


Abstract
The biologic evaluation in living rats of 68Ga-labeled oligonucleotides as imaging agents for PET is reported. Methods: 68Ga, a positron-emitting radionuclide (half-life, 68 min), along with a macrocyclic chelating agent, 1,4,7,10-tetraazacyclododecane-N,N′,N″,N‴-tetraacetic acid (DOTA), was used for labeling of antisense oligonucleotides targeting activated human K-ras oncogene. The biologic properties of 3 different forms of the oligonucleotides—that is, 2′-deoxyphosphodiester (PO), 2′-deoxyphosphorothioate (PS), and 2′-O-methyl phosphodiester (OMe)—were studied first. The biodistribution and biokinetics were evaluated in vivo in athymic rats, each bearing a tumor of A549 cells, containing K-ras point mutation in codon 12, and a tumor of BxPC-3 cells, containing wild-type K-ras. Dynamic PET imaging lasting up to 2 h was performed immediately after intravenous injection of 68Ga-oligonucleotide. Blank studies were performed using 68GaCl3 or 68Ga-DOTA alone without oligonucleotide. The 68Ga-antisense oligonucleotide uptake in tumors was also compared with the 18F-FDG and 68Ga-sense oligonucleotide uptakes. In addition, oligonucleotide binding to human plasma proteins and to human albumin was examined by means of ultrafiltration. Results: The oligonucleotides can be stably labeled with 68Ga and DOTA chelate. Intravenously injected 68Ga-oligonucleotides of 17-mer length revealed high-quality PET images, allowing quantification of the biokinetics in major organs and in tumors. The biodistribution and biokinetics of intravenously administered 68Ga-oligonucleotide varied considerably with the nature of the oligonucleotide backbone. Conclusion: We conclude that 68Ga labeling of oligonucleotides is a convenient approach for in vivo imaging and quantification of oligonucleotide biokinetics in living animals with PET.
At the moment, in vivo imaging of genes is still in its infancy (1–4). Several methods that describe the labeling of antisense oligonucleotides with positron-emitting isotopes—for example, 11C, 18F, or 76Br—have been published (5–9). Yet, only a few molecular imaging trials with experimental animals have been described (5,10). The present project focuses on the use of 68Ga-labeled antisense oligonucleotides as tracers for PET imaging, placing special emphasis on the assessment of the biologic parameters of neoplastic tissue. Alterations in the cellular genes, which directly or indirectly control cell growth and differentiation, are generally considered to be the main cause of cancer.
In a development program directed toward in vivo imaging of gene expression using PET, we have chosen K-ras oncogene as a model because of its significance in cancer biology and oncology. The ras oncogene mutations are found in a variety of human tumors but not in normal tissue, which makes ras messenger RNAs (mRNAs) suitable targets for demonstration of gene expression by noninvasive imaging with 68Ga-antisense oligonucleotides. The incidence of K-ras point mutation is extremely high (>80%) in pancreatic carcinomas (11). The sequence of K-ras oncogene is well characterized (12). Monia et al. have tested several antisense oligonucleotides targeted to ras oncogenes (13,14). Selectivity for the activated ras and the binding ability of differently modified oligonucleotides to wild-type versus mutated ras oncogenes have also been examined by others (15–18).
The natural forms of oligonucleotides—that is, 2′-deoxyphosphodiester (PO)—are unstable in vivo, degrading rapidly by nuclease digestion both intracellularly and in the systemic circulation; the half-life in plasma is about 5 min (19). Considering the nuclease sensitivity of oligonucleotides, several modifications have been explored to find good candidates possessing enhanced permeability, stability, and target affinity. Among various sugar-phosphate backbone modifications, 2′-deoxyphosphorothioate (PS) features a significantly longer half-life (ranging from a few hours to days), thus being available for the biologic target within the cell (20,21). However, the 2′-O-alkyl modifications, such as 2′-O-methyl phosphodiester (OMe), that are relatively resistant to endo- and exonucleases have a better affinity to the target (22).
In this article we present the in vitro and in vivo characteristics of 3 68Ga-antisense oligonucleotides—that is, PO, PS, and OMe sequences—specific to human K-ras oncogene activated by point mutation. Knowledge about the development and use of radiolabeled antisense oligonucleotides for in vivo imaging of specific gene targets will be of significance, considering not only the biology of tumors but also the possible applications in designing novel molecule-targeted therapies of cancer.
MATERIALS AND METHODS
68Ga Labeling of Oligonucleotides
The 17-mer PO and PS oligonucleotides bearing a 5′-aminohexyl tether and aimed at targeting codon 12 point mutation of human K-ras oncogene (23) were purchased from Scandinavian Gene Synthesis AB. A similarly derivatized OMe oligoribonucleotide was assembled by the conventional phosphoramidite strategy from the 5′-O-(4,4′-dimethoxytrityl)-2′-O-methylribonucleoside 3′-(2-cyanoethyl-N,N-diisopropylphosphoramidite) building blocks (Glen Research) and purified as described below. According to a GenBank search, the selected sequence did not have any complementary sequence in rat tissue. Oligonucleotides corresponding to the mutation-specific sense sequence were used as controls. Accordingly, the antisense and sense sequences used here were 5′-CTACGCCACTAGCTCCA-3′ and 5′-TGGAGCTAGTGGCGTAG-3′, respectively.
The amino-tethered 2′-O-methyl phosphodiester oligonucleotide was purified by ion-exchange high-performance liquid chromatography (HPLC) (SynChropak AX300, 250 × 4.6 mm; EiChrom Technologies). HPLC conditions were as follows: A/B gradient, where solution A was KH2PO4 (0.05 mol/L, pH = 5.6) in formamide:water (50:50, v/v) and solution B was solution A + (NH4)2SO4 (0.6 mol/L). Gradient elution from 10% to 70% B was applied. The product was desalted by reverse-phase (RP) HPLC (Hypersil ODS, 250 × 10 mm, 5 μm; Merck), applying gradient elution from water to 50% (v/v) aqueous acetonitrile. The identity of the oligonucleotide conjugate was verified by mass spectrometry (PE Sciex API 365 Triple Quadropole LC/ESI-MS/MS; Perkin-Elmer) and by capillary electrophoresis (HP3DCE; Hewlett-Packard).
1,4,7,10-Tetraazacyclododecane-N,N′,N″,N‴-tetraacetic acid (DOTA; Macrocyclics) and N-hydroxysulfosuccinimide (Sulfo-NHS; Fluka Chemie AG) in H2O were added to 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (Sigma Chemical Co.) in H2O and kept on ice for 30 min and then warmed to room temperature to yield DOTA-Sulfo-NHS (24). The DOTA-NHS-solution was added to the oligonucleotide in carbonate buffer (1 mol/L, pH 9) and cooled on ice. The mixture was left at room temperature for 10 h (25). The reaction mixture was sequencially purified through NAP5 (Sephadex G-25; Amersham Pharmacia Biotech AB) and C18 solid-phase extraction (SPE) columns (Supelclean LC-18 SPE tubes; Supelco). The DOTA-oligonucleotide was eluted with acetonitrile:water (50:50, v/v), evaporated, redissolved in water, and stored in 4°C until use.
