


Abstract
Noninvasive molecular imaging of dynamic processes has benefited tremendously from the use of reporter genes. These genes encode for proteins that emit light, bind radiolabeled probes, or, as covered in this review, modulate MRI contrast. Reporter genes play a pivotal role in monitoring cell trafficking, gene replacement therapy, protein–protein interactions, neuronal plasticity, and embryonic development. Several strategies exist for generating MRI contrast: using enzyme-catalyzed chemical modification of metal-based contrast agents or (phosphorus) metabolites, iron-binding and iron-storage proteins to accumulate iron as a contrast agent, and artificial proteins for imaging based on chemical exchange saturation transfer. MRI reporter genes have the advantage that the specific signal can be coregistered with soft-tissue anatomy and functional tissue information and have, therefore, become an active and growing area of scientific interest.
With more than 20,000 genes in the human genome now identified and a similar number of genes in the rat and mouse genome known (1), explanation of the function of genes has become the major challenge. Clarification of the function of genes has been facilitated by the development of reporter genes, which are revolutionizing the way many current biomedical studies are designed and performed. A reporter gene is a gene whose product can be readily detected and either be fused to the gene of interest or can replace it. The main applications for these reporters include monitoring gene expression levels, investigating dynamic molecular interactions between proteins, studying cellular interactions, tracking cells in normal and abnormal development or in cell transplantation therapy, and monitoring gene replacement therapy.
Optical reporter genes are probably the most commonly used for imaging and widely developed. Throughout the years, multiple genes were cloned from a variety of organisms that emit light via bioluminescence or fluorescence in multiple distinguishable wavelengths. Probably the most recent and exciting implementation of optical reporter genes was the development of the transgenic “brainbow” mouse, in which combinatorial expression of fluorescent proteins in the mouse brain resulted in the production of more than 89 distinguishable colors (2). An emerging new class of reporter genes encodes for proteins with affinity for radioisotopes or positron emitter probes. These receptors, transporters, or enzymes can provide quantitative images on administration of suitable radiolabeled probes (3). MRI reporter genes are unique among all reporter genes used with the various imaging modalities because they can provide information on gene expression that can be coregistered with anatomic and functional information (4). MRI reporters enable serial temporal imaging within the same subject. This is particularly useful for studying dynamic processes, for example, migration of stem cells and progenitors (5), neuronal plasticity, mechanisms of development and adaptation, disease progression and response to trauma or illness, and processes of memory and learning.
The purpose of this review was to give a basic, brief overview of the recent developments in the field of MRI reporter genes and to evaluate and compare the currently existing genes (Table 1) while suggesting future directions. A more detailed review can be found in Gilad et al. (4) and Modo et al. (6). For further reading, the context of MRI reporter genes is closely linked to certain MRI contrast agents that can act as sensors for changes in biologic or physiologic conditions (7).
Comparison of Existing MR Reporter Genes
BASIC PRINCIPLES OF MRI CONTRAST
MRI relies on measuring magnetization of magnetic nuclei subjected to radiofrequency irradiation inside a magnetic field. A variety of nuclei can be studied, with the most common one being the hydrogen nucleus (proton, 1H). For example, MRI detects properties of the water signal, mostly in terms of amount of water (so-called proton or spin density), its relaxation back to equilibrium after excitation (time constant, T1), and its signal broadening (relaxation times, T2 for natural signal width, and T2*, which includes T2 effects plus magnetic field inhomogeneity contributions). Any tissue difference in terms of these properties provides contrast. Also, besides water, MR spectroscopy (MRS) can detect individual proton signals in metabolites and nuclei different from protons, such as 31P, 13C, 23Na, 19F, and many more, all of which have specific frequencies. The signal acquired from different nuclei depends on their abundance in the tissue and their chemical interaction with other molecules, allowing multiple manipulations with various MRI acquisition schemes to create different contrasts. When certain metals or organic compounds are present, endogenous contrast mechanisms may be affected. T1 relaxation can be enhanced using compounds containing paramagnetic lanthanides, with gadolinium being widely used in the clinic. Iron oxide particles, on the other hand, can drastically shorten T2 and T2*, which is manifested by darkening of the MR images. As a consequence, many iron-binding proteins are potential candidates for MRI reporter genes.
