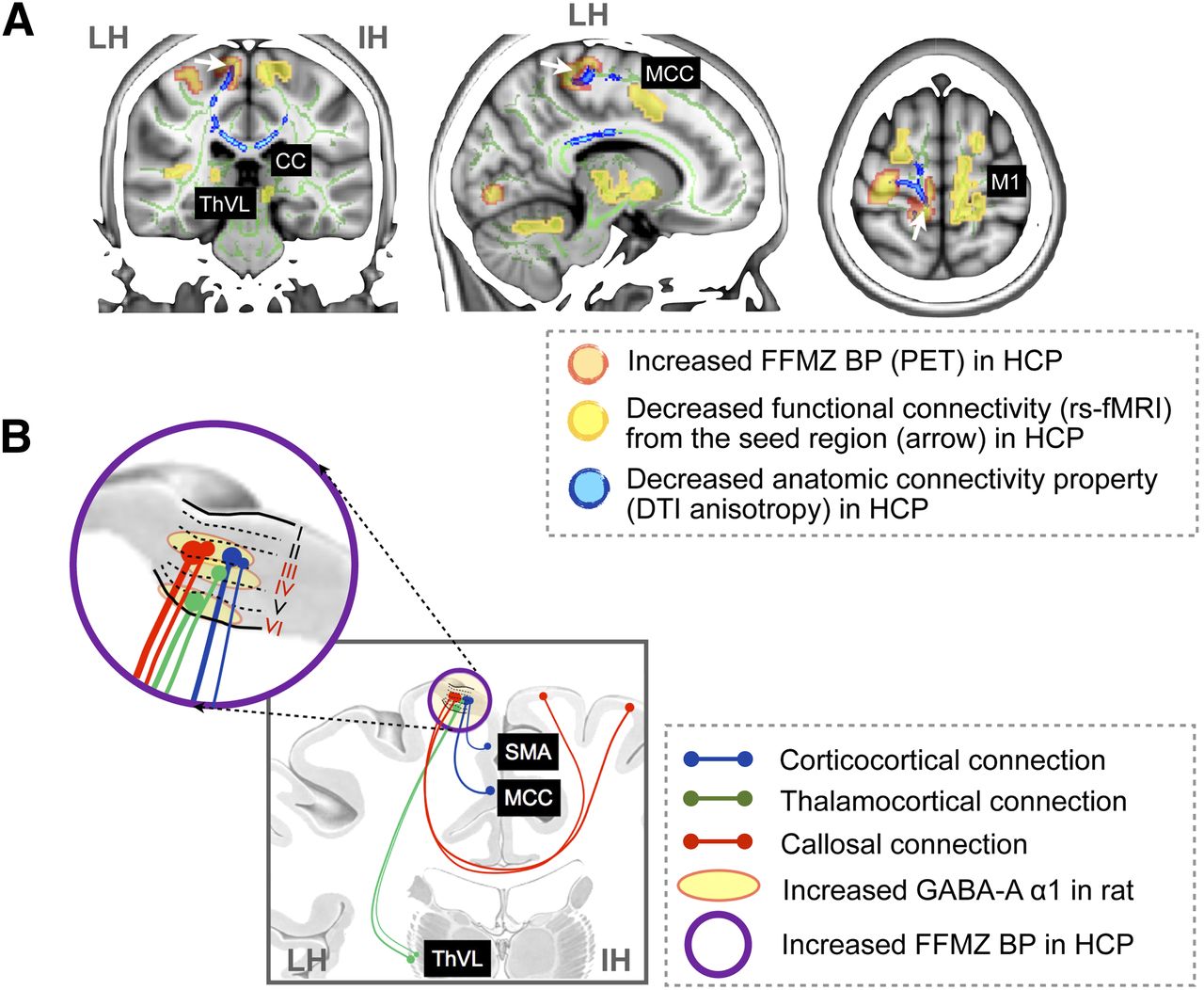
As summarized in Figure 4, the increased 18F-fluoroflumazenil BP regions in the paracentral gyrus of the affected hemisphere had reduced functional connectivity with the bilateral thalamus and motor cortical area. The decreased FA area in patients with hemiplegic CP corresponded to the functional connectivity results (Fig. 4A). This altered connectivity in neuroimaging data was highly associated with neural circuits corresponding to cortical layers III, IV, and VI, where overexpressed GABA-A receptor α1 subunit was observed in rats (Fig. 4B).