68Ga was obtained in 0.1 mol/L HCl (1 mL) from a 68Ge/68Ga generator (Cyclotron C; Obninsk), and sodium acetate was added to give pH ∼5.5 (26). Then, DOTA-oligonucleotide (10–100 nmol) was added and the mixture was incubated in 100°C for 10 min. The reaction mixture was applied on the C18 SPE column, the product was eluted in ethanol:water (50:50, v/v), and the ethanol was evaporated.
The product was analyzed by radio-HPLC using a μBondapak C18 column (125Å, 10 μm, 7.8 × 300 mm; Waters); flow: 6.0 mL/min A = 20 mmol/L triethylammonium acetate buffer; B = 100% acetonitrile; C = 50 mmol/L phosphoric acid; linear gradient: 0–3 min A (100%–95%), B (0%–5%); 3–6 min A (95%–90%), B (5%–10%); 6–10 min A (90%–85%), B (10%–15%); 10–15 min A (85%–50%), B (15%–50%); 15–20 min A (50%–0%), B (50%–0%), C (0%–100%); 20–30 min C 100%; λ = 260 nm. The HPLC system consisted of LaChrom Instruments (Hitachi; Merck) and of a Radiomatic 150TR radioisotope detector (Packard).
Protein Binding Assays
The 68Ga-oligonucleotide was spiked at different concentrations to human plasma and incubated at 37°C for 30 min. Subsequently, plasma was transferred to Ultrafree-MC (Millipore) ultracentrifuge inserts (Biomax-30 polysulfone membrane with a nominal molecular weight limit of 30,000) and centrifuged at 12,300g for 20 min, collecting ∼20% of the loaded volume in the filtrate. Radioactivity in whole plasma and in the filtrate was measured using a γ-counter (1480 Wizard 3″ γ-Counter; EG&G Wallac). The fraction bound to protein was calculated by subtracting the unbound (filtrate) from 100%.
A fixed amount of 68Ga-oligonucleotide was mixed with increasing concentrations of human albumin (fatty acid-free serum albumin; Sigma Chemical Co.) and incubated for 1 h in phosphate-buffered saline containing 10 mmol/L ethylenediaminetetraacetic acid (EDTA) and 0.005% Tween 80. Blank studies were performed using 68GaCl3 or 68Ga-DOTA alone without oligonucleotide. After the incubation, the samples were loaded onto Microcon YM-30 filters (regenerated cellulose membranes with a nominal molecular weight limit of 30,000; Millipore) and centrifuged at 9,500g for 10 min. Radioactivity was measured from both the filtrate and the unfiltered solutions, and the amount of oligonucleotide bound to albumin was calculated as described above.
Hybridization in Solution
The 68Ga-antisense oligonucleotide (PO, PS, and OMe) in 50% ethanol was evaporated and dissolved in TES buffer (50 mmol/L Tris, pH 8.0, 50 mmol/L NaCl, 1 mmol/L EDTA). The hybridization mixture was prepared as follows: A gradually increased concentration of PO sense oligonucleotide (0.33–80 pmol) was added to a constant concentration of 68Ga-oligonucleotide (20 pmol), and the total volume was adjusted to 10 μL with TES buffer. As reference solutions, 40 pmol of 68Ga-oligonucleotide or unlabeled PO sense oligonucleotide were used. All mixtures were kept on ice until hybridization at 40°C for 10 min. Subsequently, samples were run on a 20% polyacrylamide gel electrophoresis (PAGE) gel along with a molecular weight marker. After electrophoresis, the gel was stained using ethidium bromide and photographed under ultraviolet light. Then, the gel was exposed to a phosphor imaging plate for 12 h, scanned using a Phosphorimager SI device, and analyzed using Image Quant 5.1 software (Molecular Dynamics Inc.).
RT-PCR and PCR
The mRNA and genomic DNA were isolated from A549 tumor cells bearing point mutated K-ras gene. The mRNA was converted into single- strand complementary DNA (cDNA) by a sequence of reverse transcription (RT) reaction and polymerase chain reaction (PCR), using a RobusT I RT-PCR kit, which is based on avian myeloblastosis virus reverse transcriptase (AMV-RT) in the first-strand cDNA synthesis and on DyNAzyme EXT DNA polymerase in the second-strand synthesis (Finnzymes Oy). Unlabeled, DOTA-conjugated, or 68Ga-oligonucleotides were used as reverse primers. The sequence of the forward primer (Pr1) was 5′-GACTGAATATAAACTTGTGG-3′. In a control reaction, the reverse primer (Pr2) had a sequence of 5′-CTATTGTTGGATCATATTCG-3′, allowing amplification of a 108-base-pair (bp) PCR product around codon 12 of human K-ras oncogene as described earlier (27). The protocol for RT-PCR amplification was as follows: 40 min at 48°C followed by 2 min at 94°C (1 cycle); denaturing at 94°C for 1 min followed by annealing at 52°C for 1 min and extension at 72°C for 1 min (35 cycles); and final extension at 72°C for 5 min. The DNA samples underwent 40 cycles of PCR: 1 min at 94°C, 1 min at 52°C, and 1 min at 72°C.
The PCR products were run on a 10% PAGE gel along with a molecular weight marker. The DNA bands were visualized with ethidium bromide and photographed under ultraviolet light.
Nuclease Stability
The 68Ga-oligonucleotide was mixed with heparinized human blood and incubated for 5, 30, 60, or 120 min at 37°C. Plasma retained after centrifugation was filtered through Microcon YM-30 filters for the separation of proteins and analyzed by radio-HPLC as described above.
Tumor Model
Two human tumor cell lines—that is, A549 lung carcinoma cell line harboring codon 12 mutated K-ras oncogene and BxPC-3 pancreatic adenocarcinoma cell line harboring wild-type K-ras—were purchased from American Type Culture Collection and grown in RPMI 1640 medium containing 10% fetal bovine serum without antibiotics. Male athymic Hsd:RH-rnu/rnu rats were obtained from Harlan, Netherlands, at the age of 6 wk and kept in the Central Animal Laboratory of the University of Turku in filter-top cages and allowed food and water ad libitum. Tumor cells (107 per rat) were injected subcutaneously into the flank region, and tumors were allowed to grow to a size of 1–2 cm in diameter. All animal studies were approved by the local University Laboratory Animal Committee.
