


Abstract
Aromatase is a rate-limiting enzyme for estrogen biosynthesis and has been implicated in pathophysiological states of various diseases via estrogen production. This enzyme is known to be widely distributed in extragonadal and gonadal tissues including the stomach. In contrast to circulating estrogen, the functional role of gastric aromatase/estrogen has not been elucidated in detail, because there is no efficient methodology to investigate spatiotemporal changes of gastric aromatase/estrogen in vivo. Recently, (S)-11C-6-[(4-chlorophenyl)(1H-1,2,4-triazole-1-yl)methyl]-1-methyl-1H-benzotriazole (11C-labeled vorozole), based on a potent nonsteroidal aromatase inhibitor, has been developed as a tracer to investigate aromatase distribution in living animals and humans using a noninvasive PET technique. In the present study, we investigated gastric aromatase expression by means of PET with 11C-vorozole. Methods: After bolus injection of 11C-vorozole into the tail vein, emission scans were obtained for 90 min on male and female rats under isoflurane anesthesia. Displacement studies with unlabeled vorozole and autoradiographic analysis were conducted for demonstration of specific binding. Immunohistochemistry was performed to confirm aromatase expression. Results: PET scans revealed that 11C-vorozole highly accumulated in the stomach and adrenal glands. Displacement studies and autoradiography demonstrated that aromatase was expressed in the stomach but that the accumulation of 11C-vorozole in the adrenal glands might be through nonspecific binding. Immunohistochemical analysis revealed that aromatase is expressed in gastric parietal cells but not in adrenal glands. Moreover, the accumulation of 11C-vorozole in the stomach was significantly increased in fatigued rats. Conclusion: These results suggest that the 11C-vorozole PET technique is a useful tool for evaluation of gastric aromatase dynamics in vivo, which may provide important information for understanding the molecular mechanisms of gastric aromatase/estrogen–related pathophysiological processes and for the development of new drugs.
Aromatase is a key enzyme for the production of sex steroids and catalyzes the final step of the biosynthetic pathway of estrogens. Aromatase has been reported to be widely distributed in extragonadal and gonadal tissues including the ovaries (1), testis (2), placenta (3), adipose tissues (4), skin (5), and brain (6). Aromatase is well known to take part in various important functions, such as cell proliferation and differentiation, through estrogen biosynthesis in an endocrine, autocrine, or paracrine fashion (7). Recently, Ueyama et al., using morphological and biochemical methods, determined that parietal cells in rat gastric mucosa express aromatase and substantially secrete estrogen into the portal vein (8,9). Though gastric aromatase/estrogen has been implicated in several pathophysiological states, such as liver regeneration (10) and feminization (11), by acting as a local regulator of the gastrohepatic axis, the functional role of gastric aromatase/estrogen has still to be elucidated. One of the obstacles to clarification of its role is that gastric estrogen is thought to be trapped and metabolized in the liver and rarely overflows into the systemic circulation (9). Therefore, an adequate method for evaluation of the spatiotemporal changes in gastric aromatase/estrogen in vivo is necessary.
Noninvasive molecular imaging techniques, such as PET, provide powerful tools for quantitative investigation of the tissue-distribution and dynamic changes of functional molecules in vivo, because of their high sensitivity and spatiotemporal resolution. 11C-labeled vorozole, (S)-11C-6-[(4-chlorophenyl)(1H-1,2,4-triazole-1-yl)methyl]-1-methyl-1H-benzotriazole (12), was developed as a PET tracer to investigate the in vivo distribution of aromatase in living animals and humans (13,14). Consistent with previous reports, we have shown that the accumulation of 11C-vorozole was increased in several brain regions of rats and monkeys treated with anabolic steroids (15–18), indicating that 11C-vorozole is a useful molecular probe for aromatase imaging. In the present study, we used 11C-vorozole PET to evaluate gastric aromatase under physiological and pathological conditions.
MATERIALS AND METHODS
Animals
Male and female Sprague–Dawley rats, 8–9 wk old and weighing approximately 300 g, were purchased from Japan SLC, Inc. During the experiment, body temperature was maintained at 37°C. All experimental protocols were approved by the Ethics Committee on Animal Care and Use of RIKEN and were performed in accordance with the Guide for the Care and Use of Laboratory Animals (19).
Chemicals
(S)-11C-6-[(4-chlorophenyl)(1H-1,2,4-triazole-1-yl)methyl]-1-methyl-1H-benzotriazole (11C-vorozole) was synthesized from its N-demethylated derivative via a reaction of N-methylation with 11C-methyl iodide (20). Synthesized 11C-vorozole was purified using reversed-phase high-performance liquid chromatography. Specific radioactivity of 11C-vorozole was 32–49 GBq/μmol, and radiochemical purity was greater than 99.5%. The solvent was removed in vacuo, and the residue was dissolved in saline for injection. Injected volume was 0.5 mL. Unlabeled vorozole was a kind gift from the Janssen Research Foundation. Letrozole was purchased from LKT Labs, Inc.
