


Abstract
Prader–Willi syndrome (PWS) is a genetic disorder caused by the nonexpression of paternal genes in the PWS region of chromosome 15q11-13 and is the most common cause of human syndromic obesity. Methods: We investigated regional brain metabolic impairment in children with PWS by 18F-FDG PET. Sixteen children with PWS (9 males, 7 females; mean age ± SD, 4.2 ± 1.1 y) and 7 healthy children (4 males, 3 females; mean age ± SD, 4.0 ± 1.7 y) underwent brain 18F-FDG PET in the resting state. The images of PWS children were compared using statistical parametric mapping analysis with those of healthy children in a voxelwise manner. Results: Group comparison showed that children with PWS had decreased glucose metabolism in the right superior temporal gyrus and left cerebellar vermis, regions that are associated with taste perception/food reward and cognitive and emotional function, respectively. Metabolism was increased in the right orbitofrontal, bilateral middle frontal, right inferior frontal, left superior frontal, and bilateral anterior cingulate gyri, right temporal pole, and left uncus, regions that are involved in cognitive functions related to eating or obsessive–compulsive behavior. Interestingly, no significant metabolic abnormality was found in the hypothalamus, the brain region believed to be most involved in energy intake and expenditure. Conclusion: This study describes the neural substrate underlying the abnormal eating behavior and psychobehavioral problems of PWS.
Prader–Willi syndrome (PWS), the most common cause of syndromic obesity, is caused by the deletion or disruption of a gene or several genes on the proximal long arm of paternal chromosome 15, maternal uniparental disomy, or imprinting defect. These genetic defects occur because the genes on maternal chromosome 15q11-13 are virtually inactive through imprinting (1), which leads to the lack of expression of parental copies of the imprinted SNRPN gene, the necdin gene, and possibly other genes.
Clinically, the syndrome is characterized by various symptoms, including infantile hypotonia and poor sucking reflex, mental retardation, delayed sexual development, and obesity by incessant food intake. Among these symptoms, food-related difficulties are the most striking and widely recognized marker of this syndrome.
In addition to these well-known problems, those with PWS also suffer from a range of psychiatric and behavioral problems—for example, high rates of depression, obsession, and compulsion (2–4). Cognitive deficits such as impaired social cognition, literal mindedness, and cognitive inflexibility are also present in PWS. Neuropsychological studies have demonstrated deficits in attention, working memory, and sequential information processing in PWS patients (5–7), which may be related to frontal lobe dysfunction.
Although morphologic abnormalities in brain imaging are not cardinal features of PWS, several studies have attempted to identify structural problems in the PWS brain (8–10). One study compared anterior pituitary gland sizes, an important site of growth hormone secretion, in PWS and normal control subjects using MRI, but no statistically significant differences were found (8). Another MRI study, which included 5 PWS patients, described subtle morphologic abnormalities, including mild ventriculomegaly, cortical atrophy in the frontal lobe, and a small brain stem (9). Yoshii et al. reported a case of cortical malformation in a 40-wk-old girl with PWS (10). MR images showed a diffusely abnormal gyral folding pattern and multiple regions involving all lobes where the gray–white matter junction was markedly irregular. However, morphologic studies have not provided an insight into the neural substrates underlying the neuropsychiatric, behavioral, and cognitive deficits of PWS.
In an attempt to identify functionally impaired brain regions in PWS and to correlate functional deficits with cognitive and behavioral symptoms, we investigated regional brain metabolic abnormalities in PWS children using 18F-FDG PET.
MATERIALS AND METHODS
Participants
Sixteen children with PWS (9 males, 7 females; mean age ± SD, 4.2 ± 1.1 y) and 7 age- and sex-matched healthy children (4 males, 3 females; mean age ± SD, 4.0 ± 1.7 y) were enrolled in this study.
