


Abstract
Our goal in this investigation was to develop a method for iodinating annexin V that would be suitable for the in vivo detection of apoptosis. Methods: Annexin V was iodinated with 125I using 2 different techniques: direct iodination with IODO-BEADS, resulting in the iodination of tyrosine residues; and use of the Bolton-Hunter reagent, which binds to lysine. The active fraction of the labeled preparation was purified by affinity chromatography. We assessed thyroid accumulation of free iodide by comparing mice with blocked and unblocked thyroids. We tested the ability of iodinated annexin V to bind apoptotic cells in vitro using irradiated neuroblastoma cells and immobilized phosphatidylserine and in vivo using C3H mice subjected to whole-body irradiation. Results: The efficiency of IODO-BEADS iodination was just below 30%; with the Bolton-Hunter protocol we were able to achieve 40% efficiency. When the IODO-BEADS-labeled preparation was injected into nude mice, activity accumulated rapidly in the thyroid. Two hours after injection, uptake in the thyroid region was clearly visible on a gamma-camera scan. This uptake was absent in mice that had had their thyroids blocked. We concluded that the IODO-BEADS method of labeling resulted in a protein that was rapidly deiodinated in vivo. By contrast, when annexin V was labeled using the Bolton-Hunter protocol, there was no evidence of activity accumulating in the thyroid. The Bolton-Hunter-labeled annexin V bound to apoptotic cells and immobilized phosphatidylserine in vitro. The active fraction of Bolton-Hunter-labeled annexin V was approximately 0.75. In C3H mice given 5-Gy whole-body irradiation, there was a significant induction of apoptosis in the spleen, as measured by the terminal deoxynucleotidyl transferase deoxyuridine triphosphate nick-end labeling assay, and a 4-fold increase in 125I activity in the spleens relative to that of the control animals. Conclusion: Direct iodination of annexin V on tyrosine residues is a poor technique suffering from rapid deiodination in vivo. With Bolton-Hunter chemistry, one can produce a molecule that retains its label in vivo and binds to apoptotic cells in vitro and in vivo.
Healthy cells and, to a lesser extent, malignant cells undergo apoptosis or programmed cell death in response to a variety of stimuli, including oxygen deprivation and cytotoxic drugs. In apoptosis, a complex signaling pathway is activated, leading to loss of cytochrome c from the mitochondria; consequent activation of the caspase family of proteases; and, ultimately, enzymatic digestion of the cell’s own DNA (1). At an early stage in this process the cell membrane changes so that phosphatidylserine (PS), a lipid normally present on the membrane’s inner surface, is exposed on the outer surface (2). This change in the membrane can be detected by the binding of annexin V to the external PS, and this has formed the basis for an in vitro assay for apoptosis (3).
Blankenberg et al. (4) have applied annexin V to the in vivo imaging of apoptosis by labeling annexin V with 99mTc. With this technique, they have been able to image apoptosis in murine lymphoma, in rat coronary grafts (5), in rats rejecting lung transplants (6), and in mouse livers after anti-FAS treatment (7). In humans, 99mTc-annexin V localizes to the kidney and liver (8).
To extend the use of annexin V to PET, it would be very desirable to iodinate the molecule. 124I has a relatively long half-life (4.18 d) for a positron emitter and is relatively easy to make compared with 94Tc. Lahorte et al. (9,10) have shown the feasibility of iodinating annexin V with immobilized N-chlorobenzenesulfonamide (IODO-BEADS; Pierce, Rockford, IL). In this article, we examine the biodistribution in mice of IODO-BEADS-labeled annexin V and compare it with a Bolton-Hunter-labeled compound.
MATERIALS AND METHODS
Chemicals
Na125I in 10 μmol/L NaOH was purchased from New England Nuclear (Boston, MA). Water-soluble Bolton-Hunter reagent and IODO-BEADS were obtained from Pierce; chloramine-T and sodium metabisulfite were obtained from Aldrich (St. Louis, MO); human recombinant annexin V and anti-FAS antibody, clone Jo2, were supplied by Pharmingen (San Diego, CA); and PS, cholesterol, and bovine serum albumin (BSA) were purchased from Sigma (St. Louis, MO). Terminal transferase and biotin-16-deoxyuridine triphosphate (dUTP) were purchased from Roche (Indianapolis, IN); avidin/biotin blocking reagent, avidin-biotin-horseradish peroxidase complex, and diaminobenzidine were obtained from Vector Laboratories (Burlingame, CA); and Dulbecco’s modified essential medium (DMEM) and fetal bovine serum (FBS) was obtained from GIBCO-BRL (Gaithersburg, MD).