PET Scans
The rats (n = 6) were anesthetized and maintained using a mixture of 1.5% isoflurane and nitrous oxide/oxygen (7:3) and positioned in the PET scanner gantry (microPET Focus220; Siemens Co., Ltd.). After a bolus injection of 11C-vorozole (injected radioactivity, 95–98 MBq; chemical mass, 0.7–1.5 μg per animal) into the tail vein, a 90-min emission scan was obtained. The abdominal region was scanned for up to 5 min and sorted into 60 × 5 s dynamic signograms. Whole-body scans were then obtained with 180-s duration for 1 bed pass. Emission data were acquired in the list mode. For whole-body scans, the scanner bed was moved continuously in a reciprocating motion to ensure scanning of the entire rat body, and the data were sorted into dynamic signograms for every 1 bed pass. Emission data were reconstructed by Fourier rebinning and maximum likelihood expectation maximization. For the abdominal scans, the radioactivity concentrations in the region of interest were expressed as standardized uptake value or percentage injected dose. For the whole-body scans (continuous bed-motion acquisition), the radioactivity in the region of interest was estimated by percentage of total radioactivity at each frame.
Displacement Studies
The rats were anesthetized and maintained using a mixture of 1.5% isoflurane and nitrous oxide/oxygen (7:3), and 11C-vorozole (injected radioactivity, 96–100 MBq; chemical mass, 0.9–1.7 μg per animal) was administered by bolus injection into the tail vein. Fifteen minutes after 11C-vorozole administration, unlabeled vorozole (100 μg/kg; n = 3) dissolved in 4% ethanol or vehicle (n = 3) was injected into the tail vein as a bolus. The abdominal region was scanned for 90 min (33 × 5, 55 × 30, and 20 × 180 s; a total of 108 frames). For the displacement studies with the other aromatase inhibitor, 11C-vorozole was injected into a tail vein as a bolus in coadministration with letrozole (1 mg/kg) or vehicle.
Radiometabolite Analysis
Radiometabolite analysis of rat stomach and adrenal gland was performed. At 30 min after intravenous injection of 11C-vorozole, the blood flow was terminated, and then the tissues were quickly removed and homogenized. Sample preparation and chromatographic separation was performed according to previously described procedures with slight modification (18).
Autoradiography
Frozen rat brains (n = 3), stomachs, and adrenal glands were sliced into serial coronal sections (25 μm) using a cryostat microtome, and the sections were thaw-mounted onto glass slides. Consecutive sections were preincubated at room temperature (∼22°C) for 10 min in phosphate-buffered saline (pH 7.4) and incubated with 2 nM 11C-vorozole in the presence or absence of unlabeled vorozole (1 μM) at room temperature for 30 min. The sections were washed for 2 min in the buffer 3 times and rapidly air-dried. The sections were exposed on imaging plates (BAS-SR-2040; Fujifilm) at room temperature for 40 min to generate the autoradiograms. FLA-7000 (Fujifilm) was used for scanning the imaging plates.
Immunohistochemistry
Rats (n = 3) were perfused with 4% formaldehyde buffered with 0.1 M phosphate-buffered saline under deep anesthesia. The stomach and adrenal glands were removed and further fixed in the same fixative at 4°C for 24 h. Frozen sections (10 μm thick) were prepared in a cryostat microtome and thaw-mounted on Superfrost glass slides (Matsunami Glass). The immunohistochemical staining procedure was described in the previous reports (17). After immunohistochemical detection, the slides were stained with hematoxylin and eosin.
Fatigue-Loading Procedure
The fatigued condition in rats was induced using the method previously established by our group (21). Briefly, rats (male, n = 4) were transferred to cages filled with water (23°C ± 1°C) to a height of 2.2 cm and kept in the water cage for 5 d. The water was changed once a day. Compared with the control group, rectal temperatures remained within the normal range in these rats during fatigue-loading, and no gastric ulcers were observed up to the end of fatigue-loading. Using this animal model of comprehensive fatigue, which encompasses both physical and mental fatigue, we previously reported a decrease in physical performance (21), alternation of the expression levels of peptidases in the pituitary gland (22,23), and significant changes in amino acid metabolism in the plasma (24). Such fatigue-induced changes in behavior and biochemical indicators were completely reversed after a return to ordinary conditions.
Data Analysis
All values are presented as the mean ± SEM. Two-way ANOVA with Scheffe multiple-comparison procedure was used to test for differences in temporal changes in time–activity curves between displacement and vehicle-treated groups. A 2-tailed unpaired t test was used to test for statistical difference in radioactivity between fatigued and control rats. The differences discussed in the text were considered to be significant at a value of P less than 0.05.