Dynamic PET Studies
For PET imaging, rats weighing 353 ± 44 g were anesthetized with intraperitoneal. injection of pentobarbital (60 mg/kg; Mebunat Vet, Orion). PET studies were performed with an Advance PET scanner (General Electric Medical Systems) operated in 2-dimensional mode. The scanner has 18 rings of bismuth germanate detectors and the axial length of the imaging field of view (FOV) is 152 mm. All 35 transaxial image slides were reconstructed with an ordered subsets expectation maximization algorithm and the central 200-mm-diameter transaxial FOV was used. the image pixel size was 1.56 × 1.56 mm in a 128 × 128 matrix. Scatter correction, random counts, and dead time corrections were all incorporated into the reconstruction algorithm. Radiation attenuation in each animal was measured with 2 rotating rod sources containing 68Ge/68Ga before tracer injection, and the images were corrected for radiation attenuation.
All PET studies started with an 8-min transmission scan. The tracer was injected intravenously via the tail vein as a bolus. The median dose was 11.49 ± 4.52 MBq. The dynamic scanning started immediately after injection. The acquisition times were as follows: 6 × 10 s, 4 × 60 s, and 23 × 300 s (duration, 120 min).
Regions of interest were drawn on major organs and on the tumors. Pharmacokinetic curves, representing the radioactivity concentrations (percentage of injected dose per gram of tissue = %ID/g), versus time after injection were determined accordingly.
Metabolite Analysis
Immediately after PET imaging, blood was obtained by cardiac puncture, followed by an intracardial pentobarbital injection for killing each animal. Subsequently, urine was obtained directly from urinary bladder. Both plasma retained after centrifugation and urine were filtered through Microcon YM-30 filters by centrifugation at 9,500g for 10 min for the separation of proteins. Filtrates were analyzed by radio-HPLC as described above.
Organ Distribution
Samples of blood, urine, tumors, heart, lung, liver, kidney, pancreas, spleen, intestine, adrenal, urinary bladder, testis, brain, muscle, bone, bone marrow, skin, and parotid gland were excised, weighed, and measured in a calibrated γ-counter for 30 s (1480 Wizard 3″ γ-Counter). The radioactivity concentration was expressed as standardized uptake value ([organ activity/organ weight]/[total given radioactivity/rat body weight]).
RESULTS
68Ga Labeling of Oligonucleotides
The 68Ga labeling of oligonucleotides is straightforward and applicable to any sequence and oligonucleotide chemistry. The 68Ga-labeling method was originally developed at Uppsala PET Centre and the Department of Organic Chemistry of Uppsala University, Sweden, and later set up at Turku PET Centre, Finland. According to our experience, the method is easily transferable from one laboratory to another and requires only minor investments, the generator being the most expensive acquisition.
In the present study, the reaction of 68GaCl3 with DOTA-conjugated oligonucleotide was rapid—that is, 10 min at 100°C. In general, 25 MBq of purified 68Ga-oligonucleotide were obtained from 100 MBq of 68GaCl3 and 55 nmol of DOTA-conjugated oligonucleotide in 30 min with 95%–99% radiochemical purity. The specific activity of 68Ga-oligonucleotide was 270 ± 90 MBq/mg. According to the radio-HPLC analysis, free 68Ga and its DOTA chelate were the main impurities, both present at a very low concentration. The 68Ga-oligonucleotide remained stable in H2O at room temperature for >4 h as demonstrated by repeated radio-HPLC analyses (data not shown).
Biologic Properties of 68Ga-Oligonucleotides
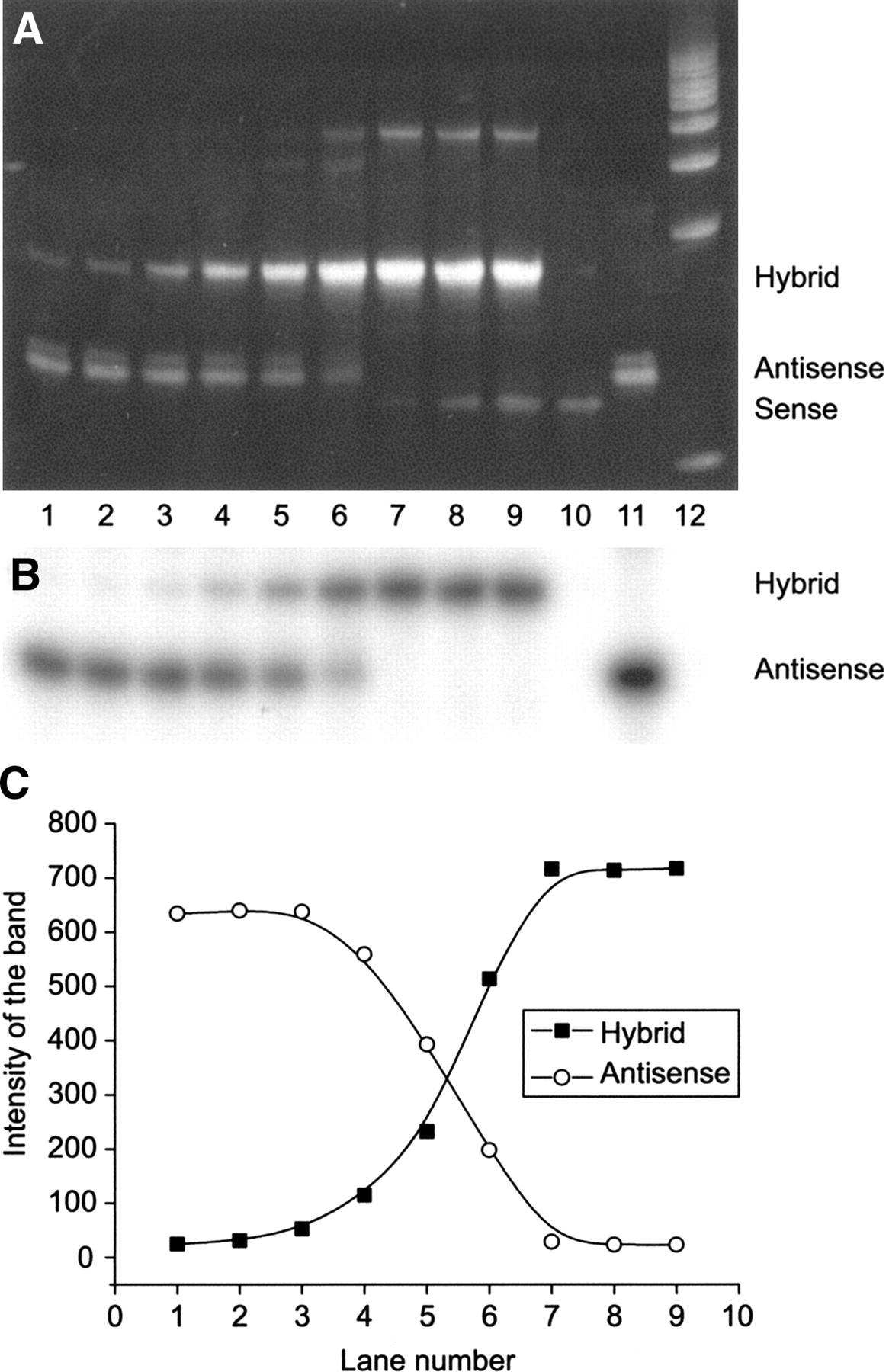
The ability of 68Ga-oligonucleotides to hybridize with a complementary sequence was investigated in a cell-free approach. Our results revealed that 68Ga labeling of oligonucleotides did not alter their hybridization properties. Instead, the hybridization of 68Ga-oligonucleotides with a PO sense oligonucleotide of the same length was concentration dependent (Fig. 1). The stained PAGE gels showed that all 3 68Ga-oligonucleotides (PO, PS, and OMe) were able to hybridize. The hybrid formation increased proportionally with the concentration of sense oligonucleotide.

