


Abstract
The endogenous opioid system of the brain has been implicated in feeding behavior. Abnormal repeated activation of this system may constitute a neural substrate for the compulsive eating behavior observed in bulimia nervosa. This study examined the binding potential of the brain μ-opioid receptor (μ-OR) in bulimia nervosa. Methods: Eight women with bulimia nervosa and 8 female controls underwent brain MRI followed by 11C-carfentanil PET. Voxel-based methods were used to assess group differences in μ-OR binding between controls and bulimic subjects and to correlate μ-OR binding with the frequency of recent self-reported abnormal eating behaviors in bulimic subjects. Results: μ-OR binding in the left insular cortex was less in bulimic subjects than in controls and correlated negatively with recent fasting behavior. Conclusion: Changes in μ-OR binding in the insula may be important in the pathogenesis or maintenance of the self-perpetuating behavioral cycle of bulimic subjects because the insula is the primary gustatory cortex and has repeatedly been implicated in the processing of the reward value of food.
Bulimia nervosa, characterized by a driven behavioral cycle of restricting, bingeing, and vomiting, shares phenomenologic similarity with the addiction disorders. This similarity suggests involvement of the opioid system in maintenance of the bulimic cycle, and high-dose naltrexone, an opiate blocker, has been used with some success in the treatment of bulimia (1). Consistent with its role in reward-driven behaviors, the endogenous opioid system of the brain modulates palatability and deprivation-induced feeding (2,3). We have previously shown in rats that cyclic food deprivation followed by access to food alters μ-opioid receptor (μ-OR) binding in limbic regions (4), suggesting that repeated activation of the opioid system by restricting and bingeing may help sustain bulimic behavior. In this study, we compared in vivo opioid function in bulimic subjects and controls and correlated μ-OR binding with abnormal eating behaviors in bulimic subjects.
MATERIALS AND METHODS
Subjects
The study was approved by the Johns Hopkins Institutional Review Board. Eight normal-weight, medication-free female subjects (mean age ± SD, 24.4 ± 4 y; range, 19–29 y) with DSM-IV–defined bulimia nervosa (5) and 8 healthy female controls (mean age, 26.4 ± 4.3 y; range, 22–32 y) provided written informed consent to participate. The body mass index, defined as weight (kg) × height (m)−2, averaged 21.7 ± 1.9 (range, 18.4–23.9) for the bulimic subjects and 23.3 ± 2.8 (range, 19.6–27.3) for the controls. Bulimic subjects were screened by a psychiatrist and underwent a full psychiatric evaluation to rule out comorbid psychiatric conditions. The Eating Disorders Examination Questionnaire, or EDE-Q (6), was administered to bulimic subjects to assess the frequency of fasting, bingeing, vomiting, laxative abuse, and excessive exercise behaviors, as well as their concerns about weight, shape, and eating, during the prior month. All controls were screened by psychiatric interview and were determined to have no prior psychiatric history and to be free of psychotropic medications. None of the controls reported fasting or having abnormal eating behaviors in the previous month.
Imaging Protocol
The methods for imaging and processing data have been described previously (7). An individually fitted thermoplastic facial mask was used during MRI and PET scans of the brain to restrict motion and allow reproducible positioning. In addition, both controls and bulimic subjects were asked not to eat within 3 h of their scheduled PET scan.
T1-weighted MR images were acquired on a 1.5-T Signa Advantage system (GE Healthcare) using 3-dimensional spoiled gradient-recalled acquisition in the steady state. PET data were acquired in 2-dimensional mode on a 4096 Plus PET scanner (GE Healthcare). A 10-min transmission scan using rotating 68Ge rods was followed by a dynamic 90-min (6 × 30 s, 5 × 60 s, 5 × 120 s, and 9 × 480 s) emission PET scan after intravenous bolus administration of 621.6 ± 74.0 MBq (range, 488.4–703.0 MBq) of 11C-carfentanil with a high specific activity of 116,416.8 ± 38,572.5 MBq/μmol (range, 69,275.1–173,567.0 MBq/μmol). Fifteen axial slices (center-to-center interslice separation, 6.5 mm) extending from the inferior temporal gyrus to the high-convexity region were acquired. The transmission scan was used for attenuation correction, and the data were reconstructed using a ramp filter into 128 × 128 2-mm-pixel matrices and were decay corrected. The final resolution, in full width at half maximum, with these parameters was 7 mm in plane.