RESULTS
Whole-Body PET Studies
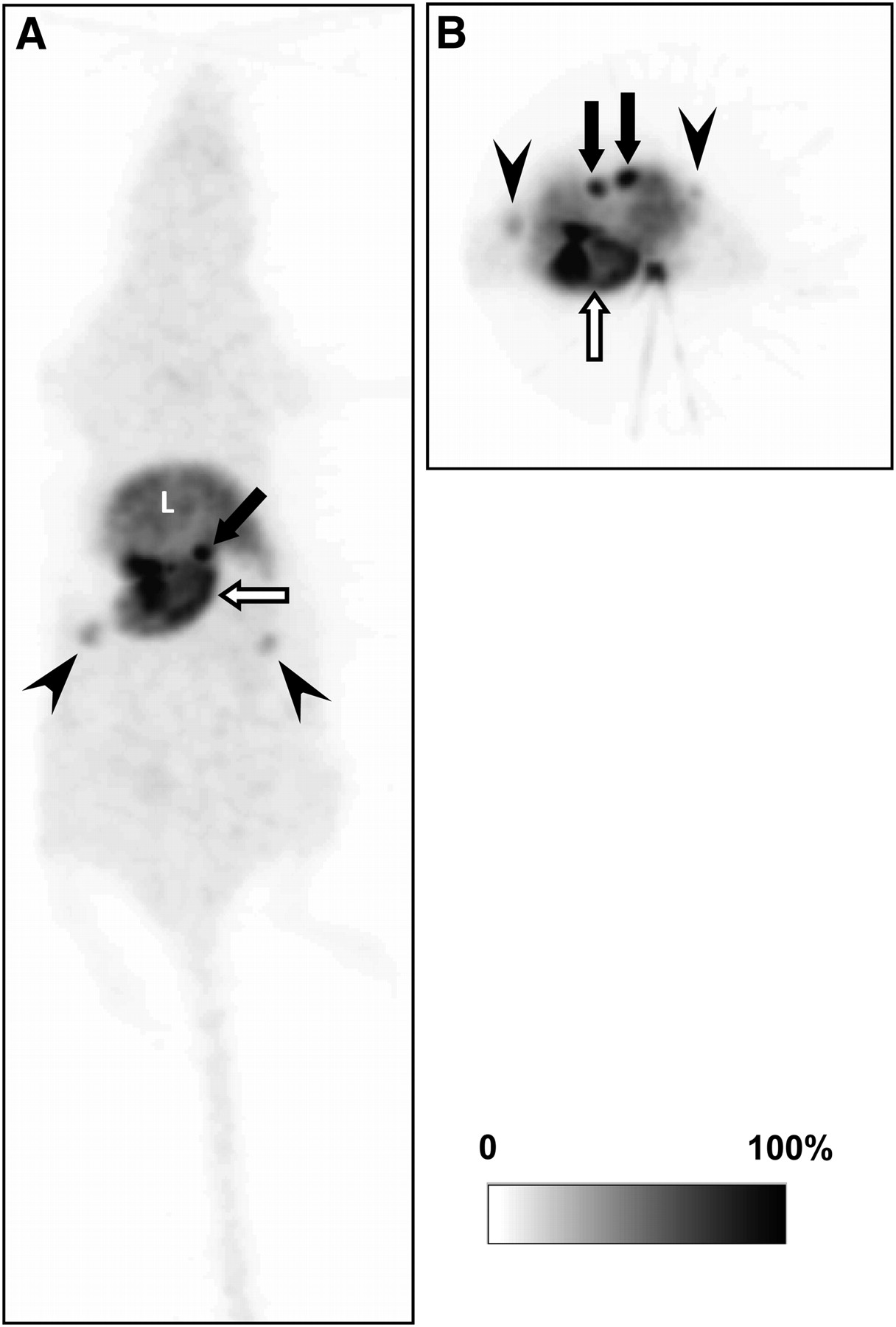
A representative whole-body maximum-intensity-projection image after bolus injection of 11C-vorozole is shown in Figure 1. High accumulation of 11C-vorozole was detected in the stomach and the adrenal glands (Fig. 1). In the female rats, the radioactivity was also observed in the ovarium (Fig. 1). Detectable 11C-vorozole radioactivity was observed in the testis of male rats (data not shown). The temporal changes in radioactivity of 11C-vorozole in these tissues were variable. 11C-Vorozole accumulation in the stomach increased until the end of the PET scans but accumulation in other tissues reached a peak and then gradually decreased thereafter. The maximum radioactivity detected in the stomach was 4.31% of the total radioactivity (at the end of the PET scan, 90 min).
