


Abstract
Fatty acid oxidation (FAO) is a major energy-providing process with important implications in cardiovascular, oncologic, neurologic, and metabolic diseases. A novel 4-thia oleate analog, 18-18F-fluoro-4-thia-oleate (18F-FTO), was evaluated in relationship to the previously developed palmitate analog 16-18F-fluoro-4-thia-palmitate (18F-FTP) as an FAO probe. Methods: 18F-FTO was synthesized from a corresponding bromoester. Biodistribution and metabolite analysis studies were performed in rats. Preliminary small-animal PET studies were performed with 18F-FTO and 18F-FTP in rats. Results: A practical synthesis of 18F-FTO was developed, providing a radiotracer of high radiochemical purity (>99%). In fasted rats, myocardial uptake of 18F-FTO (0.70 ± 0.30% dose kg [body mass]/g [tissue mass]) was similar to that of 18F-FTP at 30 min after injection. At 2 h, myocardial uptake of 18F-FTO was maintained, whereas 18F-FTP uptake in the heart was 82% reduced. Similar to 18F-FTP, 18F-FTO uptake by the heart was approximately 80% reduced at 30 min by pretreatment of rats with the CPT-I inhibitor etomoxir. Folch-type extraction analyses showed 70–90% protein-bound fractions in the heart, liver, and skeletal muscle, consistent with efficient trafficking of 18F-FTO to the mitochondrion with subsequent metabolism to protein-bound species. Preliminary small-animal PET studies showed rapid blood clearance and avid extraction of 18F-FTO and of 18F-FTP into the heart and liver. Images of 18F-FTO accumulation in the rat myocardium were clearly superior to those of 18F-FTP. Conclusion: 18F-FTO is shown to be a promising metabolically trapped FAO probe that warrants further evaluation.
The oxidation of long-chain fatty acids is the major aerobic, energy-producing process in the heart, liver, and skeletal muscle. Abnormalities of fatty acid oxidation (FAO) are associated with a variety of diseases, including ischemic heart disease (1), heart failure (2), cardiomyopathy (3), oncology (4), neurodegeneration (5), alcoholism (6), nonalcoholic fatty liver (7), obesity or metabolic syndrome (8–10), and diabetes (8,11).
Over the last 20 y, our laboratory has investigated radiolabeled thia-substituted long-chain fatty acid analogs as metabolically trapped probes of myocardial FAO (12–17). The first-generation molecule 14-18F-fluoro-6-thia-heptadecanoic acid (18F-FTHA) was a 6-thia analog that showed excellent myocardial imaging properties but lacked specificity for mitochondrial FAO in certain conditions such as oxygen insufficiency (14,15). Accumulation of 18F-FTHA in the liver and skeletal muscle was largely in complex lipid pools and therefore not useful for the indication of mitochondrial FAO in those tissues (16). The second-generation FAO probe 16-18F-fluoro-4-thia-palmitate (18F-FTP), a 4-thia fatty acid analog, improved on 18F-FTHA by increasing specificity to myocardial FAO, particularly in response to hypoxia (12,14). However, myocardial retention of 18F-FTP was suboptimal, and metabolic trapping in the liver was not specific to mitochondrial function (14).
Oleate is one of the most prevalent long-chain fatty acids in human plasma (17,18) and, like palmitate, is used as an energy substrate or incorporated into various complex lipids for storage or as cellular structural components. Although oleate and palmitate have been shown to be similarly taken up and oxidized in human myocardium (18), their use in the whole body and skeletal muscle in humans has shown some notable differences (19–22). Because dietary oleate (18:1) is preferentially oxidized relative to palmitate (16:0) and stearate (18:0) (22), we anticipated that an oleic acid analog of 18F-FTP might show a higher specificity for mitochondrial FAO than 18F-FTP itself. Therefore, we synthesized and preliminarily evaluated in rats the 4-thia substituted oleate analog 18-18F-fluoro-4-thia-oleate (18F-FTO) (1) as a novel PET probe of FAO (Fig. 1). Results of biodistribution and small-animal PET studies were compared with those for the previously developed 4-thia–substituted palmitate analog 18F-FTP, showing enhanced myocardial imaging characteristics and increased specificity for evaluation of FAO rates in vivo.