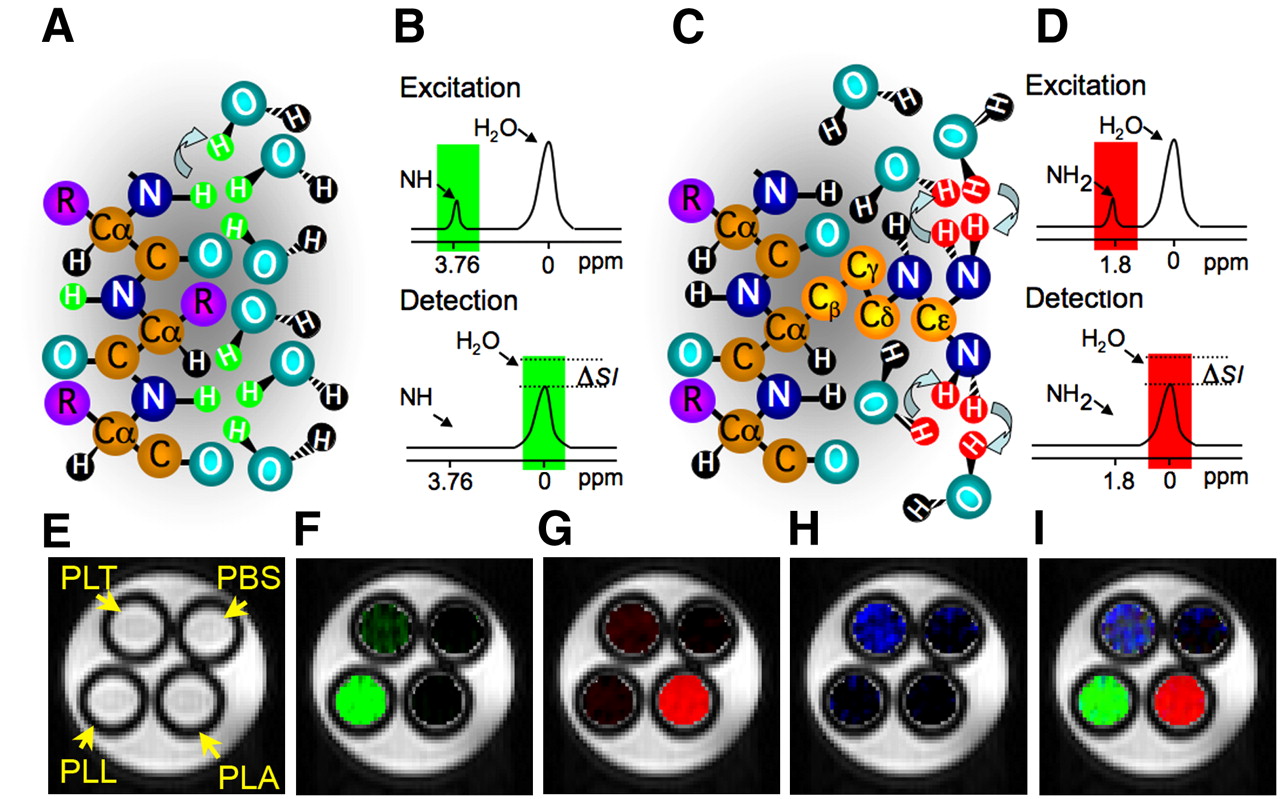
Chemical exchange saturation transfer (CEST) is a new type of MRI contrast that has been developed; it relies on the chemical exchange of protons of solutes such as contrast agents with bulk water (8). A variety of organic and organometallic compounds have a sufficient number of protons with suitable chemical exchange rates and specific MRI frequencies to be excited selectively and detected sensitively. A radiofrequency pulse, called a saturation pulse, is applied at the exchangeable proton resonance frequency. This saturation, or signal loss, is then transferred via exchange to bulk water, producing a fractional reduction in the water signal (Figs. 1A–1D).