Summary of neuroimaging results and animal study results. (A) Fluoroflumazenil BP-increased regions (red in Fig. 3A), diffusion anisotropy-decreased regions (blue in Fig. 3B), and functional connectivity-increased regions (yellow in Fig. 3C) in patients with hemiplegic CP were overlaid on MR coronal, sagittal, and axial sections. Both functional connectivity from increased fluoroflumazenil BP region defined using resting-state functional MR imaging and corresponding anatomic connectivity property defined by FA of diffusion tensor imaging were reduced in hemiplegic CP. (B) Summary illustration shows motor cortex with reduced connectivity in patients with hemiplegic CP (Fig. 3) and neural circuits corresponding to cortical layers III, IV, and VI with overexpressed GABA-A α1 in rats (Fig. 2). CC = corpus callosum; DTI = diffusion tensor imaging; FFMZ = fluoroflumazenil; HCP = hemiplegic CP; IH = intact hemisphere; LH = lesioned hemisphere; M1 = primary motor cortex; MCC = middle cingulate gyrus; rs-fMRI = resting-state functional MR imaging; SMA = supplementary motor area; ThVL = ventrolateral thalamus.
DISCUSSION
In this study, patients with hemiplegic CP showed increased 18F-fluoroflumazenil BP within the affected-side motor cortex. Since 18F-fluoroflumazenil PET is not specific for GABA receptor subunit types and does not resolve their distributions within cortical layers, this study using immunohistochemistry of rat pups revealed that the GABA-A receptors were hyperpolarizing the α1 subunit and were highly expressed exclusively within cortical layers III, IV, and VI of the motor cortex in the rat model. The motor cortex of patients with increased GABA-A receptor binding had reduced functional and anatomic connectivity with the thalamus or other cortical regions, as might be linked to increased GABA-A receptor distribution in cortical layers in rats.
In addition to the increased 18F-fluoroflumazenil BP at the motor cortex, increased 18F-fluoroflumazenil BP at the visual cortex and decreased BP at the frontal lobe were similarly found and discussed in our previous studies with diplegic CP (12,13). The frontal lobe decrease of 18F-fluoroflumazenil BP was also evidenced in a postmortem study that showed a significant loss of GABAergic neurons and receptor density at the frontal regions of infants with periventricular leukomalacia (23).
Since the underlying etiologies of hemiplegic spastic CP are different from those of diplegic spastic CP, the consistent 18F-fluoroflumazenil BP finding between patients with hemiplegic and diplegic CP (13) suggests that the 18F-fluoroflumazenil BP increase in the motor cortex may be a common effect of neural processes after various types of brain injury during a critical period. The current study with hemiplegic CP provides further evidence that the 18F-fluoroflumazenil BP increase was in the affected-side motor cortex rather than the contralateral hemisphere. Considering a well-known phenomenon that the intact contralateral hemisphere is often reorganized to compensate symptomatic deficits caused by the lesion, we may not simply associate the ipsilateral 18F-fluoroflumazenil BP alteration with the compensatory plasticity.
The increased GABA-A BP at the motor cortex in the older children with CP (mean age, 12.5 y) was replicated in the 10-d-old hypoxic rat pups (corresponding to 36–40 wk of gestation in humans (21)). This replication in very young brains implies that increased GABA-A receptor binding in the motor cortex may be associated with adaptive processes after brain damage during prenatal development, rather than practice-dependent changes (i.e., compensatory plasticity) during childhood.
The immunohistochemistry of hypoxic rat pups further suggests that the increased 18F-fluoroflumazenil BP in the motor cortices of the human brain may be attributable to hyperpolarizing α1 subunit-type GABA-A receptors, under the plausible assumption of no additional alteration in the GABA-A subunit after prenatal development. The underlying developmental mechanism of the increased GABA-A receptor α1 subunits in the affected-side motor cortical areas of spastic CP is not clear. To explain CP etiology, early damage of the subplate neurons is often supposed. Ablation of the subplate neuron in the visual ocular column inhibited normal developmental changes in GABA-A receptors and increased α2 and α3 subunits without conversion to α1 subunits (24). However, the increased α1 subunits found in this study might not be explained by the traditional subplate neuronal injury theory, which has focused mainly on the development of the sensory cortex via thalamocortical circuits (24). Thus, mechanisms regulating expression of GABA-A receptor α1 subunits in the motor area remain to be elucidated.
This overexpressed pattern of GABA-A receptor α1 subunit due to brain injury during the developmental period contradicts the expression pattern after brain injury in adults, in whom GABA-A receptor expression has to be reduced to facilitate compensatory plasticity (25–27). This contradiction of GABA-A receptor expression in early and adult brain damage is not surprising since the motor cortex of congenital hemiplegia patients has shown increased metabolism (28) whereas decreased glucose metabolism has been shown in adults damaged by stroke (29). It is well known that cortical disinhibition promotes neural plasticity, especially practice-dependent plasticity (30). Ischemic conditions in the mature brain cause transient downregulation of GABA-A receptors in the affected periinfarct viable neurons or contralateral hemisphere because of extracellular liberation of excessive GABA, thereby prompting practice-dependent plasticity in the motor cortex (31). Conversely, long-term increases in GABA-A receptor function in the motor cortex prevent development of proper practice-dependent plasticity (30) or block somatosensory input-driven motor excitability in the adult brain (32). Therefore, the increased GABA-A receptor binding might adversely affect compensatory plasticity in patients with CP by preventing effective reorganizations between motor areas for the production of movement.