Structures of 18F-labeled thia fatty acids developed as FAO probes.
MATERIALS AND METHODS
Synthesis of Radiotracers
The chemical structures for the 18F-labeled thia fatty acids that were synthesized and evaluated—18F-FTP and 18F-FTO—are shown in Figure 1. The synthetic scheme is shown in Figure 2. Details of the synthesis of the radiolabeling precursor and reference standard of FTO are available in the supplemental materials (online only at http://jnm.snmjournals.org).

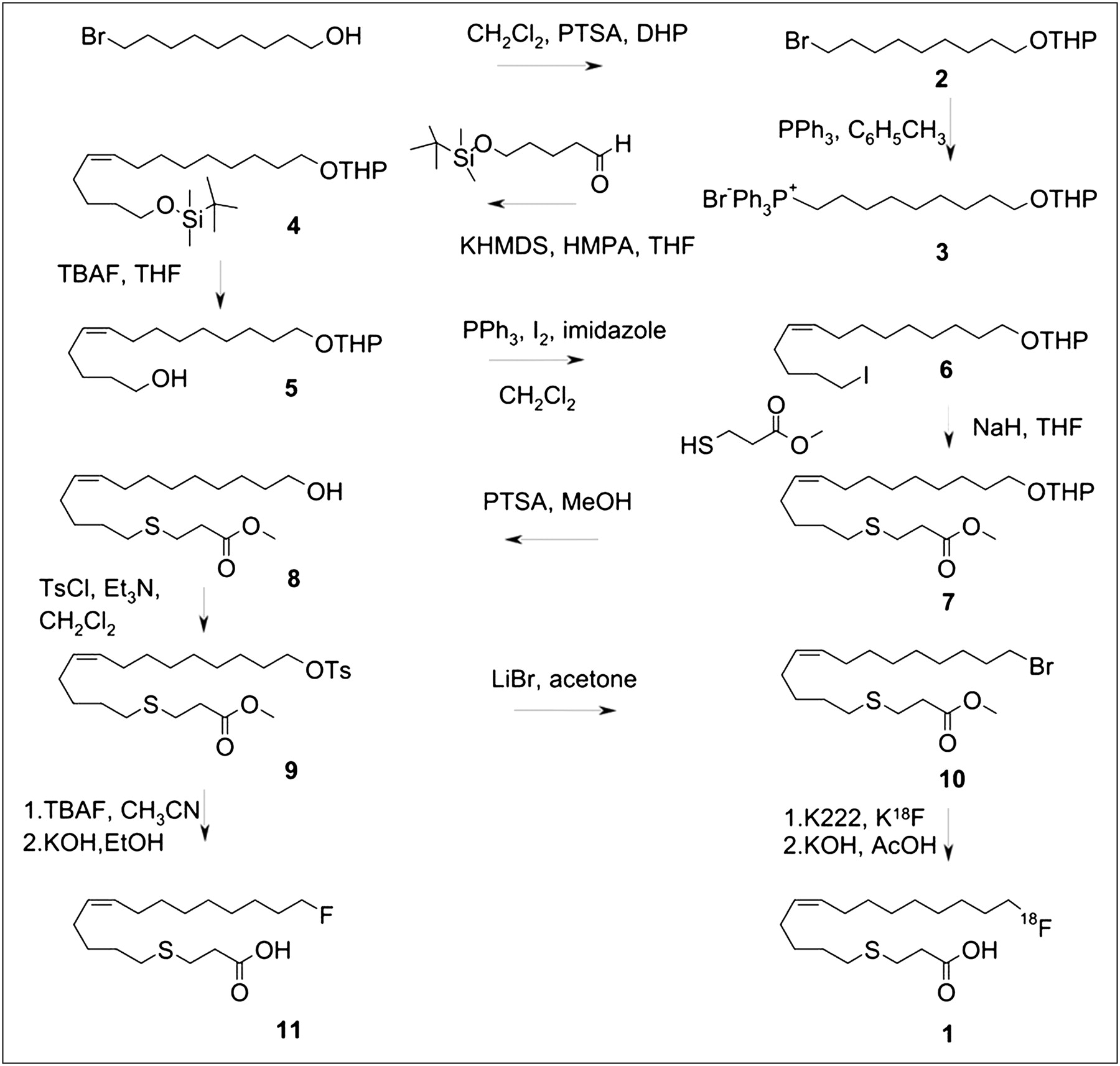
Synthetic scheme for 18F-FTO (1), FTO reference standard (11), and labeling precursor (10).