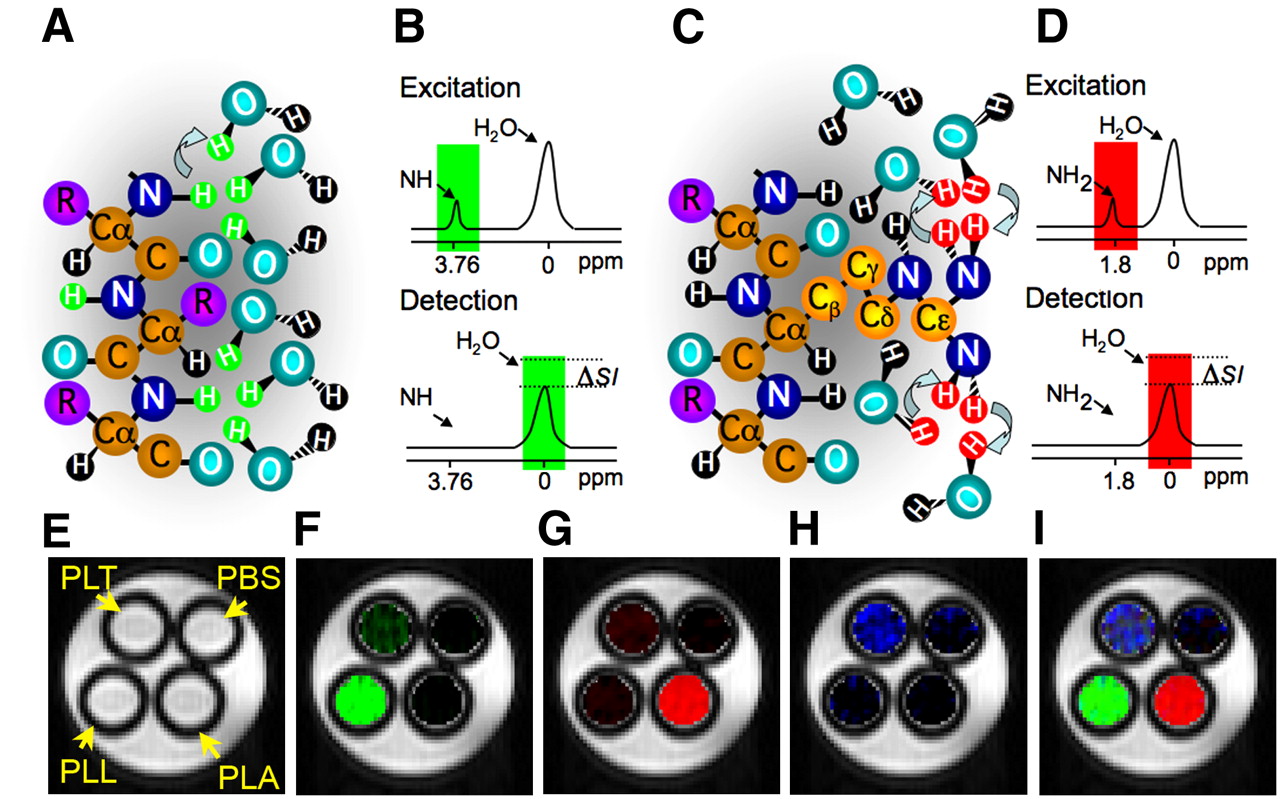
Principles of CEST contrast mechanism. Frequency-selective saturation pulse is applied to label amide protons (A, green) or guanidyl protons (C, red) of polypeptide contrast agent. Labeled protons exchange with water protons, which leads to reduction in MRI signal intensity (ΔSI) in frequency-selective manner (B and D). CEST maps of MRI phantoms were acquired with frequency-selective saturation pulses (E) at ±3.7 ppm, which enhances PLL (F); at ±1.8 ppm, which enhances PLA (G); and at ±0.8 ppm, which enhances PLT (H). Merged composition image of maps (F–H) is shown in I. PBS = phosphate-buffered saline. (Modified from McMahon et al. (23) and Gilad et al. (24)).