Hybridization of 17-mer 68Ga-labeled PO antisense oligonucleotide with unlabeled PO sense oligonucleotide of the same length in a cell-free approach. Ethidium bromide-stained (A) and autoradiographic (B) images of 20% PAGE gels demonstrate the concentration-dependent hybrid formation (C). The concentration ratios of antisense:sense in the lanes are as follows: lane 1, 1:1/60; lane 2, 1:1/30; lane 3, 1:1/15; lane 4, 1:1/5; lane 5, 1:1/2; lane 6, 1:1; lane 7, 1:2; lane 8, 1:3; lane 9, 1:4, and the references are lane 10, unlabeled PO sense oligonucleotide; lane 11, 68Ga-labeled PO antisense oligonucleotide; and lane 12, a molecular weight marker.
Interestingly, only PO worked as a reverse primer both in RT-PCR and in PCR reactions. This property was retained after conjugation with DOTA and also after labeling with 68Ga, as shown in Figure 2. No amplification with PS or OMe was observed in either case.

Ethidium bromide-stained 10% PAGE gel of the PCR and RT-PCR products, using either unlabeled, DOTA-conjugated, or 68Ga-labeled PO antisense oligonucleotide as a reverse primer. A 41-bp fragment corresponding to the K-ras codon 12 region is seen. The A549 cells (human lung carcinoma) contain K-ras oncogene point mutation in codon 12. MW = molecular weight marker.
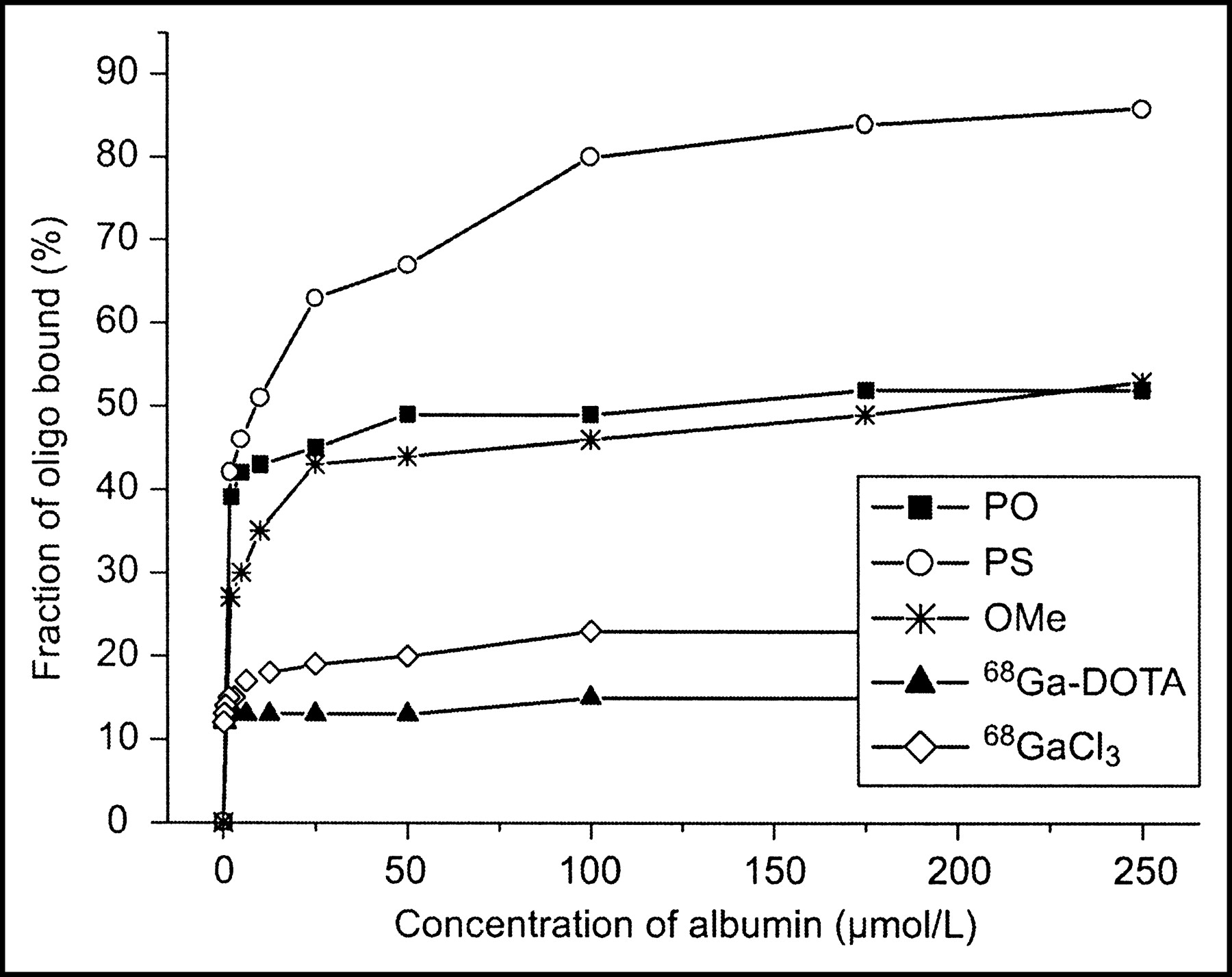
The binding of 68Ga-oligonucleotides to human plasma proteins was studied using the ultrafiltration method. Binding to human albumin seemed to be dependent of the backbone chemistry; PS was bound to the highest extent, and the binding clearly exhibited saturation (Fig. 3). The percentage of protein binding (10%–20%) for both 68Ga-DOTA and 68GaCl3 was much lower compared with that of labeled oligonucleotides (Fig. 3). Binding to whole plasma proteins was 99%, 99%, and 96% for PS, OMe, and PO, respectively.