18F-Labeling Procedure
The labeling precursors for 18F-FTO and 18F-FTP were the methylesters of their corresponding bromides (Fig. 2). Nucleophilic 18F-fluorination and subsequent hydrolysis of the ester protective group were performed as previously described (14). Briefly, cyclotron-produced 18F-fluoride was dried down under nitrogen at 95°C in a 2-mL glass vial containing Kryptofix 2.2.2 (10 mg), acetonitrile (0.8 mL), and K2CO3 (4 mg) solution in water (0.15 mL). The residue was further dried by azeotropic distillation with acetonitrile (2 × 0.5 mL). A solution of the precursor (∼2 mg) in acetonitrile (0.5 mL) was added, and the vial was sealed and heated at 75°C for 15 min. The vial was placed in ice water for brief cooling. Subsequent hydrolysis of the resulting 18F-fluoroester was performed in the same vessel by adding 0.15 mL of 0.2N KOH to the solution and heating it at 90°C for 4 min. The mixture was cooled, acidified with concentrated acetic acid (25 μL), filtered, and applied to the semipreparative high-performance liquid chromatography (HPLC) column (Luna C-18, 5 μm, 10 × 250 mm; Phenomenex). The mobile phase was acetonitrile/water/trifluoroacetic acid (90:10:0.005 v/v), and flow rate was 5 mL/min. An in-line ultraviolet detector (210 nm) was used to monitor the elution of unlabeled materials. The 18F-fluoro–fatty acid fraction was diluted in 50 mL of water and trapped on a C-18 Sep-Pak cartridge (Accel C-18 Plus; Waters), followed by washing of the Sep-Pak with 10 mL of water. The product was eluted from the Sep-Pak in 1 mL of ethanol. The volume of the ethanol solvent was reduced to approximately 0.1 mL under a nitrogen stream at 40°C and then formulated in 0.5–1% albumin in isotonic NaCl solution and filtered through a 0.22-μm filter (Millex-GS; Millipore). Radiochemical purity was checked by radio-HPLC in the system described in the supplemental materials or by radio–thin-layer chromatography (silica gel; hexane/ethyl acetate/trifluoroacetic acid, 75:25:0.05 [v/v]; Rf = 0.4).
Biodistribution Studies in Rats
Female Sprague–Dawley rats (165–215 g; Charles River) were fasted overnight or fed ad libitum. The rats were anesthetized with isoflurane (3.5% induction, 1.5% maintenance). The 18F-labeled radiotracer (0.7–1.4 MBq) was injected into the tail vein. The heart, liver, lung, blood, kidney, bone (femur), brain, and skeletal muscle were procured after a prescribed time. The tissues were counted for 18F radioactivity and weighed. Radiotracer uptake was calculated as:
In 1 group of fasted rats, the CPT-I inhibitor etomoxir (40 mg/kg; Sigma-Aldrich) was administered intraperitoneally 2 h before 18F-FTO injection. Crude analysis of the nature of the metabolites in the heart, liver, and muscle was performed by a Folch-type extraction procedure. Approximately 0.5 g of tissue was excised and thoroughly homogenized and sonicated (20 s) in 7 mL of chloroform/methanol (2:1) at 0°C. Urea (40%, 1.75 mL) and 5% sulfuric acid (1.75 mL) were added, and the mixture was sonicated for an additional 20 s. After centrifugation for 10 min at 1,800g, aqueous, organic, and protein interphase (pellet) fractions were separated and counted for 18F radioactivity.