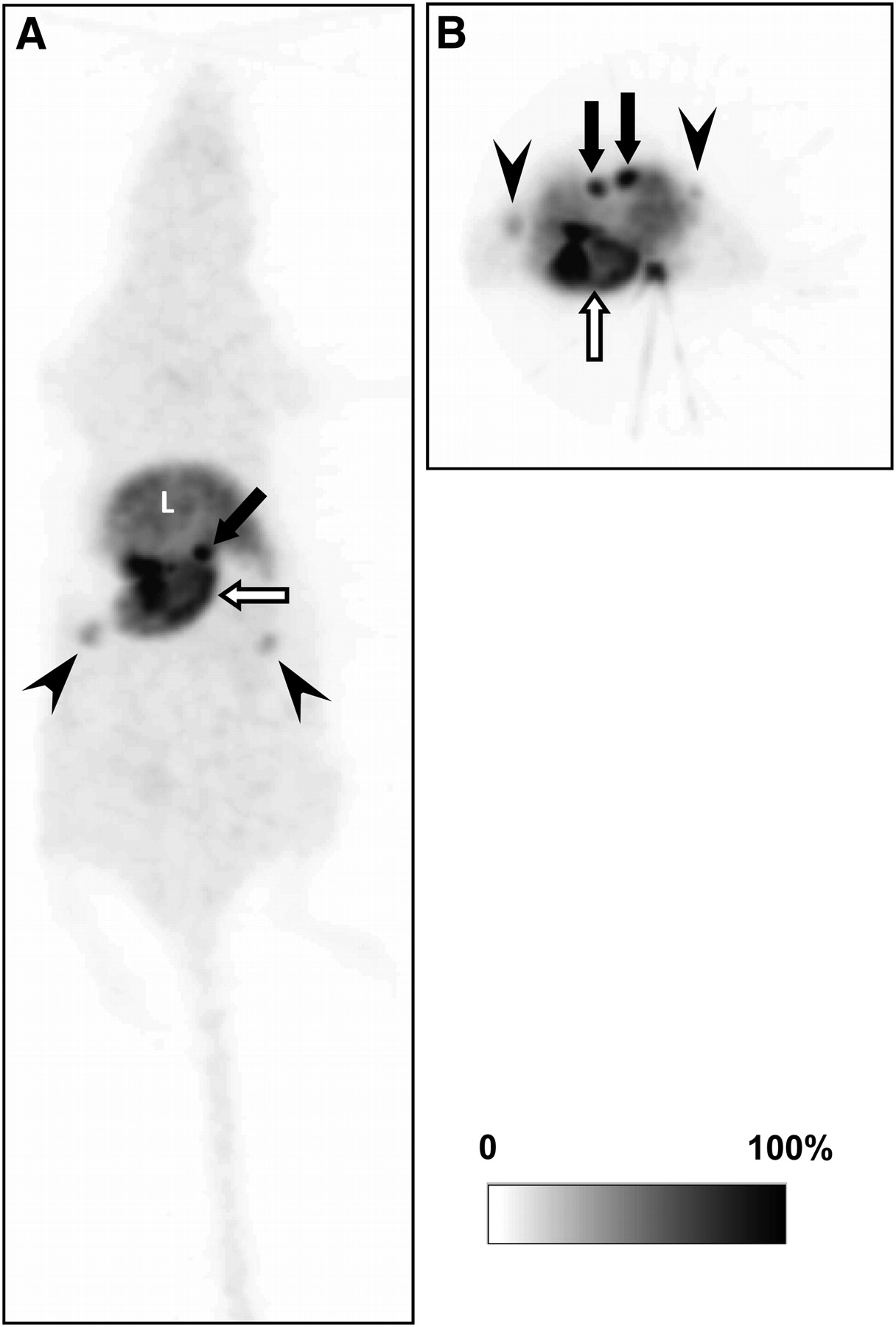
Representative whole-body maximum-intensity-projection image of 11C-vorozole in female rat: coronal (A) and transverse (B) images. 11C-vorozole was highly accumulated in stomach (open arrows), adrenal glands (closed arrows), and ovarium (arrowheads). L = liver.
Displacement Studies
The radioactivity of 11C-vorozole in the stomach immediately decreased after administration of unlabeled vorozole 15 min after bolus injection of 11C-vorozole (Fig. 2A). The mean values of radioactivity in the stomach were significantly decreased in the unlabeled ligand–treated group (P < 0.05, 2-way ANOVA with Scheffe multiple-comparison procedure). However, radioactivity levels in the adrenal glands were not changed by administration of unlabeled vorozole (Fig. 2B). The plasma concentration–time curve showed no difference between the displacement and the control group, suggesting that the metabolic and excretion pathway had not been influenced by injection of unlabeled vorozole (Supplemental Fig. 1; supplemental materials are available online only at http://jnm.snmjournals.org). In addition, coadministration with unlabeled letrozole (1 mg/kg), the other aromatase inhibitor, decreased the distribution of the radioactivity in the stomach wall (Supplemental Fig. 2). Ex vivo autoradiographic images also confirmed that the radioactivity predominantly accumulated in the gastric gland region was drastically decreased by coadministration with letrozole, whereas that in adrenal glands was not significantly affected by the administration of letrozole (Supplemental Fig. 3).