Statistical Analysis
Statistical parametric mapping software (SPM99; Wellcome Department of Cognitive Neurology) was used for voxel-based statistical analysis. Data for the 0- to 90-min scan time were spatially normalized to an 11C-carfentanil template in the Montreal Neurological Institute space, and the transformation matrix was applied to the 34–82 min used for the subsequent analysis. Parametric maps of μ-OR binding (binding potential = [regional/occipital counts] − 1) using the 34- to 82-min spatially normalized data were smoothed using a gaussian kernel of 10 mm in full width at half maximum. A voxel-based t test comparison of control and bulimia nervosa groups was performed after analysis of covariance adjustment for global differences in μ-OR binding. The relationship between μ-OR binding and the frequency of recent (past 28 d) self-reported abnormal eating behaviors in bulimic subjects was also assessed.
Clusters showing differences between groups and correlations with frequency scores for abnormal eating behaviors were retained if the t statistic for all voxels in the cluster met a height threshold of P ≤ 0.001 and the size of the cluster had a corrected P value (Pcorr) of ≤0.05 for occurrence in the volume analyzed. These clusters are characterized by their size and their maximum t value or equivalently maximum z value (zmax) and its x, y, and z coordinates in the Montreal Neurological Institute space.
RESULTS
There were no differences (unpaired t test, P < 0.05) in age (P = 0.35), body mass index (P = 0.21), or whole-brain mean μ-OR binding (P = 0.41) between controls and bulimic subjects. In bulimic subjects, an average 17% decrease in μ-OR binding was observed in the left temporoinsular cortex (binding constant [k] = 141 voxels; Pcorr = 0.047; zmax = 3.78 at x = −48, y = −14, z = 4). No other regions showed significant group differences in μ-OR binding (Fig. 1).