Small-Animal PET/CT Studies in Rats
Two female Sprague–Dawley rats (167–172 g) were fasted overnight. The rats were anesthetized with isoflurane (3.5% induction, 1.5% maintenance). An eXplore Vista CT scanner (GE Healthcare) was used. Before the radiotracer imaging study, a whole-body CT scan was obtained. This scan was used for anatomic visualization and attenuation correction of the emission data. 18F-FTO and 18F-FTP were studied in 2 different rats to allow biodistribution studies to be performed after the PET scan. The 18F-labeled radiotracer (9–11 MBq) was injected into the tail vein, and the emission data were collected in dynamic acquisition for 50 min over the upper thorax, followed by a static whole-body scan from 55 to 115 min after injection. The dynamic images were analyzed to give time–activity curves of the left-ventricular myocardium, blood pool, lung, liver, and bone (rib) at the following consecutive intervals: 6 × 10, 4 × 30, 5 × 60, and 8 × 300 s. The time–activity curves were expressed as the commonly used standardized uptake value, which is the PET-measured radioactivity concentration (Ct) normalized to body weight and injected dose:
Statistical Analyses
All results were expressed as the mean ± SD. Comparison of the means was made using a 2-tailed Student t test. The acceptable level of significance was set at a P value of less than 0.05.
RESULTS
Synthesis of 18F-FTO (1)
Figure 2 shows the synthetic scheme. To make (Z)-18-fluoro-4-thia-octadec-9-enoic acid (11) (FTO), 1-bromononanol was converted to 2-(9-bromononyloxy)tetrahydro-2H-pyran (2), which on subsequent reaction with triphenylphosphine gave [9-((tetrahydro-2H-pyran-2-yl)oxy)nonyl]triphenylphosphonium bromide (3) as an ylide for the Wittig reaction. The ylide precursor was treated with 5-(tert-butyldimethylsilyloxy) pentanal, giving (Z)-tert-butyldimethyl(14-(tetrahydro-2H-pyran-2-yloxy)tetradec-5-enyloxy)silane (4). The formation of compound 4 was characterized by the presence of olefinic protons at δ 5.38–5.32 (m, 2H) and carbons at δ 130.1, 129.6 in their nuclear magnetic resonance spectra. Compound 4 was further treated with tetrabutyl ammonium fluoride in tetrahydrofuran to deprotect the hydroxyl group. On iodination, hydroxyl analog 5 gave compound 6. Methyl 3-mercaptopropanoate was then treated with compound 6 in tetrahydrofuran using sodium hydride to obtain (Z)-methyl 18-(tetrahydro-2H-pyran-2′-yloxy)-4-thia-octadec-9-enoate (7). The tetrahydropyran group in compound 7 was removed using p-toluenesulfonic acid in methanol. The obtained hydroxyl analog (8) was further reacted with tosyl chloride to obtain the corresponding tosylester (9), which on reaction with lithium bromide gave (Z)-methyl 18-bromo-4-thia-octadec-9-enoate (10), the optimal bromoester precursor for radiofluorination. The cold fluorinated standard (11) was also synthesized from compound 9. The intermediates and the final precursor were characterized by detailed spectroscopic analyses.
Radiofluorination of the bromoester (10) proceeded efficiently, with more than 70% incorporation of 18F-fluoride into the corresponding 18F-fluoroester. Hydrolysis of the methylester group by the addition of 0.2N KOH and continued heating was quantitative, and overall radiochemical yields after preparative HPLC isolation were in the 20–30% uncorrected range and not further optimized. Radiochemical purity of the product (1) by radio-HPLC or radio–thin-layer chromatography was more than 99%. The product was effectively formulated in 0.5–1.0% bovine serum albumin solution in saline and sterile-filtered before being administered to rats. Likewise, the palmitate analog, 18F-FTP, was synthesized by a similar approach as previously described (14).