MRS: ENZYME-BASED REPORTER GENES
Historically, the first MRI reporter gene was creatine kinase (CK), an enzyme that catalyzes adenosine triphosphate conversion to adenosine diphosphate–producing phosphocreatine (PCr) and can be detected by 31P MRS. The first demonstration of CK was in its transgenic overexpression in the liver (9), in which high levels of PCr were observed; PCr, however, is absent in the livers of control mice. CK was subsequently used as a marker of low-density lipoprotein receptor (LDLr) expression in LDLr-deficient mice after treatment with an adenovirus engineered to coexpress recombinant human LDLr and the CK-B reporter gene (10). In a recent study, MRS was used to examine different promoters in yeast cells. Genes that catalyze the formation of polyphosphate were cloned and placed after different promoters, after which polyphosphate levels could be monitored with 31P MRI (11).
MRI: ENZYME-BASED REPORTER GENES
The overexpression of human tyrosinase initiates a chain of reactions including melanin production, which is followed by higher metal binding that may result in enhanced MRI signal intensity. A tetracycline-inducible system of tyrosinase has been developed in MCF-7 breast cancer cells (12) using this concept.
Another enzyme that was exploited as an MRI reporter gene is β-galactosidase (β-gal). β-gal is an enzyme encoded by the LacZ gene of Escherichia coli; it catalyzes hydrolysis of β-d-galactosides and is widely used in microscopy because of its large variety of substrates. The use of β-gal in MRI was demonstrated by its effect on (1-(2-(β-galactopyranosyloxy)propyl)-4,7,10-tris(carboxymethyl)-1,4,7,10-tetraazacyclododecane)gadolinium(III) (EgadMe) (13). Once EgadMe was cleaved by β-gal and its free coordination site of gadolinium was exposed, it altered the relaxation time of water and created contrast. β-gal was also used in spectroscopic imaging (14).
MRI: IRON-BASED REPORTER GENES
Another possibility for creating MRI reporter genes is the expression of iron-related proteins. Iron, depending on its oxidation state and its structure, can act as a paramagnetic reactive metal, which is essential for body homeostasis. The manipulation of iron concentration is expected to yield a detectable change in MRI contrast, which will be able to produce a real-time report on cellular events. Therefore, several iron-related proteins were tested as possible MRI reporters.
In the bloodstream, iron is bound to the plasma protein transferrin and enters cells via interaction of the iron–transferrin complex with the transferrin receptor. Administration of transferrin linked to monocrystalline iron-oxide nanocompounds to nude mice that had hTfR-transfected 9-L gliosarcoma tumors led to a significant difference in MRI signal 24 h after administration (15). The transferrin receptor has also been used for monitoring therapeutic gene expression when expressed as part of a vector that carried several genes, among them a prodrug therapy gene. All 3 transgenes were expressed in the same cell simultaneously, and MRI demonstrated the ability to use transferrin receptor as a reporter for gene therapy (16).