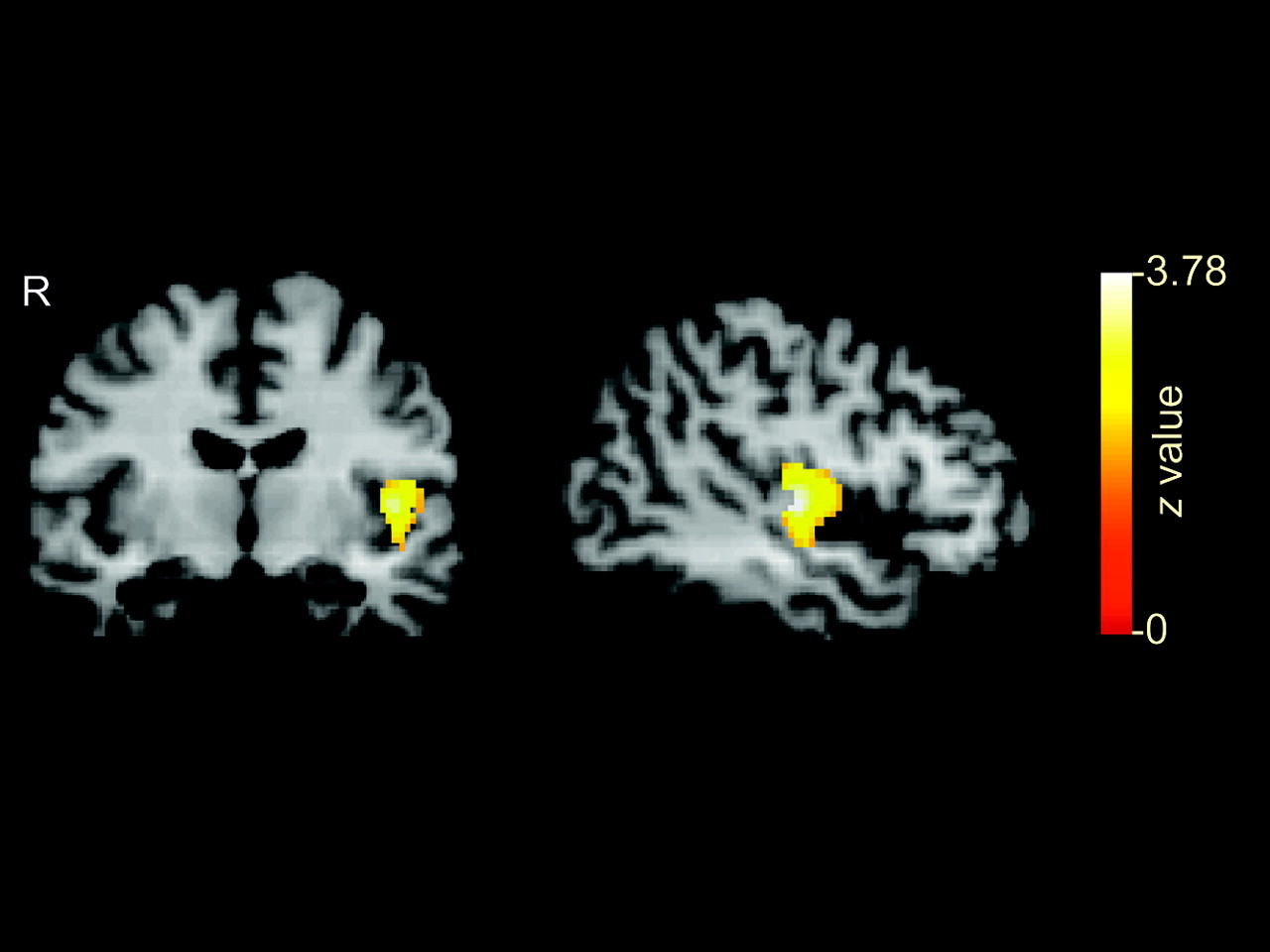
Decreased μ-OR binding in bulimia subjects relative to controls. Cluster of 142 voxels (P = 0.047; zmax = 3.78) shows decreased μ-OR binding in left temporoinsular cortex displayed on coronal and sagittal sections of coregistered brain MRI template.
Within the bulimia nervosa group, an inverse correlation of μ-OR binding with frequency of fasting behavior in the prior month was found in both the left temporoinsular cortex (k = 44; Pcorr = 0.042; zmax = 3.68 at x = −52, y = −10, z = −16) and the right temporoinsular cortex (k = 174; Pcorr = 0.031; zmax = 4.39 at x = 52, y = 0, z = 6) (Fig. 2). Among all behavioral and cognitive symptoms assessed by the EDE-Q, frequency of fasting in the prior month was the only one to show a significant statistical relationship with μ-OR binding in the temporoinsular cortex. We defined fasting using question 2 of the EDE-Q: “On how many of the past 28 days have you gone for long periods of time (8 hours or more) without eating anything in order to influence your shape or weight?”

Negative correlation between μ-OR binding and fasting behavior in bulimia subjects. Clusters were found bilaterally in temporoinsular cortex. Plot of fasting frequency in prior 28 d against average μ-OR binding for 44 voxels (P = 0.042; zmax = 3.68) in left temporoinsular cortex cluster reveals inverse linear relationship.
Although previous CT studies have reported the occurrence of a slight degree of sulcal widening in about one third of normal-weight bulimic subjects (8), the decrease in μ-OR binding and negative correlation with fasting observed in this group of bulimic subjects were not explained by structural brain loss. There was no evidence of brain atrophy on a masked examination by 2 neuroradiologists of subject’s high-quality MRI studies.