Iodination of Annexin V with IODO-BEADS
Na125I was neutralized before use with an equivalent volume of 3 μmol/L phosphoric acid. Annexin V was iodinated as described by Lahorte et al. (9). Two beads were washed with phosphate-buffered saline (PBS) and dried on filter paper. They were added to 18.5 MBq Na125I in 30 μL PBS for 5 min, when 50 μg annexin V were added in 100 μL of 0.1 mol/L phosphate (pH 6.5). The reaction was allowed to proceed for 10 min, the IODO-BEADS were removed, and the annexin was purified on an NAP-5 column (Amersham Pharmacia Biotech, Piscataway, NJ).
Iodination with IODO-GEN
Fifty micrograms of annexin V were dissolved in 1 mL of 0.1 mol/L phosphate buffer (pH 7.4) and reacted with 37 MBq Na125I in a V-vial (2-mL capacity; Pierce), the inner surface of which was coated with 100 μg 1,3,4,6-tetrachloro-3α6α-diphenylglycouril (IODO-GEN; Pierce). After gentle agitation of the vial for 15 min, the solution was removed from the vial, the vial was rinsed with 100 mL phosphate buffer, and the total solution was passed through a Millipore vented filter containing 1 g of Dowex 1 × 8, 100–200 mesh, chloride (Bio-Rad, Hercules, CA).
Iodination with Water-Soluble Bolton-Hunter Reagent
The reaction was performed in a glass vial because labeled annexin would adhere to the walls of polypropylene tubes. Water-soluble Bolton-Hunter reagent was dissolved in dimethyl sulfoxide at 1.5 mmol/L (0.55 mg/mL). Ten microliters of chloramine-T (5 mg/mL in PBS) were added to Na125I (37 MBq) neutralized as above, in 10 μL PBS, followed by 1 μL water-soluble Bolton-Hunter reagent. The reaction was allowed to proceed for 1 min, stopped by the addition of 10 μL sodium metabisulfite (12 mg/mL in PBS), and immediate addition of the annexin (50 μg protein in 0.5 mol/L borate [pH 9.2]). After 1 h the reaction was stopped by the addition of 300 μL of 0.2 mol/L glycine in 0.2 mol/L borate (pH 8.0). Annexin V was purified by spinning repeatedly through a Microcon concentrator (Millipore, Bedford, MA). The same procedure was used to label BSA with 131I, except that after labeling the BSA was dialyzed overnight against 0.5% BSA in PBS.
Affinity Purification of Active Annexin V
For all in vivo experiments, iodinated annexin V was further purified as follows. PS was immobilized in polyacrylamide as described by Uchida and Filburn (11). Five milligrams PS and 25 mg cholesterol were dissolved in chloroform in a glass scintillation vial. The chloroform was evaporated under N2, and 0.5 mL ethanol was added. The vial was placed in boiling water and swirled to disperse the lipids. Five milliliters of 15% acrylamide/5% bisacrylamide solution were added. This was added to a glass test tube and polymerized with 50 μL ammonium persulfate (140 mg/mL) and 2.5 μL N,N,N′,N′-tetramethylethylenediamine (TEMED). The glass tube was broken, the gel was washed with deionized H2O, minced with a razor blade, and homogenized in 20–30 mL water. The acrylamide was allowed to settle, the water was removed, and the homogenization was repeated.
Two hundred microliters of PS-polyacrylamide (equivalent to approximately 100 μg of PS) were loaded into a 2-mL column(Bio-Rad). The PS-polyacrylamide was washed 3 times with 1 mL of 10 mmol/L Tris (pH 7.5), 2 mmol/L CaCl2, and 0.5% BSA, and the annexin was added in the same buffer. The first run-through was reapplied to the column, and the column was washed 4 times with 1 mL of 10 mmol/L Tris (pH 7.5), 2 mmol/L CaCl2, and 0.5% BSA. Bound annexin was eluted by washing the column 3 times with 0.33 mL of 10 mmol/L Tris (pH 7.5), 5 mmol/L ethylenediaminetetraacetic acid (EDTA), and 0.5% BSA. The annexin released was separated from EDTA using Microcon concentrators.