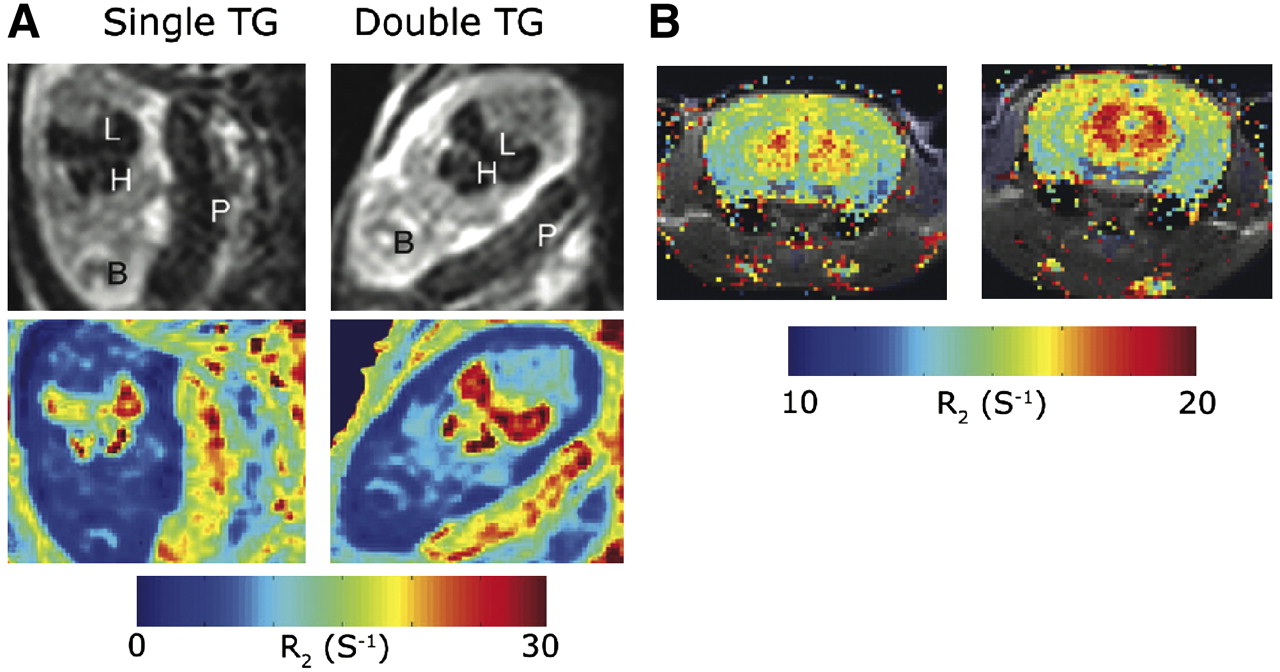
Although iron has a crucial role in body homeostasis, excess of iron can cause damage to cells by catalyzing the Fenton reaction, which generates reactive oxygen species. Therefore, excess intracellular iron is stored in the ferrihydrite core of the ferritin protein. The first use of ferritin in the design of C6 glioma MRI reporter cells was done with a construct that carried enhanced green fluorescent protein and the heavy chain of ferritin, which exhibits ferroxidase activity, under tetracycline control. Inoculation of these tumor cells in nude mice revealed a significant decrease in T2 on tetracycline withdrawal (17). Transgenic mice that expressed the heavy chain of ferritin in a tissue-specific and tetracycline-dependent manner revealed a significant change in T2 values that was tissue-dependent: decreased T2 when expressed exclusively by endothelial cells as expected (Fig. 2) and surprisingly increased T2 when expressed by liver hepatocytes. The reason for the opposite direction of T2 changes in the liver was found to be related to the overexpression of H-ferritin in the hepatocytes, which induced mild toxicity and resulted in water vacuole formation. These water vacuoles contributed to the elevation in T2 values due to the long T2 of free water (18).

MRI detection of endothelial overexpression of H-ferritin in transgenic mice. Expression of ferritin was regulated by vascular endothelial cadherin promoter in double transgenic mice but not in single transgenic siblings. Elevated R2 was observed in liver and heart of E13.5 embryos studied in utero (A) and in brain of adult mice (B). TG = transgenic. (Adapted from Cohen et al. (18)).
Genove et al. demonstrated the use of ferritin as an MRI reporter gene by injecting an adenovirus that encoded for human ferritin into the brain parenchyma of mice, resulting in significant loss of signal at the site of inoculation (19). Coexpression of ferritin and the transferrin receptor showed signal loss on T2- and T2*-weighted MR images of neuronal stem cells in an iron-enriched environment (20). Further improvements in ferritin MRI reporter gene sensitivity may be possible. For instance, Bennett et al. proposed that aggregation of ferritin could augment its efficacy. Chemical cross-linking of ferritin to actin in vitro led to polymerization, which resulted in a 20% decrease in T2 (21).
Recently, Mag A, a bacterial iron transporter in Aquaspirillum magnetotacticum from the magnetotactic bacteria family, was expressed in 293FT cells under doxycycline regulation. Administration of doxycycline and iron resulted in significantly shorter T2, and inoculation of these cells to a mouse brain induced significant signal loss on T2*-weighted images (22).