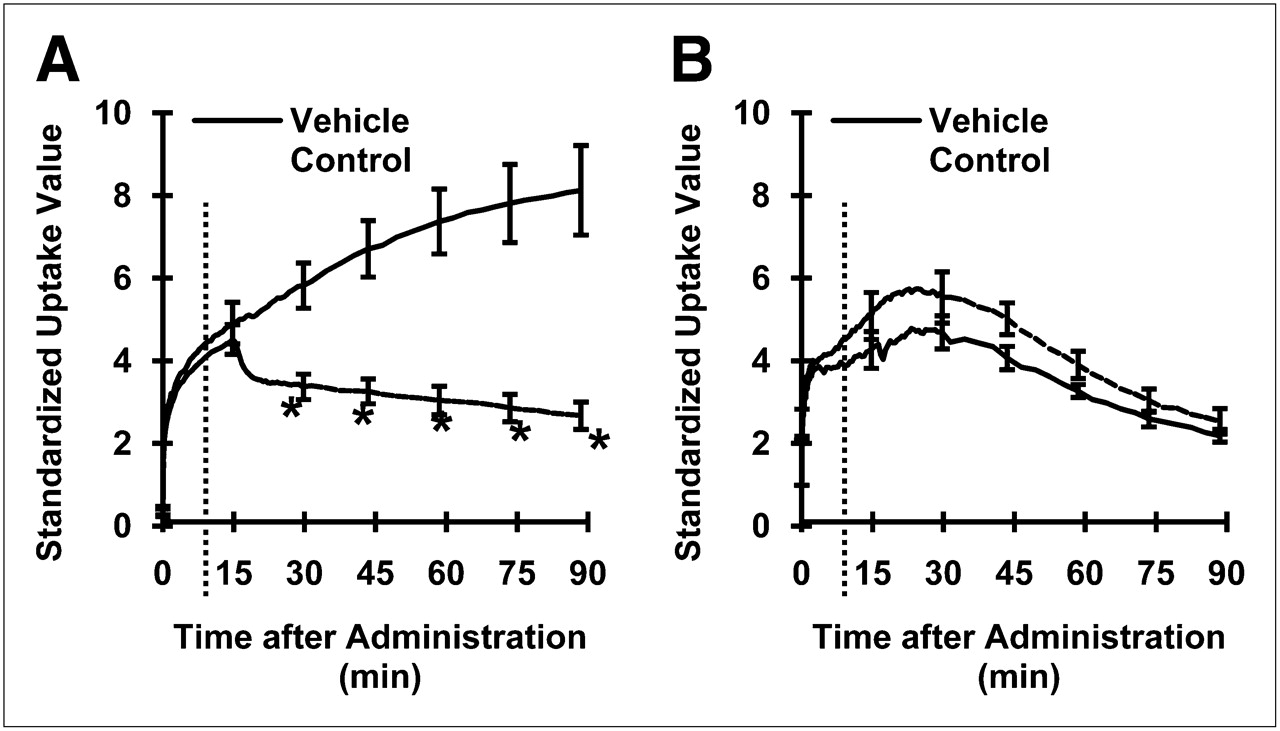
Time–activity curves for 11C-vorozole displaced by excess unlabeled vorozole in stomach (A) and adrenal glands (B). Unlabeled vorozole or vehicle was administered 15 min after start of PET scans. Broken line indicates time at which unlabeled ligand was administered. Asterisks indicate significant difference (P < 0.05, 2-way ANOVA with Scheffe multiple-comparison procedure).
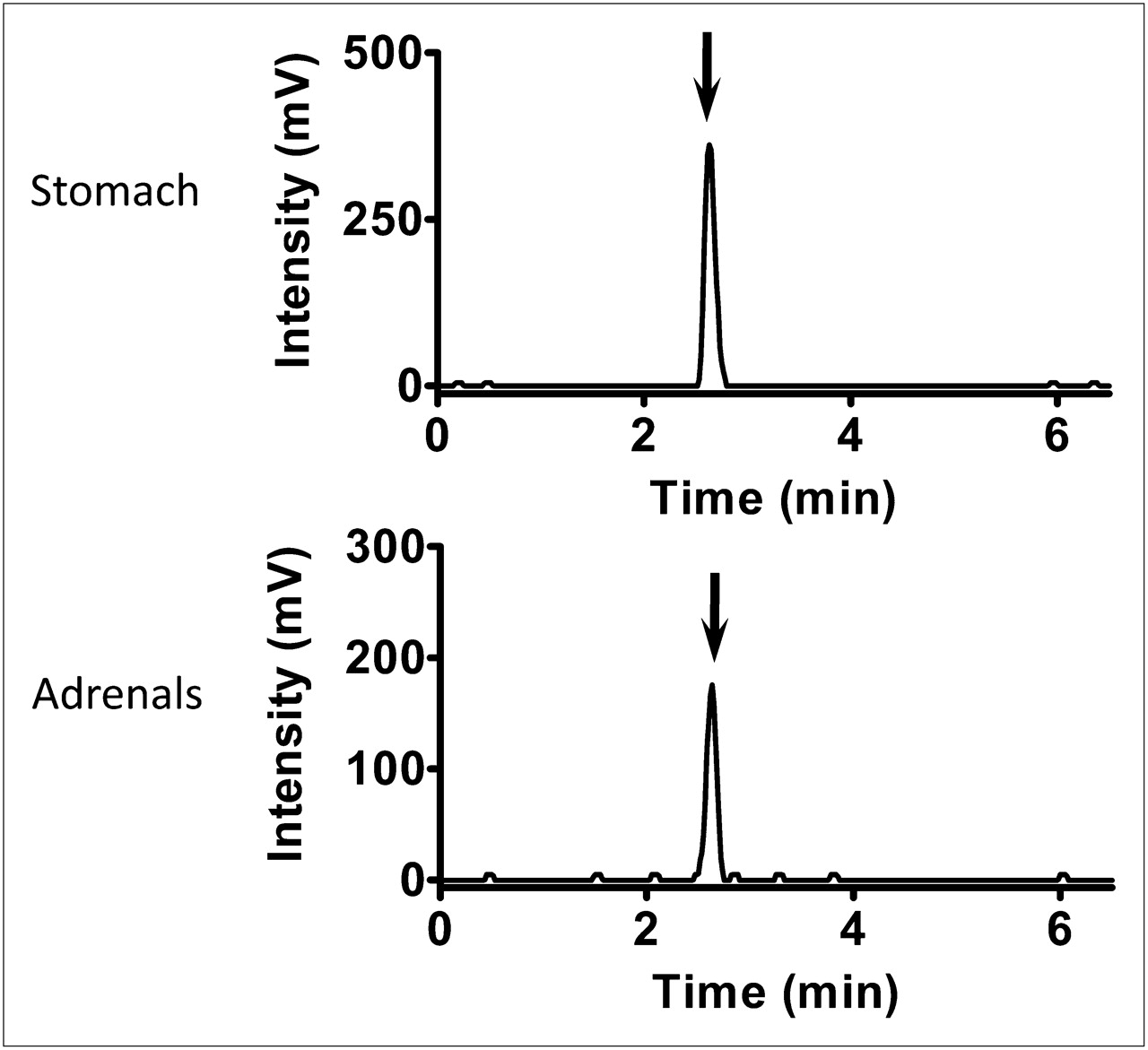
Analysis of radiochromatograms revealed that almost 100% of the radioactivity in these tissues was derived from 11C-vorozole (Fig. 3).

Representative high-performance liquid radiochromatograms of stomach and adrenal glands extracts after intravenous administration of 11C-vorozole (∼300 MBq/kg).
Autoradiography and Immunohistochemistry
Autoradiographic images of tissue sections of rat stomach, brain, and adrenal glands are shown in Figure 4. As reported previously (14), the binding of 11C-vorozole to the brain amygdala was clearly displaced by unlabeled vorozole. In a similar fashion, the strong signal of 11C-vorozole in the stomach was clearly eliminated by the addition of unlabeled vorozole. However, addition of unlabeled vorozole had no effect on the 11C-vorozole signal in the adrenal glands. Immunohistochemical studies were then performed to confirm