PWS patients were genetically confirmed using the standard methylation test. Of these 16 patients, 14 had deletion of the paternally transmitted chromosome 15q11-13, whereas the other 2 patients had uniparental disomy. All control subjects were brothers, sisters, or relatives of the patients. They did not have the clinical symptoms of PWS and were negatively confirmed by genetic testing. All control subjects were healthy without a history of seizure disorder. We explained the purpose and methods of this study and obtained informed consent from the parents of the PWS patients and control subjects. The study design was reviewed and approved by the Samsung Medical Center Institutional Review Board. The PWS patients and control subjects had no history of growth hormone or sex hormone treatment and were not being treated with either hormone at study commencement. Clinical characteristics of the PWS patients are summarized in Table 1.
Clinical Characteristics of PWS Patients and Control Subjects
18F-FDG PET Procedures
PET scans of 10 min were acquired in 3-dimensional (3D) mode using an Advance PET scanner (GE Healthcare), starting 40 min after the intravenous injection of 4.8 MBq/kg 18F-FDG. In-plane and axial resolutions of the scanner were 4.9-mm and 3.9-mm full width at half maximum (FWHM), respectively. Subjects were fasted for at least 4 h before PET. Thirty minutes before PET scans, study subjects were orally sedated with chloral hydrate (50 mg/kg). PET images were reconstructed using a Hanning filter (cutoff frequency = 4.5 mm) and displayed in a 128 × 128 matrix (pixel size = 1.95 × 1.95 mm with a slice thickness of 4.25 mm). Attenuation correction was performed with a uniform attenuation coefficient (μ = 0.096 cm−1).
Statistical Parametric Mapping (SPM) Analysis of Regional Glucose Metabolism
Before statistical analysis, using SPM99 (Wellcome Department of Cognitive Neurology, Institute of Neurology, London, U.K.) (11) implanted in the Matlab 5.3 environment (MathWorks Inc.), all images were preprocessed for spatial normalization into the MNI (Montreal Neurologic Institute) template to remove intersubject anatomic variabilities, then smoothed with a FWHM 16-mm gaussian kernel to increase the signal-to-noise ratio and to remove subtle variations in anatomic structures. Individual voxel counts were normalized versus total brain counts (proportional scaling in SPM) to remove individual global brain metabolism differences. After spatial and count normalization, significant differences in glucose metabolism between PWS and healthy children were estimated at every voxel using t statistics. Abnormal glucose metabolism was investigated in brain areas at a height threshold of uncorrected P = 0.001 and at an extent threshold of 70 voxels for between-group analysis and at a height threshold of uncorrected P = 0.01 and an extent threshold of 70 voxels for individual-to-group analyses, after adjusting for the age factor as a covariate of no interest. To visualize the t-score statistics (SPM{t} map), significant voxels were projected onto the 3D rendered brain and a standard high-resolution MRI template provided by SPM99, thus allowing anatomic identification. The MNI coordinates of the local maximum of each cluster were converted into Talairach–Tournoux coordinates (12).
RESULTS
Figure 1 and Table 2 show brain regions with abnormal glucose metabolism in the PWS group versus the control group. Significant hypometabolism was observed in the right superior temporal gyrus and left cerebellar vermis. Metabolism was significantly elevated in the right orbitofrontal, bilateral middle frontal, right inferior frontal, and left superior frontal gyri. In addition, significant hypermetabolism was found in the bilateral anterior cingulate gyri, the right temporal pole, and the left uncus. Interestingly, no significant metabolic abnormality was found in the hypothalamus, the brain region believed to be most involved in energy intake and energy expenditure.