Albumin binding of 68Ga-labeled oligonucleotides (oligo). Mean value of 4 experiments (n = 4), except for blank studies with 68GaCl3 and 68Ga-DOTA alone without oligonucleotide (n = 2).
PET Imaging and Quantification of 68Ga-Oligonucleotide Biokinetics in Living Rats
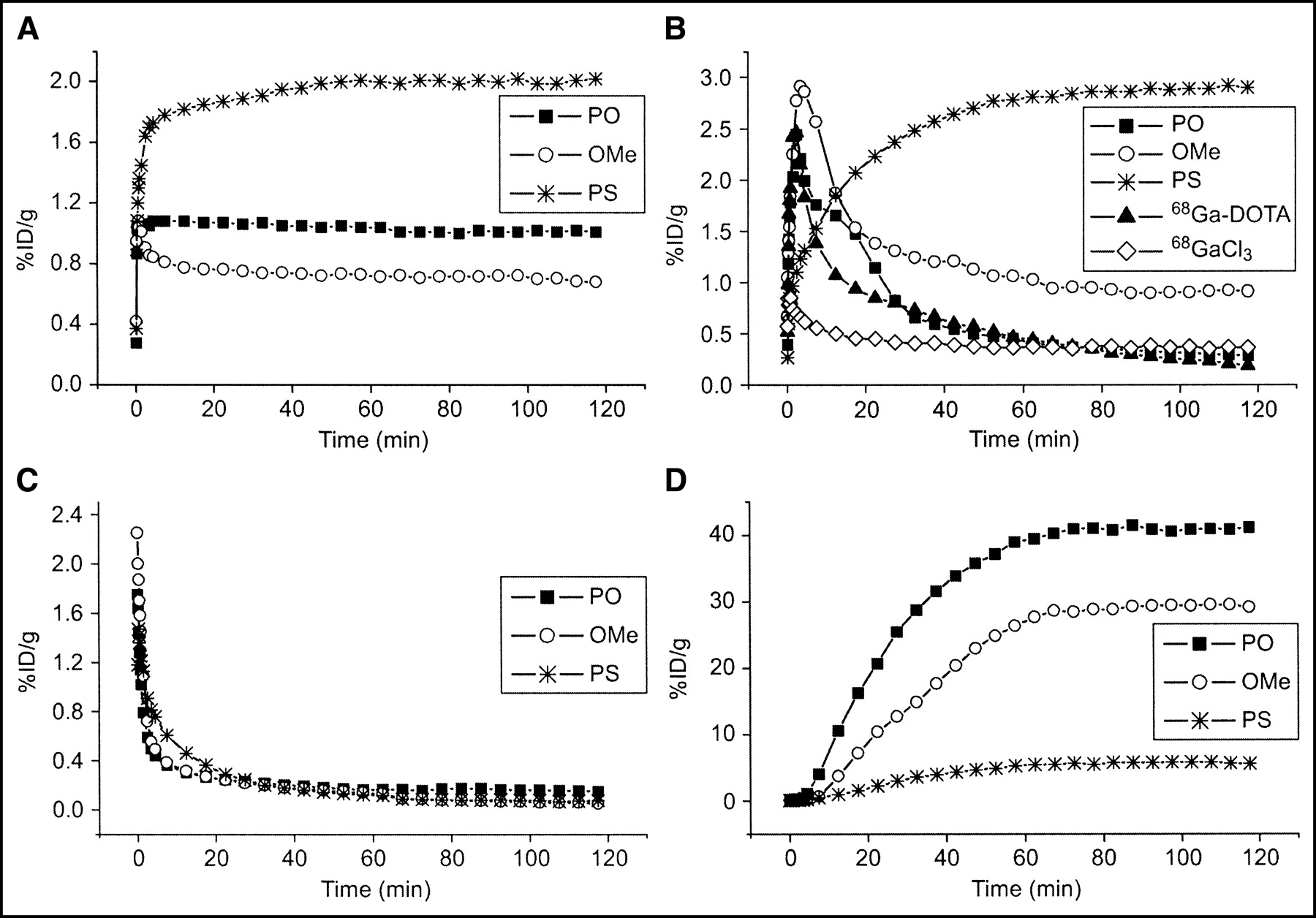
PET imaging was conducted for tumor-bearing rats, using an Advance PET scanner and 68Ga-oligonucleotides with 3 different backbone chemistries. The 68Ga-antisense oligonucleotide uptake in main organs and in tumors was compared with 18F-FDG, 68GaCl3, 68Ga-DOTA, and 68Ga-sense oligonucleotide uptakes. The quantification of biokinetics data—that is, the amount of radioactivity normalized for the injected dose versus time after injection in rat liver, kidney, heart, and bladder—is shown in Figure 4. The biokinetic differences between PO, PS, and OMe are evident.

In vivo biokinetics (amount of radioactivity vs. time after injection) of intravenously administered 17-mer 68Ga-labeled antisense oligonucleotides specific to codon 12 mutated human K-ras oncogene in rat liver (A), kidney (B), heart (C), and bladder (D). Biokinetics of 68GaCl3 and 68Ga-DOTA without oligonucleotide in rat kidney are also shown. Radioactivity concentrations are expressed as %ID/g of tissue. Mean value of 4–6 rats (n = 4–6).
Tumor Localization of 68Ga-Oligonucleotides
After the last PET imaging, the rats were killed, and tumors, blood, urine, and different organs were excised for the measurement of radioactivity. The tumor-to-blood and tumor-to-muscle ratios were calculated for PO, PS, and OMe (Table 1). Preliminary studies revealed that PS and OMe localize somewhat better in A549 tumors in contrast to their PO counterpart (n = 1–3). The slightly better uptake of PS in an A549 tumor, compared with a BxPC-3 tumor, was observed, as shown in Figures 5 and 6.

PET image of an athymic rat bearing both A549 and BxPC-3 tumors, using 68Ga-labeled PS antisense oligonucleotide specific to codon 12 mutated human K-ras oncogene. The human A549 cells (lung adenocarcinoma) contain K-ras point mutation in codon 12, and the human BxPC-3 cells (pancreatic adenocarcinoma) contain wild-type K-ras oncogene. Image is a summation from 60 to 65 min after injection and is color-coded according to the amount of radioactivity, from dark blue (lowest) to hot red (highest).