aromatase expression. The immunolocalization of aromatase in the stomach is shown in Figure 5; aromatase immunosignals were observed in the eosinophilic cytoplasm of the gastric parietal cells that are arranged in the glandular body of fundic glands. In the adrenal glands, however, no aromatase immunosignals were detected either in the adrenal cortex or in the adrenal medulla (Fig. 5).
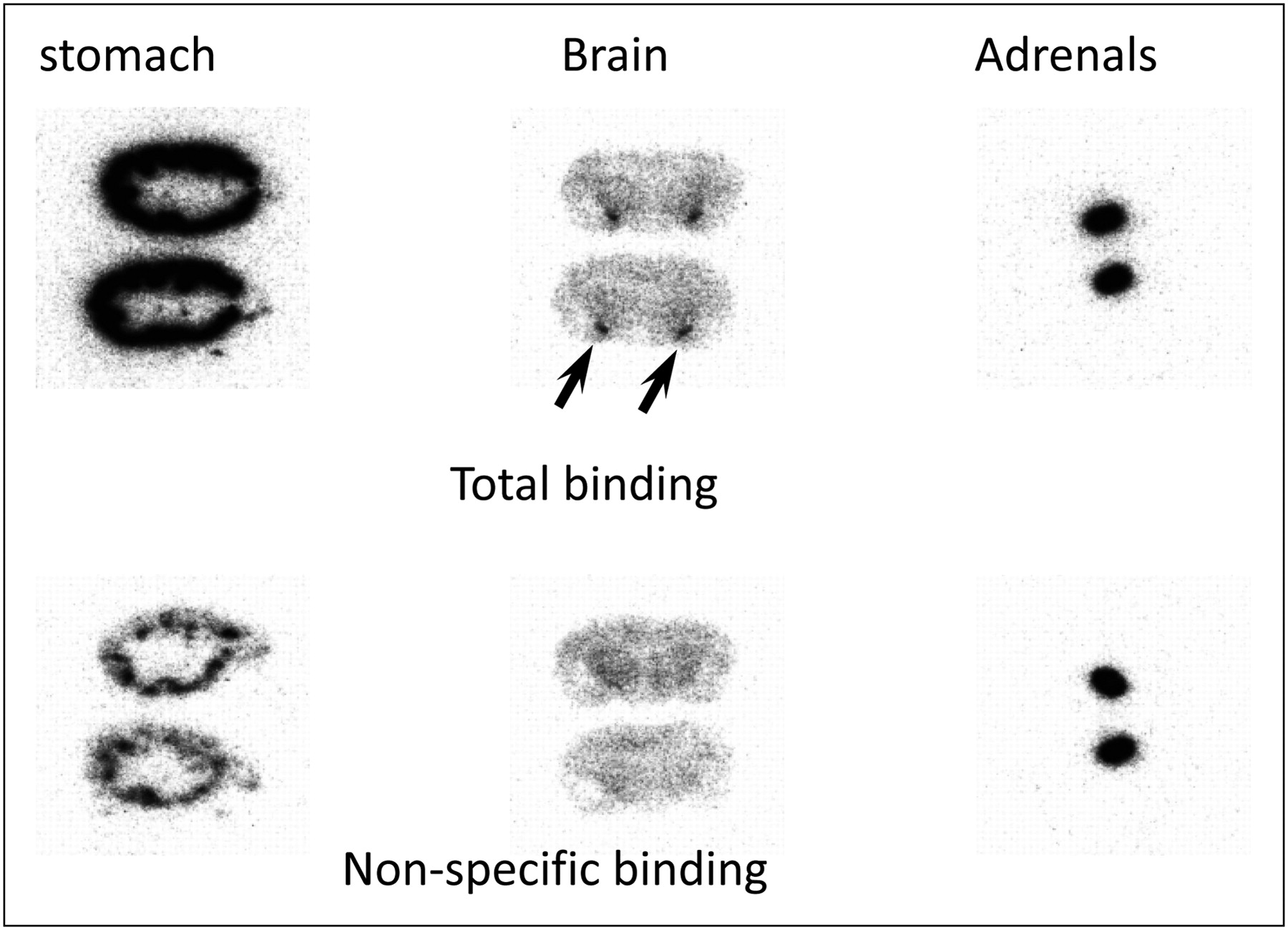
Autoradiographic images of displacement of 11C-vorozole with unlabeled vorozole in tissue slices. Autoradiographic image of tissue sections of stomach, brain, and adrenal glands that were incubated with 2 nM 11C-vorozole in absence (total binding) or presence (nonspecific binding) of 1 μM unlabeled vorozole. Arrows indicate amygdala.

Photomicrographs of aromatase immunoreactivity in stomach and adrenal gland. Tissue sections of stomach (A and B) and adrenal gland (C and D) were analyzed for aromatase immunoreactivity. Scale bars indicate 100 μm in A, 50 μm in B, 1 mm in C, and 500 μm in D. Arrows in B indicate aromatase-immunopositive gastric parietal cells. LM = lamina muscularis mucosae; SM = submucosa; TM = tunica muscularis.
PET Studies in Fatigued Rats