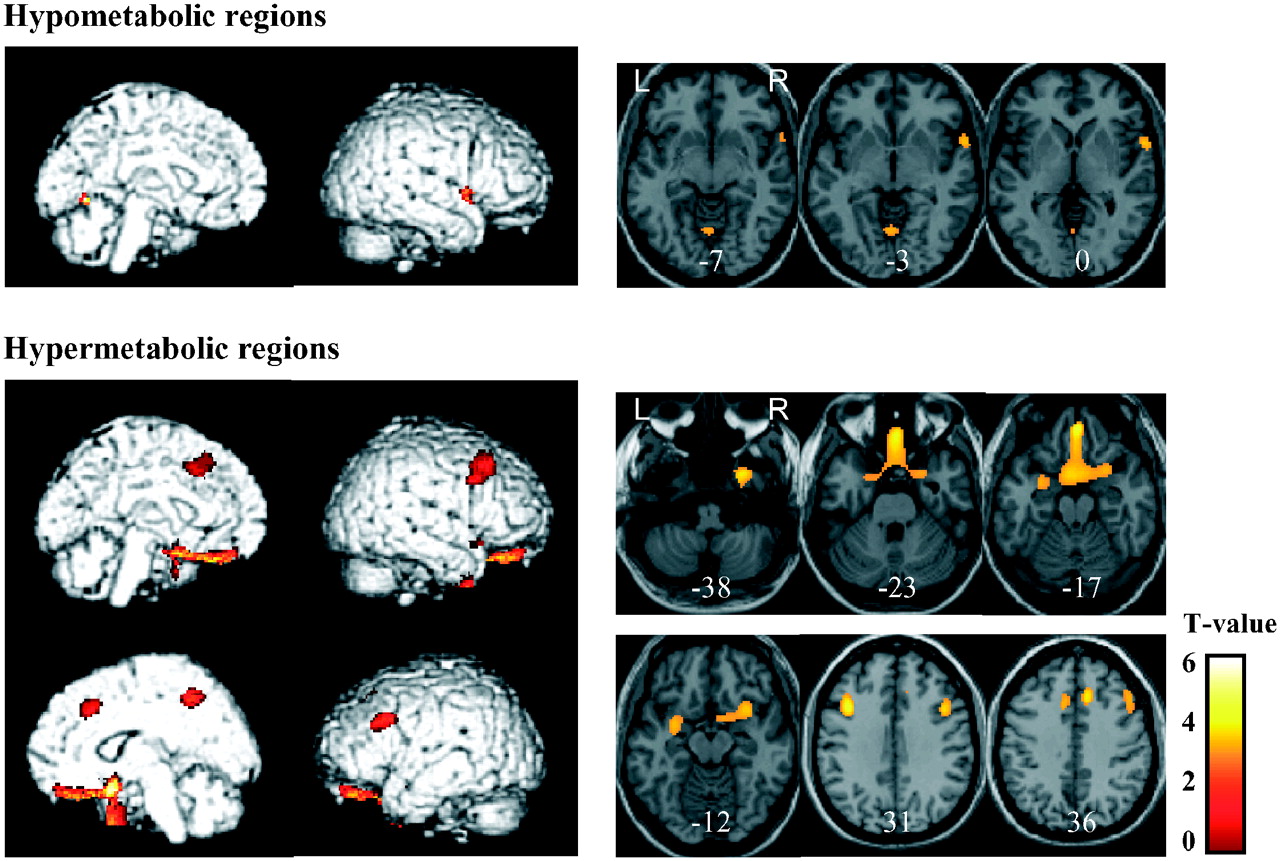
Hypo- and hypermetabolic regions in PWS patients vs. healthy control subjects. These regions are displayed on surface rendered and transaxial images. Brain regions of abnormal glucose metabolism are displayed using height threshold of uncorrected P = 0.001 and extent threshold of 70 voxels, after removing age as a covariate of no interest. Numbers in each transaxial brain image indicate distance (mm) from the anterior commissure–posterior commissure plane.
Hypo- and Hypermetabolic Regions in PWS Patient Group
Results of individual analysis are shown in Table 3, which confirm the group analysis findings. Hypometabolism in the superior temporal gyrus was found in 9 of the 16 patients. Most of the patients had hypermetabolism in the orbitofrontal (12/16), middle frontal (16/16), inferior frontal (6/16), superior frontal (14/16), and anterior cingulate (9/16) gyri and the uncus (10/16).