DISCUSSION
Bulimia nervosa is characterized by a driven behavioral cycle of restricting, bingeing, and vomiting and by a disturbance in the affective value of food whereby food stimuli acquire excessive salience and induce subjective experiences of increased arousal or anxiety. This emotional response presumably arises from the psychopathologic fear of being or becoming fat that is a hallmark of the disorder. We found that, compared with controls, bulimic individuals showed significantly decreased μ-OR binding in the left insular cortex. Furthermore, the lateral asymmetry noted in the insular region was consistent with findings from previous cerebral blood flow and metabolism studies on subjects with eating disorders (9,10).
The insula is the primary taste cortex, where gustatory sensory information such as taste discrimination (11), and higher cognitive functions such as the anticipation and reward of eating, are processed (9,11–15). Not only has the insular cortex been repeatedly implicated in other reward-driven behaviors, including drug abuse (16) and gambling (17), but it is also the most consistently activated area in anxiety neuroimaging studies (18).
Furthermore, the decrease in μ-OR binding we observed in bulimic subjects was proportional to their recent fasting behavior. That finding is of interest given several publications that highlight differential findings in imaging studies of the insula in the hungry versus satiated state (15,19,20). Self-defined chocolate lovers who were repeatedly scanned with H215O PET while eating chocolate showed insular activation only as long as they described the experience as rewarding, but not beyond the point of satiation (21), and increased insular cerebral blood flow has been observed on PET scans of hungry controls, compared with satiated controls, fed a liquid meal (20). Consistent with this suggestion that insular activity is related to the salience of a food reward, a recent PET cerebral blood flow study found increased insular activation in response to taste stimulation in hungry obese and postobese subjects but not in hungry normal-weight subjects (15). Compared with controls, subjects with eating disorders have been shown on functional MRI to have abnormally high neural activity in the left insular cortex in response to high-calorie food stimuli (14).
We propose that the cyclic restricting and bingeing behaviors of bulimic subjects may result in a state-related downregulation of opioid receptors as a result of chronically increased endogenous opioid release. Alternately, decreased μ-OR binding may be a trait-related marker that increases the reward value of dieting and bulimic behaviors in susceptible individuals.
CONCLUSION
These data indicate a need for future studies addressing the role of the opioid system in eating disorders. Establishing whether the differences in μ-OR binding are reversed by successful treatment of bulimia nervosa should be a first step in helping to clarify whether these alterations are state-related or trait-related. Future studies should also control for endogenous opioid release by assessing the interval between PET and the last episode of bulimic behavior and by standardizing food intake the day before scanning.
Acknowledgments
We thank Tim Moran, PhD, Pat Barta, MD, and Yu Kyeong Kim, MD, for helpful editing during the writing of the manuscript and Alexis J. Simich for providing administrative support. This study was financially supported by grant 1 R01 MH064115-01A2 from the National Institute of Mental Health and by the Johns Hopkins Institutional Research Grant Award. This study was presented at the 2002 International Conference on Eating Disorders, April 25–28, Boston, MA, and at the 47th Annual Meeting of the Society for Nuclear Medicine, June 3–7, 2000, St. Louis, MO.
Footnotes
Received Dec. 2, 2004; revision accepted Apr. 6, 2005.
For correspondence or reprints contact: J. James Frost, MD, PhD, Department of Radiology, Johns Hopkins University School of Medicine, JHOC 3225, 601 N. Caroline St., Baltimore, MD 21287.
E-mail: jfrost{at}jhmi.edu
REFERENCES
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Cerebral {mu}-opioid and CB1-receptor systems have distinct roles in human feeding behavior
- {mu} Opioid Receptors Modulate Action Potential Kinetics and Firing Frequency in Neocortical Interneurons
- Caught in the Bulimic Trap?: Persistence and State Dependence of Bulimia Among Young Women
- Sugar and Fat Bingeing Have Notable Differences in Addictive-like Behavior