Binding of Annexin V In Vitro
Human neuroblastoma IMR32 cells were irradiated with 5 Gy. Sixteen hours later, the detached cells were collected. This population was found to be entirely apoptotic, as judged by Hoechst staining and ultraviolet microscopy. Cells were exposed to annexin V in cell culture medium (DMEM + 10% FBS) for 30 min at room temperature in a volume of 100 μL. Cells were pelleted, and 50 μL of supernatant were removed. This binding assay was also performed using immobilized PS in place of cells. The tubes were counted on an LKB Wallac γ-counter (Amersham Pharmacia Biotech). The proportion of activity in the pellet was calculated as (AP − AS)/(AP + AS), where AP is the activity in the tube containing the pellet and half of the supernatant, and AS is the activity in the tube containing supernatant alone. Reactions were conducted using 105 and 3 × 105 cells per tube or 10 and 30 μg PS to ensure that binding fractions below unity were not caused by saturation of binding sites.
Animal Procedures
C3H mice were purchased from Jackson Laboratories (Bar Harbor, ME) and housed in Memorial Sloan Kettering’s animal facility. Annexin V was injected via the tail vein. Before scanning, the mice were anesthetized with ketamine/xylazine (15 μg/g and 10 μg/g, respectively) injected intraperitoneally. Whole-body irradiations (5 Gy) were performed on a Shepherd irradiator (JL Shepherd, Glendale, CA) with a 137Cs source and a dose rate of 2.5 Gy/min. Anti-FAS antibody was injected via the tail vein at 5 μg per mouse.
Imaging and Counting Activity
The mice were imaged on a single face of a Vertex gamma camera (ADAC Laboratories, Milpitas, CA) with the collimator removed. The grids used in this study were of a 6:1 ratio, 24 lines per centimeter with a focal range of 75 cm (but effectively unfocused when used under our geometry). The grids were placed in direct contact with the NaI detector and the mice were arranged directly on the scatter grids, with only a thin chuck laid between to prevent grid and crystal contamination. The use of scatter grids in place of a low-energy, high-resolution collimator was to increase sensitivity, enabling us to image mice with much lower activities of 125I, within the constraints imposed by the duration (approximately 20–30 min) of ketamine/xylazine anesthetic. The crossed steel mesh serves to provide collimation for the 25- to 35.5-keV x-ray and γ-ray photons emitted by 125I. The absence of collimation results in an image from which the distribution of the 125I within the mouse cannot be determined.
The anesthetized mice were placed prone with limbs extended directly on x-ray grids on the camera crystal face. Scans were performed for 20 min with a 50% energy window centered at 28 keV to capture x-rays and γ-rays in the energy range of 21–35 keV.
After the mice were killed, organs were removed and the radioactivity was counted. In experiments in which the animals had been co-injected with 125I and 131I, the 2-window counting technique was used. Counts in the 125I window for each organ were corrected for downscatter from the 131I activity. The counts per organ were also corrected for the injected activity of each isotope, taken as activity in the syringe before injection minus activity in the syringe after injection.
Assessment of Apoptosis
Upon killing the mice, spleens were removed, mounted in OCT medium (Sakura Fine Tek, Torrance, Ca), frozen, and sectioned. The terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay was performed as follows. Sections were fixed in 3.7% formaldehyde at room temperature for 30 min and then washed in PBS for 3 × 10 min. Endogenous peroxidase was blocked by soaking in ice-cold methanol with 0.3% H2O2 for 30 min. Sections were washed again in PBS and permeabilized with 0.1% Triton X-100 (Sigma) in 0.1% citrate for 2 min on ice. The sections were washed with PBS, and avidin-binding sites were blocked using an avidin/biotin blocking kit according to the manufacturer’s protocol. For labeling, sections were exposed to 50 μL of reaction mix: 5 U terminal transferase in reaction buffer with 2.5 mmol/L CoCl2 and 1 μmol/L biotin-16-dUTP. Sections were incubated for 1 h at 37°C and washed with PBS. Sections were exposed to an avidin-biotin-horseradish peroxidase complex for 30 min, washed with PBS, and exposed to diaminobenzidine and NiCl2 stain.