Hypo- and Hypermetabolic Regions in Individual PWS Patients
DISCUSSION
To our knowledge, the present study is the first functional brain mapping study in children with PWS. Interestingly, the metabolism of several brain areas was found to differ significantly in PWS and age- and sex-matched healthy children. PWS is known to be associated with obesity, but infantile PWS patients are rather weak and poorly nourished. The transition from a poorly nourished to an overweight state usually occurs before 6 y of age. In this study, we included PWS patients at the transitional period to see whether any brain metabolic changes were present at this period.
We attempted to interpret regional metabolic abnormalities versus abnormal eating behavior and psychobehavioral problems in PWS. We found that PWS children have reduced metabolism in the right superior temporal gyrus and left cerebellar vermis, regions that are associated with taste perception/food reward and cognitive and emotional function, respectively. The superior temporal region has been associated with liquid meal taste perception in the unpleasant state of extreme hunger (13). In addition, the superior temporal sulcus and the temporal pole have reciprocal connections with the insula (14), which, as a limbic/paralimbic area, constitutes a part of the taste pathway with the thalamus and hypothalamus, and which is involved in monitoring negative sensory input (15). It has also been suggested that the insular cortex is involved in incentive learning for food rewards (16). Moreover, cognitive and emotional problems may accompany cerebellar abnormalities or be their principal clinical presentation. Pathology in the cerebellar vermis (limbic cerebellum) and lateral hemisphere of the posterior cerebellum (involved in cognitive processing) is associated with cerebellar cognitive affective syndrome, which includes impairments in executive, visual-spatial, and linguistic abilities, with affective disturbances ranging from emotional blunting and depression to disinhibition and psychotic features (17).
In the present study, significant hypermetabolism was found in the right orbitofrontal, bilateral middle frontal, right inferior frontal, left superior frontal, and bilateral anterior cingulate gyri, right temporal pole, and left uncus in PWS children. The pre- and middle frontal and anterior cingulate regions are related to food intake in humans, and the frontal lobe is involved in cognitive functions related to eating; moreover, a feeding-suppression mechanism has been suggested to be located in the frontal lobe (18). Supporting this, frontal regions have been postulated to exert suppressing effects on other cerebral regions, such as the hypothalamus, which respond to food-related stimuli (19). The orbitofrontal cortex is a main cerebral area related to motivation and reward and is especially sensitive to food and monetary reward (20). Studies in nonhuman primates have shown that the orbitofrontal cortex is a cerebral region where responses to visual representations of food are modulated by an animal's hunger status (21). In humans, there was increased activation to visual food stimuli in the medial frontal/orbitofrontal cortex, amygdala, and insula in the premeal condition, which resolved after ingestion of a small meal (22,23). Frontal regions have also been shown to be associated with hunger and satiation in healthy humans—that is, hunger with increased blood flow in the posterior orbitofrontal cortex and satiety with increased blood flow in the ventromedial and dorsolateral prefrontal cortices (24). In obsessive–compulsive disorder, PET studies have identified hypermetabolism in the orbitofrontal cortex, caudate nucleus, and anterior cingulate cortex (25–27). In addition, a volumetric study found that patients with obsessive–compulsive disorder have more total gray matter in the anterior cingulate gyrus (28) Orbitofrontal and anterior cingulate hypermetabolism, which is the most consistent finding in obsessive–compulsive disorder, and which was observed in our PWS patients, may constitute a neural correlate of obsessive–compulsive symptoms in PWS (29–31). Studies in monkeys have revealed a role for the temporal pole in a variety of functions, including taste and olfaction. The uncus is a bulbar allocortical protrusion located in the anterior–inferior medial part of the temporal lobe and receives a projection from the lateral striae of the olfactory tract. Gustatory and olfactory hallucination or auras have long been thought to originate from lesions in this area (32,33). In summary, our results indicate that PWS children have decreased glucose metabolism in the right superior temporal gyrus and left cerebellar vermis, which are associated with taste perception/food reward or cognitive and emotional function, respectively, and increased metabolism in the right orbitofrontal, bilateral middle frontal, right inferior frontal, left superior frontal, and bilateral anterior cingulate gyri, right temporal pole, and left uncus, regions involved in cognitive functions related to eating or obsessive–compulsive behavior.