Biodistribution Studies in Rats
Table 1 shows the biodistribution of 18F radioactivity in rats after intravenous administration of 18F-FTO at 30 and 120 min after injection. In both fasted and fed rats at 30 min, the highest uptake was seen in the liver, followed by the heart, kidney, and other organs. The only difference in biodistribution between fasted and fed rats was a 29% lower uptake in the livers of fed animals. Clearance from the liver was seen between 30 and 120 min in fasted animals, whereas heart uptake was maintained at 0.7–0.8% dose kg (body mass)/g (tissue mass). Pretreatment of fasted rats with the CPT-I inhibitor etomoxir resulted in an 80% reduction in myocardial uptake of 18F-FTO (P < 0.02), consistent with the previously reported decrease of 18F-FTP uptake by rat heart under similar experimental conditions (14). Bone uptake of 18F-FTO increased from 30 to 120 min after injection, evidencing ongoing defluorination, similar to that of 18F-FTP.
Uptake (% Dose kg/g) of 18F-FTO and 18F-FTP in Sprague–Dawley Rats
In fasted rats, myocardial uptake of 18F-FTO was similar to that of 18F-FTP at 30 min after injection (Table 1). At 2 h, myocardial uptake of 18F-FTO was maintained, whereas 18F-FTP uptake in the heart was 82% reduced. Hepatic clearance was faster for 18F-FTP than for 18F-FTO. Bone uptake for the 2 tracers was comparable at 120 min, suggesting that their defluorination rates in rats were equivalent.
Heart-to-organ ratios for 18F-FTO and 18F-FTP at 120 min after administration of tracers are shown in Figure 3. Heart-to-blood, heart-to-lung, and heart-to-liver ratios for 18F-FTO were 4-, 4.8-, and 3.4-fold higher, respectively, than for 18F-FTP.

Heart-to-organ ratios for 18F-FTO (open bars) and 18F-FTP (filled bars) in fasted rats at 2 h after administration of radiotracers (n = 6, both groups). All 18F-FTO values are statistically higher than 18F-FTP values (P < 0.05).
Folch-type analysis of tissue extracts showed that 18F-FTO is metabolized predominantly to protein-bound species in control hearts (Table 2), consistent with previous reports for 18F-FTP (14) and 18F-FTHA (16). At 30 min, the protein-bound fraction of radioactivity was significantly higher for 18F-FTO (∼90%) than for 18F-FTP (∼60%) (P < 0.05). Pretreatment of rats with etomoxir caused a dramatic shift from protein-bound species (>90%) to organic-soluble species (>65%) for 18F-FTO at 30 min (Fig. 4), demonstrating the dependence of the formation of protein-bound metabolites on CPT-I dependent FAO. At 120 min, protein-bound, aqueous, and organic fractions in the liver were similar for 18F-FTO and 18F-FTP, but 18F-FTO showed a 60% higher (P < 0.01) protein-bound fraction in the skeletal muscle than did 18F-FTP (Table 2). Both organic and aqueous fractions were higher for 18F-FTP than for 18F-FTO in the skeletal muscle. The balance of non–protein-bound radioactivity for 18F-FTO was split nearly evenly between aqueous and organic fractions as unidentified metabolites, whereas for 18F-FTP, a larger fraction was found in the organic fraction. Thus, the data suggested that metabolic incorporation of the oleate analog 18F-FTO into mitochondrial proteins of skeletal muscle appears to be similar to that shown in the heart (80–90%), whereas mitochondrial trapping of 18F-FTP represents approximately one half of its metabolic fate.