RESULTS
In Vivo Stability of 125I-Annexin V Labeled with IODO-BEADS
Annexin V was labeled using the IODO-BEADS method. In 3 separate experiments, an average of 21% of the original activity was incorporated into the annexin V. Approximately 50% of the labeled activity bound to apoptotic IMR32 cells in vitro. Six mice were each injected with approximately 330 kBq of IODO-BEADS-labeled annexin V. Three of the mice were given iodine in their drinking water for 3 d earlier to block the thyroid. Figure 1 shows images of the mice obtained 2 h after injection with the labeled annexin V. In the untreated mice, a significant fraction of the activity appears in the neck area. Thyroid blocking abolishes this uptake, which suggests that annexin V has been deiodinated in vivo. One mouse (Fig. 1, top row, center) has much lower activity than the others. The reasons for this are unknown but presumably indicate more rapid excretion.

Mice were injected with approximately 740 kBq 125I-annexin V labeled using IODO-BEADS. (Top) Mice were given KI in drinking water for 3 d before injection. (Bottom) Mice received no added iodide in diet.
Annexin V was also labeled using IODO-GEN; like IODO-BEADS, this technique results in the direct addition of iodine to tyrosine residues (12). Two hours after injection of this preparation into mice, we observed accumulation of activity in the thyroid region, suggesting that this preparation was equally prone to deiodination in vivo. (Two independent batches were injected into a total of 5 mice [data not shown].)
Iodination of Annexin V with Water-Soluble Bolton-Hunter Reagent
Annexin V was labeled with iodine in a 2-step reaction. The iodine was reacted with the Bolton-Hunter reagent, followed by addition of the iodinated reagent to annexin V. Although 125I-labeled Bolton-Hunter reagent is commercially available, our aim was to devise a method that could be applied to 124I.
The reaction was optimized for pH and temperature. Figure 2 shows their effect on labeling efficiency and annexin V activity. The efficiency of the labeling reaction is seen to increase with increasing pH. On the basis of 3 independent experiments, at pH 8.1 the incorporation is 0.26, rising to 0.35 at pH 8.6 and to 0.41 at pH 9.2. Using a 2-tailed t test, these differences are all statistically significant (pH 8.1 vs. pH 8.6, P = 0.015; pH 8.6 vs. pH 9.2, P = 0.012). The binding activity of the labeled product to PS is 0.8 over this pH range.
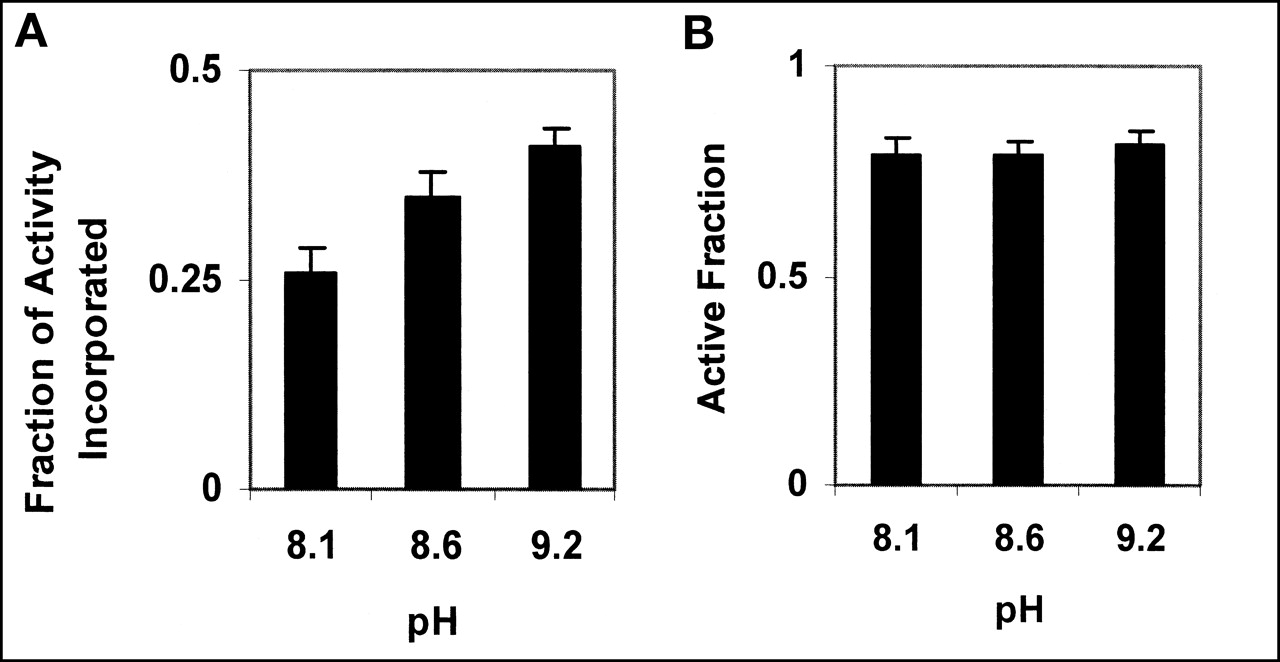
(A) Activity incorporated in annexin V increases with increasing pH. (B) Activity of annexin V, as measured by its ability to bind to PS, is unaffected by pH of labeling reaction. Error bars represent 95% confidence intervals, based on 3 independent experiments.