For further investigation of the pathophysiological role of gastric aromatase, we focused on the regulatory effect of gastric estrogen on ghrelin production. Ghrelin, an endogenous ligand for the growth hormone secretagogue receptor, is predominantly produced in the stomach (25), and its expression and release are thought to be regulated by gastric estrogen in a paracrine fashion (26). Recently, ghrelin levels have been observed to be upregulated in a chronic stress model in rats (27), which was originally developed by our group as a comprehensive fatigue model (21). To confirm whether gastric aromatase is also upregulated in such a fatigued condition, we performed 11C-vorozole PET studies in these fatigued rats. Consistent with our expectations, the accumulation of 11C-vorozole in the stomach was increased in the fatigued group (n = 4), compared with the control (n = 3) group (Fig. 6). The radioactivity in the stomach was significantly increased in the fatigued group (P < 0.05, 2-tailed unpaired t test).

11C-vorozole radioactivity levels are increased in stomach of fatigued rat. Representative transverse image of 11C-vorozole in control (A) and fatigued (B) rats from 5 to 90 min after bolus injection of 11C-vorozole. Closed arrow indicates stomach. (C) 11C-vorozole accumulation in stomach of control (n = 3) and fatigued (n = 4) rats. Asterisk indicates a significant difference (P < 0.05, 2-tailed unpaired t test).