Recent functional brain imaging studies demonstrated abnormal brain responses after food intake in adults with PWS (34). This study found that the satiety response in the orbitofrontal cortex is insensitive to food intake in PWS adults. Shapira et al. reported a delay in activation after glucose administration in the hypothalamus and other brain regions associated with satiety, such as insula, ventromedial prefrontal cortex, and nucleus accumbens in adults with PWS (35). Interestingly, we found no significant metabolic abnormality in the hypothalamus, which has a major role in the control of food intake. However, neurotracing studies have shown that the hypothalamus receives input from several other regions of the brain (e.g., the amygdala, hippocampal complex, insula, and several polymodal association areas of the prefrontal cortex) that are likely to modulate its activity (36,37). Therefore, the hypothalamus is likely to be only one of several brain centers involved in the regulation of eating behavior, all of which integrate central and peripheral signals before, in turn, transmitting impulses to sets of as yet-unidentified efferent sites. In this context, metabolic abnormalities in the prefrontal and limbic/paralimbic areas, as revealed in the present study, may represent a neural correlate of abnormal food intake in PWS.
This study has some limitations. First, a major limitation of this study is that study subjects were sedated with chloral hydrate before PET. Sedatives are known to lower cerebral glucose metabolism. Although the exact effects of chloral hydrate on cerebral glucose metabolism of pediatric patients are not known, animal studies have reported a 180% elevation of cortical extracellular glucose levels (38) and suppression of the phosphorylation process by hexokinase (39) during chloral hydrate anesthesia. During a state of generalized suppressed 18F-FDG uptake, the differences between children with PWS and children without PWS while awake are likely underestimated. Furthermore, there are differential patterns of suppression of 18F-FDG uptake in different cortical and subcortical structures induced by sedatives or during sleep. Although we found distinct cerebral metabolic features in PWS patients at a similar level of sedation with control subjects during the PET procedure, the extrapolation from the sedated state to the awake state indeed needs to be further proven. Second, the ages of PWS children in our investigation overlap the period from failure to thrive to compulsive overeating. Therefore, further analysis should be done to look at potential differences in brain metabolism during these stages of disease progression. Third, for SPM analysis, we spatially normalized PET images of child brains into the MNI template that is based on adult brains. It might be better to spatially normalize the images into the group template or an age-matched template to obtain more precise statistical results. However, our SPM analysis using an adult template yielded consistent metabolic findings across the patients. Suspicious artifacts, which were observed on the edge or outside the brain profile, could be easily differentiated by visual inspection. In addition, using a standard template (e.g., MNI template) allows more direct comparisons between data from different populations or conditions.
CONCLUSION
Our voxelwise analysis of 18F-FDG PET images of PWS children revealed metabolic abnormalities in brain regions that are directly or indirectly related with food intake and obsessive–compulsive behavior. Thus, this study describes, in part, the neural substrate underlying abnormal eating behavior and psychobehavioral problems (such as obsessions and compulsions) in PWS.
Acknowledgments
This research was supported by the Molecular and Cellular Biodiscovery Research Program (grant M10401000004-05N0100-00410), the IN-SUNG Foundation for Medical Research, and the Real Time Molecular Imaging Technology Development Program (grant M10420000007-05N2000-00710) funded by the Ministry of Science and Technology, Republic of Korea.
Footnotes
-
COPYRIGHT © 2006 by the Society of Nuclear Medicine, Inc.
References
- Received for publication September 22, 2005.
- Accepted for publication March 16, 2006.





{kind=link}