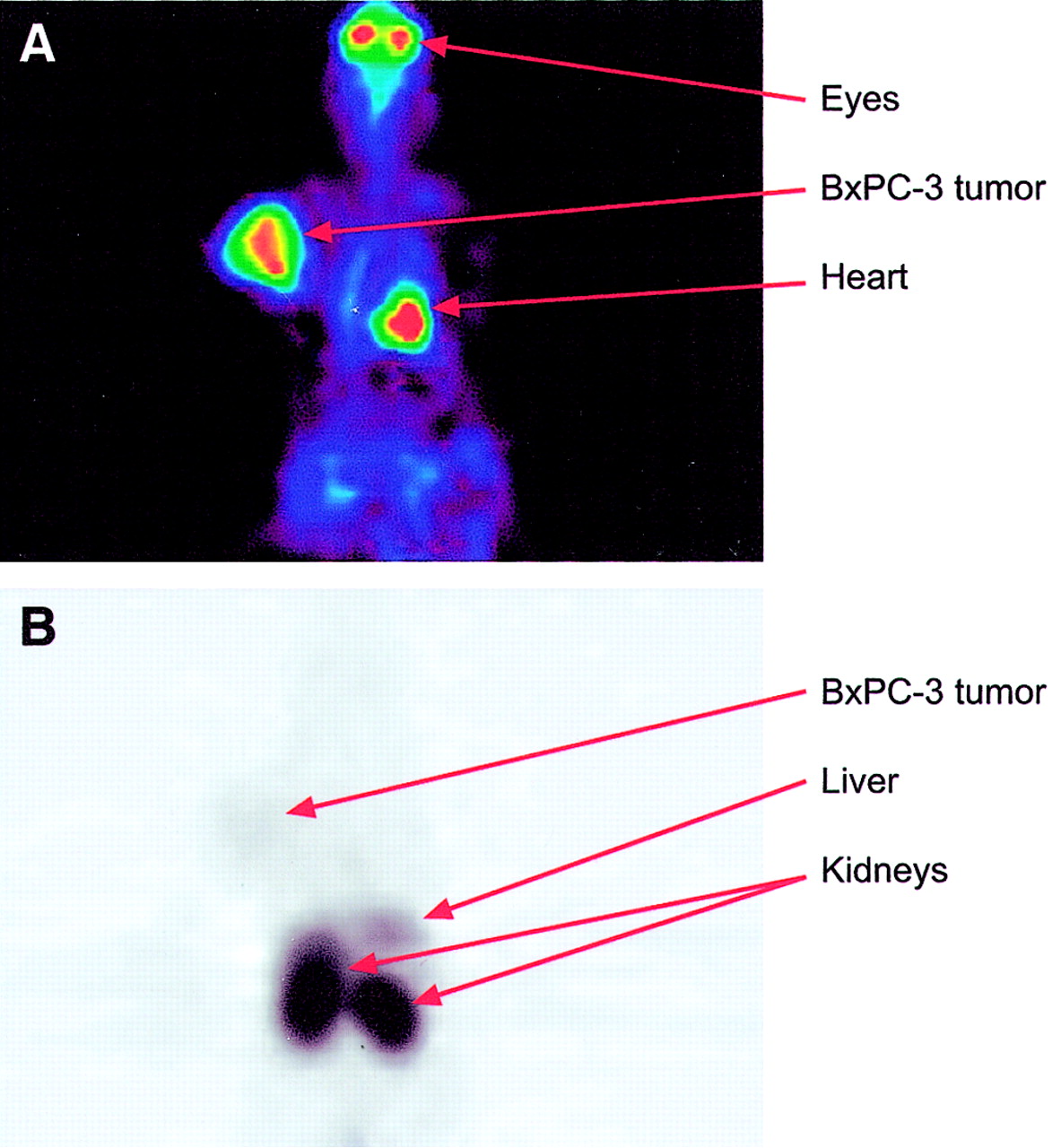
Whole-body PET images of a tumor-bearing rat. The animal underwent PET imaging with both 18F-FDG and 68Ga-labeled PS antisense oligonucleotide on separate days. Images are summations from 90- to 95-min follow-up measurements after injection. (A) BxPC-3 tumor (human pancreatic adenocarcinoma) with wild-type K-ras oncogene is clearly visible with 18F-FDG. (B) 68Ga-PS oligonucleotide shows high kidney and liver uptake, compared with low BxPC-3 tumor uptake. Note that this animal did not have an A549 tumor (human lung carcinoma) with point mutated K-ras gene to which the antisense oligonucleotide was specific.
Mean ± SD Values of Tumor-to-Blood and Tumor-to-Muscle Ratios of 68Ga-Labeled Antisense Oligonucleotides Specific to Point Mutated Human K-ras Oncogene
In Vivo Stability of 68Ga-Oligonucleotides
Samples of plasma and urine were also subjected to radio-HPLC analysis for the evaluation of in vivo stability of 68Ga-oligonucleotides. Representative radiochromatograms are shown in Figure 7. After 120 min, most of the radioactivity in urine derived from free 68Ga-chelate and short 5′-terminal fragments of the probe, not from the intact 17-mer PS oligonucleotide. In plasma filtrate, on the other hand, the PS appeared to be almost intact (Fig. 7B). In contrast, both PO and OMe were totally degraded by nucleases during the 120-min incubation time both in plasma and in urine (data not shown).