MRI: CEST-BASED REPORTER GENES
CEST contrast agents have 2 major advantages. First, they are switchable; that is, the contrast is detectable only when a saturation pulse is applied at the specific frequency characteristic of the exchangeable protons of an agent. Otherwise, the contrast agent is MRI-invisible. The second advantage is that different contrast agents with different excitation frequencies can be used for imaging simultaneously more than one target. This property might be exploited for imaging multiple cells (e.g., for studying neuronal circuits) or the expression of multiple genes (such as in genetic circuits). This so-called multicolor CEST MRI concept is shown in Figures 1E–1H for the polypeptides poly-l-lysine (PLL), poly-l-arginine (PLA), and poly-l-threonine (PLT) (23). Although PLL is detectable through its exchangeable amide protons, PLA provides strong CEST contrast through the exchangeable guanidyl protons and PLT has exchangeable hydroxyl protons, with a unique frequency. Although each of the polypeptides provides some CEST contrast in the frequency range of other polypeptides, it is relatively easy to separate the signal derived from each polypeptide with proper image processing. Therefore, the 3 different polypeptides are clearly distinguishable with MRI, making them suitable candidates for labeling different targets.
As a first demonstration, a lysine-rich protein (LRP), containing a high density of amide protons, was cloned and expressed in 9-L rat glioma cells. The LRP amino acid sequence is similar to PLL and provides MRI contrast at the amide proton frequency. LRP overexpression was detectable in vitro in cell extracts and in vivo in xenografts of LRP-expressing 9-L cells in the mouse brain (24).
FUTURE PROSPECTS
MRI reporter genes are still in their infancy and need to be further improved to facilitate wider use. A major improvement in sensitivity for both ferritin and CEST reporter genes is likely to occur with MRI scanners moving toward ever higher fields (up to 9.4 T for humans and as high as 18.8 T for scanners suitable for animal research). In addition, the acquisition of more specific reporter-induced MRI contrast may be achieved using optimized pulse sequences and improved ways of image processing (analogous to those used for detecting small changes in blood oxygen level–dependent functional MRI). From the molecular biology side, reporter genes may be improved through directed evolution in a fashion similar to that accomplished for the fluorescent reporter genes (25). A key challenge would be to resolve multiple targets while performing simultaneous imaging. This could include different reporters driven by different promoters or developing sophisticated data-processing algorithms to distinguish gene expression temporally as well as spatially.
From the clinical point of view, a few significant hurdles still remain before reporter genes can be introduced into the human body. Once this has been achieved in a safe manner, we can foresee 2 major clinical applications. The first one will be tracking genetically labeled immunotherapeutic or regenerative stem cells for monitoring cell survival, migration or homing, and differentiation. The second application will be monitoring gene replacement therapy; here, the MRI reporter genes might become the method of choice to monitor successfully induced therapeutic gene expression long before a phenotypic readout becomes available.
Acknowledgments
Drs. Gilad and Ziv contributed equally to this work. The studies described here were supported, in part, by grant KO1 EB006394 and grants from the Israel Science Foundation, the Minerva Foundation, and NIH Roadmap (R21 EB005252).
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication June 20, 2008.
- Accepted for publication September 15, 2008.





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A dual-gene reporter-amplifier architecture for enhancing the sensitivity of molecular MRI by water exchange
- Engineering water exchange is a safe and effective method for magnetic resonance imaging in diverse cell types
- Development of a Synthetic Biosensor for Chemical Exchange MRI Utilizing In Silico Optimized Peptides
- Imaging cellular immunotherapies and immune cell biomarkers: from preclinical studies to patients
- Molecular Imaging with Reporter Genes: Has Its Promise Been Delivered?
- A Genetically Encoded Reporter for Diffusion Weighted Magnetic Resonance Imaging
- Advances in using MRI probes and sensors for in vivo cell tracking as applied to regenerative medicine
- Gene Therapy Review
- In Vivo Tracking Techniques for Cellular Regeneration, Replacement, and Redirection
- Imaging Gene Expression in Live Cells and Tissues
- Recording Intracellular Molecular Events from the Outside: Glycosylphosphatidylinositol-Anchored Avidin as a Reporter Protein for In Vivo Imaging
- MR-Based Molecular Imaging of the Brain: The Next Frontier