Distribution of 18F Radioactivity (% of Total) in Aqueous, Organic, and Pellet Fractions After Folch-Type Extraction of Tissues

Effect of CPT-I inhibitor etomoxir (40 mg/kg intraperitoneal pretreatment 2 h before) on 18F radioactivity distribution in Folch-type extractions of rat hearts analyzed at 30 min after intravenous administration of 18F-FTO (control group, n = 6; etomoxir group, n = 4). Rats were fasted overnight. Statistical significance of etomoxir-induced changes: *P < 0.02 vs. control.
Small-Animal PET Studies in Rats
A preliminary small-animal PET study was performed using 18F-FTO and 18F-FTP in separate groups of fasted rats (Fig. 5). Early dynamic images showed rapid blood clearance of both tracers to low radioactivity concentrations within 2 min after injection. Likewise, lung clearance was rapid, providing low background to the heart by 2 min after injection. The predominant hepatic uptake of the 2 fatty acid analogs peaked at 5 min after injection, followed by slow clearance. Myocardial uptake plateaued at about 10 min for both radiotracers, remaining nearly constant for 18F-FTO, whereas more pronounced myocardial clearance was observed for 18F-FTP. In the 2 rats studied, myocardial uptake of 18F-FTO, expressed as standardized uptake value, was higher than that for 18F-FTP. This observation could not be explained by higher arterial concentrations of 18F-FTO: integrations of the blood pool curves over 0–2 min, when metabolites should not have a significant impact, were essentially the same for 18F-FTO and 18F-FTP. Whole-body images acquired at 55–115 min after injection demonstrated superior myocardial imaging characteristics of 18F-FTO consistent with the biodistribution data (Fig. 5). Myocardial uptake relative to liver and bone was markedly lower for 18F-FTP. Bone uptake in the ribs was seen with both radiotracers, evidencing in vivo defluorination in the rat.

Early time–activity curves for 18F-FTO and 18F-FTP in rat heart (left ventricle), liver, lung, bone (rib), and blood pool and PET thoracic images acquired at 55–115 min after administration. Heart, bone, and liver uptake is evident on PET images. Higher heart-to-liver radioactivity concentration ratio was seen with 18F-FTO. Blood-pool data were not corrected for spillover effect from myocardium.
DISCUSSION
FAO is the major energy-providing process in the heart, liver, and skeletal muscle. Because disturbances of FAO are associated with many cardiovascular, gastrointestinal, oncologic, neurologic, and metabolic disorders, a noninvasive imaging method has great potential to affect the clinical care of patients. An imaging method would be of great value for monitoring disease progression and the effects of a growing number of drug therapies aimed at modifying FAO rates (23–25). The current study extends our previous work with thia-substituted fatty acid analogs (12,14,16) to develop an 18F-labeled fatty acid molecule that can serve as a metabolically trapped, general-purpose probe for PET of FAO in humans.
To date, nearly all the PET studies of fatty acid metabolism have been performed using 11C- or 18F-labeled saturated fatty acid analogs such as 11C-palmitate and 18F-labeled palmitate. The early studies of Knust et al. (26) showed that in mice 18F-fluoro fatty acids distribute in a manner similar to palmitate. As is the case for 11C-palmitate, the FAO rate must be derived from the clearance kinetics of the radiolabel, requiring various assumptions and complex tracer kinetic modeling approaches (27). The 11C-palmitate modeling method has not been validated in ischemic myocardium because of an elevated and variable level of backdiffusion of nonmetabolized radiotracer that confounds the clearance of labeled catabolite (27,28). Various branched-chain fatty acid analogs have been developed as metabolically retained probes (29,30), but they are poorly accepted for transport into the mitochondrion through CPT-I (31), rendering them sensitive for the indication of extramitochondrial lipid incorporation processes but not sensitive for the indication of FAO. Our approach of sulfur substitution (16) followed earlier work with larger heteroatom substituents (32). It was established that the sulfur substitution greatly diminishes the incorporation of fatty acids into complex lipids (33), rendering its kinetics more specific to mitochondrial metabolism. 18F-FTHA, a 6-thia analog, was shown to be sensitive to CPT-I inhibition (16), but later studies indicated FAO-independent retention of radiotracer in hypoxic myocardium (19). Later, the 4-thia analog 18F-FTP was found to correlate well with FAO rates in isolated rat hearts in normoxic and hypoxic conditions (12,14). However, both 18F-FTHA (16) and 18F-FTP (14) showed significant clearance from rodent myocardium between an early peak (<10 min) and 2 h after administration. Slow but appreciable myocardial clearance was also observed in humans with 18F-FTHA (34), although 18F-FTP was well retained in healthy swine myocardium (14).