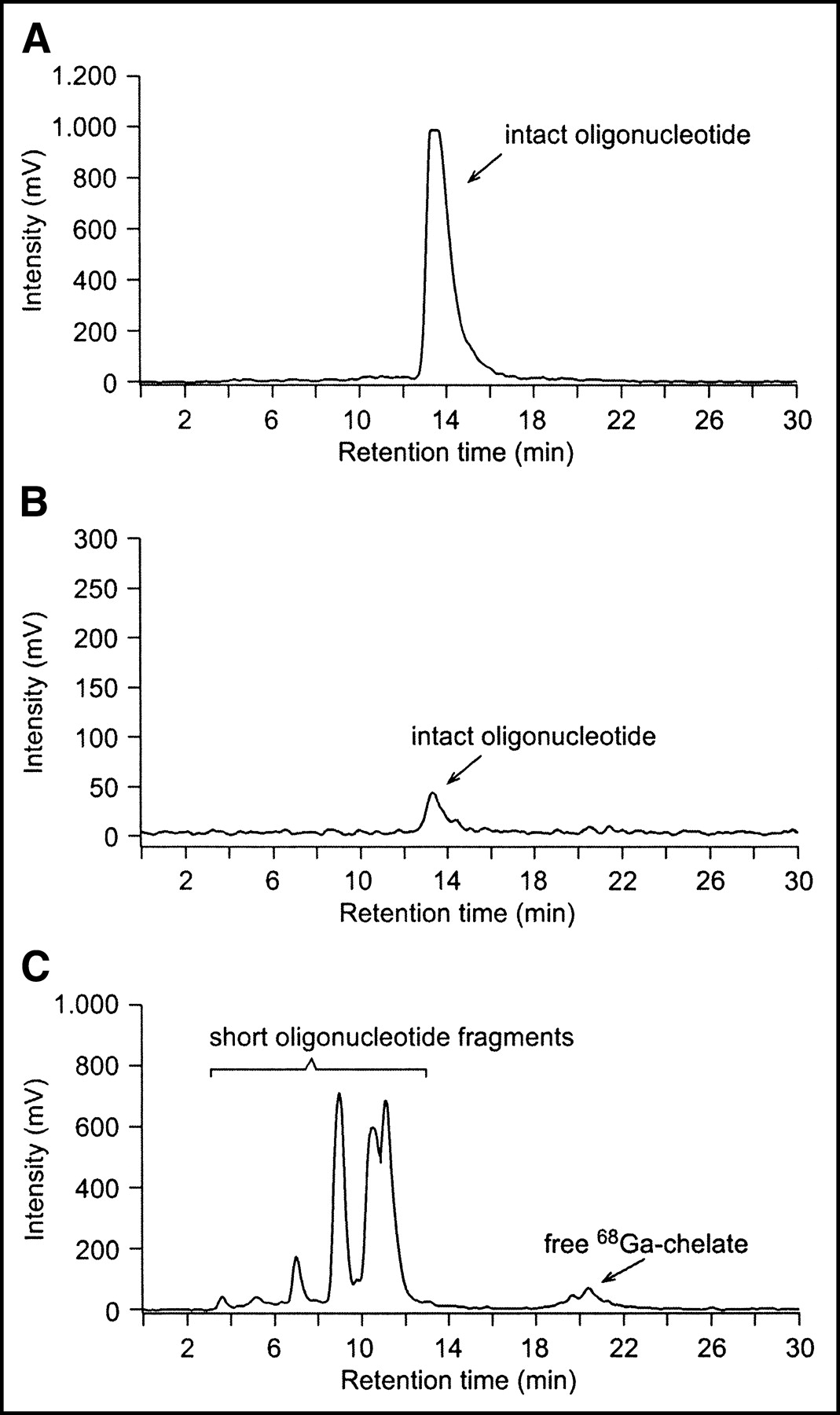
Radio-HPLC chromatograms of intact oligonucleotide (A), rat plasma (B), and rat urine (C) taken 120 min after intravenously administered 68Ga-labeled PS oligonucleotide. Most radioactivity in urine derives from free 68Ga-chelate and short oligonucleotide fragments. In plasma, the oligonucleotide appears to be almost intact.
DISCUSSION
Antisense oligonucleotides labeled with positron-emitting isotopes 11C, 76Br, and 18F have already been studied as potential PET tracers for molecular imaging (5,7,9). In this study we have used oligonucleotides labeled with generator-produced 68Ga possessing a sufficiently long half-life (68 min) and appropriate mode of decay (90% β+, 10% electron capture) for PET imaging. With respect to PET imaging, 68Ga has the following advantages over 18F, 11C, or 76Br: (a) The isotope itself is readily available using a low-cost 68Ge/68Ga generator system instead of elaborate and expensive cyclotrons. (b) The half-life is longer than that of 11C (68 min vs. 20 min). (c) The β+ decay energy of 68Ga (90% positrons, 2.921 MeV) is ideal for PET imaging. For comparison, 76Br has a longer half-life (16.2 h) but it decays only 55% by positron emission. Thus, 76Br is suitable for long-lasting in vitro applications, but not for human PET studies. For this study, we investigated the biodistribution and biokinetics of intravenously administered 68Ga-labeled 17-mer PO, PS, and OMe antisense oligonucleotides specific for codon 12 point mutated human K-ras oncogene using tumor-bearing rats.
One of the major challenges in the field of antisense research is the identification of sequences within the target mRNA that would be accessible to an antisense oligonucleotide. Owing to the 3-dimensional folding of mRNA, there are both inaccessible segments and double-stranded regions in the RNA molecule. In addition, for the in vivo applications of antisense oligonucleotides certain hurdles need to be overcome—for example, nuclease resistance, cellular transport and penetration, intracellular distribution, and turnover of the oligonucleotide. Modifications of oligonucleotides to increase their stability must be made without a loss of specificity or hybridization properties. To meet these demands, dozens of structurally modified oligonucleotides have been prepared, but apart from the extensively used PS oligonucleotides (20,28), only few of them have been properly screened in vivo. 2′-O-Alkylation of oligoribonucleotides (29), modifications at the base moiety structures (30), and freezing of the sugar ring puckering by an additional covalent bridge (31) have been shown to enhance the hybridization, whereas biodegradable lipophilic phosphate protective groups have been suggested to increase the cellular uptake (32). In practice, compromises between permeability and specificity of hybridization are inevitable. It is known that sequences of 15–20 nucleotides will enter the cytoplasm and bind mRNA (33). Although shorter oligonucleotides enter into the cells through the cell membrane, they may not attach strongly enough or may not be unique to bind the specific mRNA. Because the mRNA maintains a 3-dimensional structure with hairpins and bulges, we have to select a sequence that will bind without mismatch of a single base.
Other major concerns for antisense approaches are the cellular internalization and intracellular distribution; the mechanism by which oligonucleotides are taken up into cells is still poorly understood (34). The majority of oligonucleotides—for example, PO, PS, and OMe—are negatively charged molecules and behave as polyanions regardless of their sequence content. A hydrophilic oligonucleotide inefficiently crosses a hydrophobic cell membrane. Therefore, several attempts have been made to enhance the cellular delivery of oligonucleotides (35). So far, the most commonly and successfully used delivery systems are liposomes and charged lipids, enabling internalization by endocytosis. Essentially, after reaching the endosomal compartment, the oligonucleotide must be efficiently released and set free for binding with the specific target mRNA.
In the development of new radiopharmaceuticals for PET, high specificity and high affinity of binding are important properties. Rapid localization and clearance kinetics of the tracer and its prolonged retention in the target are general requirements for nuclear imaging, whereas ribonuclease H-dependent turnover is usually desired in antisense chemotherapy. Accordingly, a particular oligonucleotide that is found to be unsuitable for antisense chemotherapy may be the molecule of choice for antisense imaging. For successful antisense imaging, target cells should have a sufficient amount of mRNA. Furthermore, the labeling of the antisense compound should not interfere with base pairing between the mRNA and the antisense molecule.
At the moment, only a few PET imaging trials with labeled oligonucleotides have been reported. Previously, Kobori et al. have successfully visualized mRNA expression in the central nervous system using PET. The PS antisense oligonucleotide for mRNA of glial fibrillary acidic protein labeled with 11C was retained in tumor cells, yielding clear images of gliomas in rats (5). Tavitian et al. labeled PS antisense oligonucleotide with 18F and performed in vivo PET studies in baboons (10). The study was designed only to evaluate the pharmacokinetics of the tracer, not to determine its hybridization in vivo.