In the current study, overexpression of the inhibitory GABA-A receptor α1 subunit was observed mainly in layers III, IV, and VI of the rat motor cortex. Because cortical layer III is the origin and target of the intracortical and commissural connections, whereas layers IV and VI are the main sites of interconnection between the cortex and the thalamus, increased inhibitory neuronal receptors in these layers of the rat brain may be associated with the decrease in corticocortical, thalamocortical, and interhemispheric connections as revealed on our human neuroimaging data (Fig. 4).
Overexpression of GABA-A receptors in rat cortical layer III may be linked to reduced anisotropy in the callosal fibers and fibers linking motor areas in the affected hemisphere of patients with hemiplegic CP since these fibers are anchored mainly in cortical layer III. Abnormally low diffusion anisotropy in the anterior part of the isthmus of the corpus callosum, a major pathway for callosal fibers of the motor cortex (33), may be well associated with decreased functional interhemispheric connectivity between contralateral motor areas of patients in resting-state functional MR imaging (Fig. 4A).
Within the affected hemisphere, motor areas with increased 18F-fluoroflumazenil binding in patients with spastic hemiplegic CP showed reduced functional connectivity with the ipsilateral cingulate motor area and supplementary motor area. Considering that the cingulate motor area (34,35) and supplementary motor area (36) are involved in higher-order control of movements, the inhibitory neurons in layer III of the motor cortex might hinder appropriate input from these areas for motor performance. The reduced functional connectivity between the motor cortex and the right and left ventrolateral thalamus and reduced diffusion anisotropy in the lesion-side thalamic white matter could be linked to the increased GABA-A receptor expression in layers VI and IV, which are main targets for thalamocortical circuits.
Of interest, the regions showing increased GABA-A receptor binding within the overlying gray matter are linked well to the regions of reduced diffusion anisotropy within the white matter, as might be related to the reduced intra- and interhemispheric functional connectivity (Fig. 4A). In fact, restoration of the interhemispheric and intrahemispheric connection after brain injury is important for compensation of motor deficits (37,38). However, children with hemiplegic CP showed permanent reduced connections, which might be one of the important mechanisms of permanent poor motor function in CP patients.
As a limitation of the current study with hemiplegic patients, the fact that data were obtained from only 6 patients according to the selection criteria of morphologic intactness in the cortex could result in relatively low statistical power for group comparisons between controls. Age differences exist between groups for the 18F-fluoroflumazenil analysis since we could not acquire 18F-fluoroflumazenil PET data from age-matched children because of radiation hazards. Considering that the flumazenil volume of distribution declines as a function of age (39), we applied the global intensity normalization of BP to examine regional differences. We also examined hemispheric differences to compensate for age differences. Although hemispheric asymmetry might exist in the young controls, we found increased 18F-fluoroflumazenil BPs specifically in the motor cortex of the affected side of patients with hemiplegic CP.
The interindividual variability of motor symptoms in the patient group was too low (mostly, Gross Motor Function Classification System level 1 as shown in Table 1) to reveal relationships between findings from 18F-fluoroflumazenil BPs and FAs in diffusion tensor imaging or functional connectivity in resting-state functional MR imaging. In the preliminary correlation analyses for the current results (not presented in this paper), we found a tendency toward a negative correlation between 18F-fluoroflumazenil BPs at the paracentral gyrus and FAs at the corpus callosum in the affected hemisphere (r = −0.811; P = 0.05). However, we did not detect any significant correlations between 18F-fluoroflumazenil BPs and thalamocortical functional connectivity at the lesion-side motor cortex. This finding was not surprising since there was no significant thalamocortical functional connectivity at the lesioned hemisphere when we applied 1-sample t testing for this functional connectivity.
Because of the small sample size with relatively uniform clinical symptoms, we did not correlate current findings with symptoms. When a sufficient number of datasets with diverse symptom levels is accumulated, the findings of the current study would be significantly extended by associating neuroimaging data with behavioral and neurophysiologic data such as motor symptoms and GABA-A–mediated intracortical inhibition.
CONCLUSION
A GABA-A receptor binding study using 18F-fluoroflumazenil PET and a connectivity study using MR imaging techniques clearly demonstrated that alterations in the affected motor cortical GABA-A receptor binding could be an important underlying pathophysiologic mechanism of motor dysfunction in CP patients.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This work was supported by National Research Foundation of Korea (NRF) grant 20100020676 funded by the Korean government (MEST) and by a 2007 faculty research grant (6-2007-0109) from Yonsei University College of Medicine. No other potential conflict of interest relevant to this article was reported.
Footnotes
Published online Jun. 19, 2013.
- © 2013 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication November 21, 2012.
- Accepted for publication March 18, 2013.





{kind=link}
{kind=link}
{kind=link}
{kind=link}