This is the first report of a positron-emitter labeled oleate analog. The 4-thia oleate analog 18F-FTO was synthesized in practical yield by analogy with 18F-FTP. Because of the presence of the cis double bond, the synthesis of the labeling precursor of 18F-FTO involved multiple steps but was straightforward. Because both 18F-FTO and 18F-FTP share the 4-thia substitution, any differences in their biodistributions and biochemical handling can be attributed to differences in chain length and unsaturation. The initial myocardial uptake of 18F-FTO was similar to that of 18F-FTP, but myocardial retention was dramatically increased. Although there was no evidence of clearance of 18F-FTO from the rat myocardium out to 2 h after injection, there was approximately 80% clearance of 18F-FTP from 30 to 120 min. The reasons for the higher myocardial retention of 18F-FTO are not clear. There was evidence in the Folch-type tissue-extraction measurements that 18F-FTO had a higher specificity to indicate mitochondrial FAO in the heart than did 18F-FTP. The protein-bound fraction of 18F radioactivity of 4-thia (18) and 6-thia (20) fatty acid analogs indicates the accumulation of FAO-dependent metabolites in the tissue, as supported by profound decreases of this fraction with CPT-I inhibition. This fraction decreased from 80% to 45% for 18F-FTP in rat heart (30 min after injection) under control and CPT-I–inhibited conditions, respectively (18). In contrast, the protein-bound fraction in the rat heart for 18F-FTO decreased from 90% to 25% under control and CPT-I–inhibited conditions, respectively (Fig. 4). However, decreases in myocardial uptake caused by CPT-I inhibition (∼80%) at 30 min after injection were found to be equivalent for 18F-FTO and 18F-FTP (18). Both 18F-FTO and 18F-FTP appear to be highly specific probes for the indication of FAO in the heart. However, the slower myocardial clearance and 3- to 4-fold higher heart-to-blood, heart-to-lung, and heart-to-liver radioactivity concentration ratios favor 18F-FTO over 18F-FTP for heart imaging applications.
With regard to skeletal muscle, our measurements at 120 min after injection showed 18F-FTO to have a significantly higher protein-bound fraction relative to 18F-FTP (Table 2), indicating that 18F-FTO may have better utility for the evaluation of FAO in skeletal muscle. Previous work with 18F-FTHA in pigs had shown the 6-thia analog to be poorly taken up into skeletal muscle mitochondria (35). Taken together with the results of the present study, the approximate distribution of 18F-FTHA, 18F-FTP, and 18F-FTO to the protein-bound mitochondrial fraction of control skeletal muscle is 36%, 50%, and 80%, respectively. Thus, the results indicate that 18F-FTO kinetics are sufficiently specific in skeletal muscle to warrant further evaluation of 18F-FTO as an FAO probe for monitoring of FAO in metabolic disorders that involve skeletal muscle.
The suggested enhancement of specificity of 18F-FTO relative to 18F-FTP and 18F-FTHA for the indication of FAO in skeletal muscle has some support from previous studies of oleate and palmitate metabolism. A whole-body metabolism study in humans showed that dietary 13C-oleate was preferentially oxidized relative to dietary 13C-palmitate (22). Because skeletal muscle is the major consumer of fatty acids in the body, this study implied that skeletal muscle preferentially oxidized oleate relative to palmitate. Likewise, a human feeding study in which the ratio of polyunsaturated to saturated fatty acids in the diet was altered also suggested that polyunsaturated fatty acids are more highly oxidized than are saturated fatty acids (36).