DISCUSSION
The present study showed that aromatase is abundantly expressed in the stomach and is upregulated under conditions of severe fatigue, using an in vivo molecular imaging technique with 11C-vorozole. The PET study with 11C-vorozole clearly showed that the radioactivity of 11C-vorozole was highly accumulated in the stomach and adrenal glands (Fig. 1). The displacement experiments performed in vivo revealed that postadministration (15 min after bolus injection of 11C-vorozole) of the unlabeled compound induced a significant decrease in the level of radioactivity in the stomach but not in that in the adrenal glands (Fig. 2). The radioactivity in the stomach and adrenal glands was derived from 11C-vorozole (Fig. 3). Considering the fact that vorozole inhibits aromatase activity by reversibly binding to its heme domain (28,29), these results strongly suggest that the accumulation of 11C-vorozole in the stomach, but not in the adrenal glands, reflects the specific binding. These results were further supported by the in vitro autoradiographic analysis (Fig. 4). Moreover, the immunohistochemical and morphological studies also demonstrated that aromatase immunoreactivity was abundantly observed in the cytoplasm of gastric parietal cells (Fig. 5). Recently, Ueyama et al. (8,9), using immunohistochemistry and in situ hybridization, reported that aromatase messenger RNA and protein are localized in gastric parietal cells. They further noted that the production of estrone and 17β-estradiol from androstenedione in the gastric mucosa of the rat stomach might be catalyzed by endogenous aromatase that is localized in the stomach (8). The combined observations suggest that endogenous aromatase is expressed in gastric parietal cells and that the 11C-vorozole PET technique is a powerful tool for noninvasive evaluation of gastric aromatase.
The second major finding of the present study is that the level of 11C-vorozole radioactivity was significantly increased in the stomach of the fatigued rat, indicating upregulation of gastric aromatase under this pathological condition. Although a functional role for gastric aromatase and estrogen has not been elucidated in detail, a regulatory effect of gastric estrogen on ghrelin production has been proposed by Sakata et al. (26). Ghrelin, an endogenous ligand for the growth hormone secretagogue receptor, is known to be predominantly produced in the stomach (25). Sakata et al. demonstrated that ghrelin-producing cells are located close to gastric parietal cells and that gastric estrogen directly induces ghrelin production (26). Ghrelin plays important roles in growth regulation and energy homeostasis such as in stimulation of growth hormone release, in food intake, and in adiposity (30,31). The plasma level of ghrelin is known to be increased in response to energy insufficiency and subsequently induces a potent feeding response via activation of the growth hormone secretagogue receptor (32). In our fatigued model, the rat was kept in a water cage for 5 d to induce psychological and physical exhaustion, during which energy consumption would be increased. Indeed, an increase in plasma ghrelin levels has been reported in these fatigued rats (27). These observations suggest that an increase in gastric aromatase might be involved in energy homeostasis via regulation of gastric ghrelin production. However, the detailed mechanism of such regulation remains to be investigated.
Some recent reports noted that aromatase is localized in the adrenal glands. Conley et al. (33) reported that aromatase is expressed in the adrenal glands of fetal and newborn pigs. In their report, aromatase was detected in the adrenal cortex using immunohistochemistry. Illera et al. (34) reported that aromatase was distributed in the rat adrenal medulla, based on immunohistochemical analysis. However, using messenger RNA analysis, Ueyama et al. (8,9) detected no expression of the aromatase gene in rat adrenal glands. In the present study, an obvious accumulation of 11C-vorozole was observed in adrenal glands. However, this accumulation of 11C-vorozole was not displaced by unlabeled vorozole either in vitro or in vivo. The present immunohistochemical studies were also unable to detect aromatase immunoreactivity in adrenal glands. These results suggest that 11C-vorozole might nonspecifically bind to molecules other than aromatase in adrenal glands.
Because of the beneficial effect of aromatase inhibitor treatment for estrogen-related diseases, several aromatase inhibitor compounds have been widely used as therapeutic agents (35,36). For instance, aromatase inhibitors have been used for the treatment of breast cancer in postmenopausal women, in which the inhibitor target is aromatase that is expressed by the breast cancer (28). To date, gastric aromatase has not attracted much attention in terms of drug development targeted toward estrogen-related disease. However, it is known that one of the most common adverse events of aromatase inhibitors is gastrointestinal disorders, such as nausea, vomiting, and anorexia (37,38). The present study clearly demonstrated that aromatase is abundantly expressed in gastric parietal cells and interacted with 11C-vorozole that was administrated intravenously. Obviously, gastric aromatase is not the target of breast cancer; an inadequate inhibition of gastric aromatase is not desirable for preventing adverse effects of aromatase inhibitors. These observations suggest that development of a technology such as 11C-vorozole PET that can evaluate gastric aromatase/estrogen levels is necessary for drug development targeted toward estrogen-related diseases.
CONCLUSION
This study is the first time to evaluate gastric aromatase in vivo using quantitative PET with 11C-vorozole and showed that aromatase is abundantly expressed in the stomach and is upregulated in the fatigued rat. These results suggest that 11C-vorozole PET enables noninvasive investigation of aromatase dynamics in vivo, possibly providing important information for understanding the functional role of gastric aromatase/estrogen in the pathophysiological process and for the development of new drugs targeted toward estrogen-related diseases.
DISCLOSURE STATEMENT
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
This work was supported in part by consignment expenses from the Molecular Imaging Program on Research Base for Exploring New Drugs from the Ministry of Education, Culture, Sports, Science, and Technology (MEXT), Japanese Government. No other potential conflict of interest relevant to this article was reported.
Footnotes
Published online Nov. 9, 2011.
- © 2011 by Society of Nuclear Medicine
REFERENCES
- Received for publication December 25, 2010.
- Accepted for publication August 8, 2011.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.