The use of reporter genes instead of oligonucleotides is another alternative to in vivo imaging of gene expression. Current research in this particular field of imaging is productive. For example, PET imaging of the dopamine D2 receptor in mice, using a derivative of spiperone (3-(2′-18F-fluoroethyl)spiperone [18F-FESP]) as a reporter gene, has been reported (36). Furthermore, imaging of herpes virus type 1 thymidine kinase (HSV1-tk) gene transfer and expression in vivo, using 124I-5-iodo-2′-fluoro-1-β-d-arabinofuranosyluracil (124I-FIAU) and PET, have been studied (37). Gambhir et al. used 18F-fluoroganciclovir (18F-FGCV) for HSV1-tk and demonstrated its accumulation in living mice with PET (38).
Although labeling of oligonucleotides with radionuclides is becoming fairly common, to our knowledge, radiolabeling with 68Ga has not been reported by others. Considering the low abundance of mRNA targets, high specific activities of 68Ga-labeled oligonucleotides are essential. In this study, the average specific activity of all 68Ga labeling was 270 ± 90 MBq/mg, which converts to 1.6 ± 0.5 GBq/μmol. A specific activity of 74 MBq/mg for 90Y-labeled PS oligonucleotides has been reported previously (39). On the other hand, compared with, for example, 18F-labeled (111–195 GBq/μmol) or 125I-labeled (37 GBq/μmol) oligonucleotides, the obtained specific activity is much lower (9). Yet, for the present study, only 10–100 nmol of DOTA-conjugated oligonucleotide was used for the labeling reaction with 68Ga. Our preliminary studies reveal that the radiochemical yield improves with increasing concentration of oligonucleotide, which most probably increases the specific activity also (I. Velikyan, G. Lendvai, M. Välilä, et al., unpublished data, 2003).
For in vivo antisense applications, it is important to clarify how the 68Ga labeling of oligonucleotides influences their biologic properties. Our studies have revealed that the conjugate labeling with 68Ga using DOTA as a chelator did not alter the hybridization or protein binding capability of the oligonucleotides. High protein binding of PS, for example, has also been reported by others (21). Furthermore, the in vitro albumin binding studies showed that the binding was clearly saturable. Such studies are relevant since the protein binding affects both tissue distribution and cellular uptake. It has been suggested that plasma proteins—for example, albumin—can act as carrier proteins and protect oligonucleotides from degradation.
The negative PCR and RT-PCR results of PS and OMe are of interest. In general, a desired PCR product is amplified if (a) the oligonucleotide primers are able to bind complementarily to the template sequence (annealing) and (b) the DNA polymerase enzyme can attach and start copying the template (extension). Our results suggest that PS and OMe are able to inhibit the primer extension most likely by directly interacting with the enzyme. Indeed, oligonucleotides carrying a PS modification are strong inhibitors of DNA polymerases in a linkage number-dependent manner. Longer oligomers are more potent inhibitors than shorter ones, and the inhibitory effect can be avoided by decreasing the number of PS linkages at the backbone. Our negative PCR results, obtained with fully thioated oligonucleotides, are in accordance with these observations. In addition, both PS and OMe oligomers can inhibit polymerization by binding to the AMV-RT rather than to the template (40).
The overall distribution of oligonucleotides relies, among other things, on protein binding in plasma and tissues, passive diffusion across the membranes, and transport into or out of the cells. The elimination kinetics in vivo of our oligonucleotides seemed to vary significantly with the nature of oligonucleotide backbone. Since these oligonucleotides did not have any biologic target in normal rats, the radioactivity distribution to various organs basically reflects their nonspecific interactions and metabolism. The biodistribution of the 3 oligonucleotides studied presumably relates to their in vivo degradation versus stability. The results obtained partially agree with those reported by Tavitian et al. on 18F-oligonucleotides (10). Yet, the obvious kidney accumulation of PS is of interest. This behavior seems not to be caused by DOTA, 68Ga, or the oligonucleotide sequence, but by the backbone itself. Yet, previous studies have shown that the length of PS oligonucleotides has a major impact on the body distribution pattern (7).
CONCLUSION
Our results on the biokinetics of 68Ga-oligonucleotides in rats, determined using an GE Advance PET scanner, are encouraging. Intravenously injected 68Ga-oligonucleotides of 17-mer length revealed high-quality PET images, allowing quantification of the biokinetics in major organs and in tumors. Further studies in a larger number of rats include clarification of the metabolic fate of 68Ga-oligonucleotides in tumors and, most importantly, clarification of the in vivo specificity—that is, hybridization with the mutant K-ras mRNA—by performing comparative experiments with antisense, sense, and missense oligonucleotides. Our results show, however, that the simple and rapid 68Ga labeling of oligonucleotides can be applied for PET imaging. We conclude that 68Ga labeling of oligonucleotides is a convenient approach for in vivo PET imaging of the biodistribution and quantification of oligonucleotide biokinetics in living animals.
Acknowledgments
The authors thank Maija-Liisa Hoffren (SafetyCity Ltd. Oy, Turku, Finland) for excellent assistance with the rat experiments. This work was financially supported by grants from the Foundation of Finnish Cancer Institute, the Instrumentariun Foundation, the Nordic Cancer Union, the Turku University Foundation, and the Turku University Hospital.
Footnotes
Received Jun. 26, 2003; revision accepted Oct. 29, 2003.
For correspondence contact: Anne Roivainen, PhD, Turku PET Centre, Turku University Hospital, P.O. Box 52, FIN-20521 Turku, Finland.
E-mail: anne.roivainen{at}pet.tyks.fi
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Multifunctional Imaging Signature for V-KI-RAS2 Kirsten Rat Sarcoma Viral Oncogene Homolog (KRAS) Mutations in Colorectal Cancer
- In vivo imaging of pyrrole-imidazole polyamides with positron emission tomography
- A Generalizable Strategy for Imaging pre-mRNA Levels in Living Subjects Using Spliceosome-Mediated RNA Trans-Splicing
- Receptor-mediated internalization of chelator-PNA-peptide hybridization probes for radioimaging or magnetic resonance imaging of oncogene mRNAs in tumours
- Uptake Kinetics of 99mTc-MAG3-Antisense Oligonucleotide to PCNA and Effect on Gene Expression in Vascular Smooth Muscle Cells
- 68Ga-Labeled Peptides in Tumor Imaging
- External Imaging of CCND1 Cancer Gene Activity in Experimental Human Breast Cancer Xenografts with 99mTc-Peptide-Peptide Nucleic Acid-Peptide Chimeras