Heart and skeletal muscle uptake of 18F-FTO was found to be the same in fed and fasted rats. This does not imply equivalent FAO rates in heart and muscle in the different feeding conditions. Radiotracer uptake is a measure of the fraction of dose taken up per gram of tissue. Taking into consideration that plasma concentrations of nonesterified fatty acids are 2.2-fold higher in the fasted state than in the fed state in the same experimental conditions (14), equivalent uptake is consistent with higher FAO rates in fasted animals in heart and skeletal muscle.
The liver is a predominant organ of uptake for 18F-FTO, consistent with previously developed fatty acid radiotracers. Folch-type extraction analysis of livers from rats administered 18F-FTO showed that 70.9 ± 2.9% of the trapped 18F radioactivity was in the form of protein-bound metabolites at 2 h after injection, indicating that 18F-FTO may be useful for the evaluation of hepatic FAO rates. In comparison, 18F-FTHA showed 57.5 ± 1.9% protein-bound metabolites at 10 min after injection (16), and 18F-FTP was in between (63.3 ± 9.3% at 2 h after injection). Notably, the liver uptake of 18F-FTO was significantly lower in fed rats than in fasted rats (0.836 ± 0.086% vs. 1.185 ± 0.155% dose kg/g at 30 min after injection), consistent with lower hepatic FAO rates in the fed state. 18F-FTP lacked this sensitivity (1.24 ± 0.44% vs. 1.15 ± 0.16% dose kg/g) (14). The lack of a response of liver uptake of 18F-FTO to the CPT-I inhibitor etomoxir at 30 min after injection likely reflects the presence of uptake and oxidation mechanisms that are CPT-I–independent in the liver, such as peroxisomal FAO. Etomoxir was found to paradoxically stimulate palmitate oxidation in rat liver homogenates through a peroxisome-dependent pathway (37). The PET-measured hepatic concentrations of the thia fatty acid analogs are relevant to hepatic fatty acid uptake and oxidation, with 18F-FTO being the most specific to date for indicating FAO.
The in vivo defluorination of ω-labeled, non-β-oxidizable 18F-fluoro-fatty acids is known to be problematic in rodents (38) but has not been observed to be a problem in pigs or humans (14). Because the synthesis of labeling precursor is more straightforward for labeling at the ω-position, we developed 18F-FTO as the first oleate analog. Should defluorination of 18F-FTO be found in subsequent studies in large animals and humans, an ω-3–labeled analog will be developed to stabilize the 18F radiolabel (38).
CONCLUSION
A practical and high-purity synthesis of the oleate analog 18F-FTO was developed. 18F-FTO exhibited 3- to 4-fold higher heart-to-background tissue radioactivity concentration ratios, resulting from higher myocardial retention relative to 18F-FTP. 18F-FTO uptake by the heart was approximately 80% reduced by pretreatment of rats with the CPT-I inhibitor etomoxir. Folch-type extraction analyses showed 70–90% protein-bound fractions in the heart, liver, and skeletal muscle, consistent with the high trafficking of 18F-FTO to the mitochondrion with subsequent metabolism to protein-bound species. Small-animal PET images of 18F-FTO accumulation in the rat myocardium were clearly superior to those of 18F-FTP. 18F-FTO has been shown to be a promising metabolically trapped FAO probe that improves on the myocardial imaging characteristics of 18F-FTP and warrants further evaluation.
Acknowledgments
We thank Aditya Bansal for his assistance. We acknowledge the support of the NIH (grants R01 HL-63371 and R01 CA108620). Dr. Timothy DeGrado is a shareholder of Tracera, LLC.
- © 2010 by Society of Nuclear Medicine
REFERENCES
- Received for publication December 22, 2009.
- Accepted for publication April 8, 2010.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}