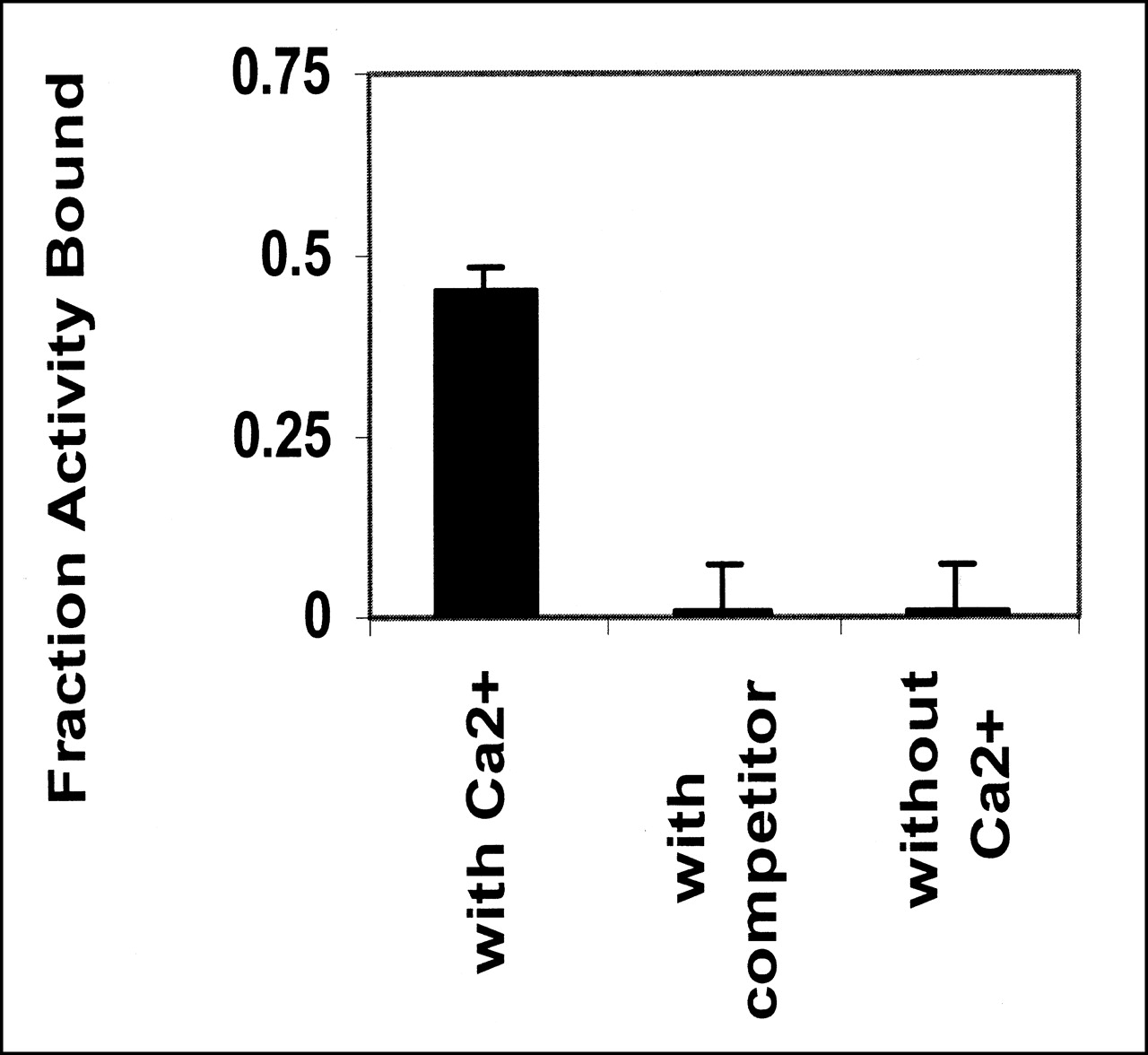
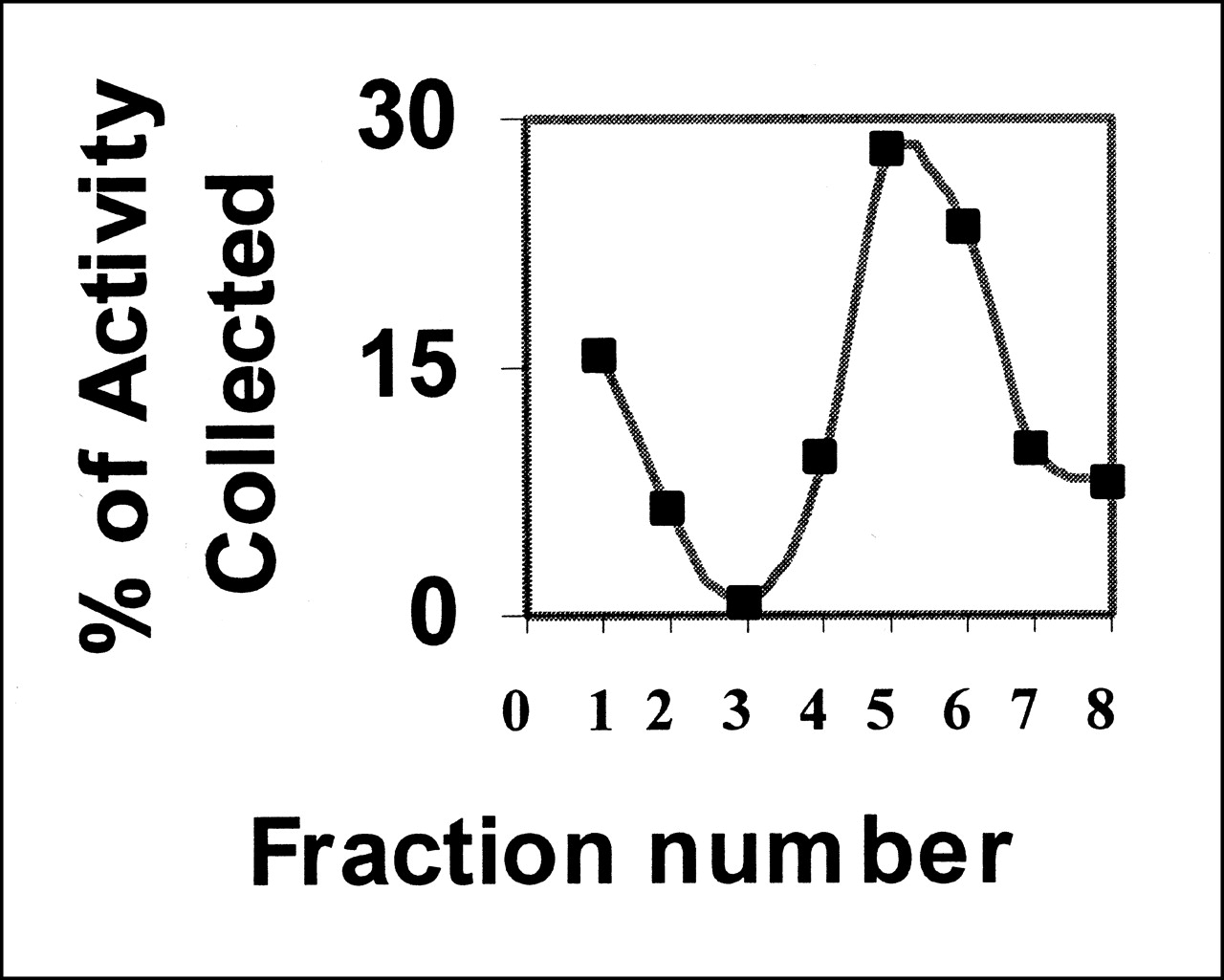
Binding of annexin V is calcium dependent (13). The preparation does not bind to cells in the absence of calcium, and binding can be abolished by an excess of unlabeled competitor (Fig. 3). The active fraction of annexin V was always <1. Further purification of annexin V can be obtained by adding the preparation to a PS-polyacrylamide column. Active annexin V will bind to the column in the presence of calcium, whereas inactive protein will pass through. Elution with buffer containing EDTA will release the bound annexin V (Fig. 4). The fraction that failed to bind in the presence of Ca2+ showed no ability to bind apoptotic cells (data not shown).
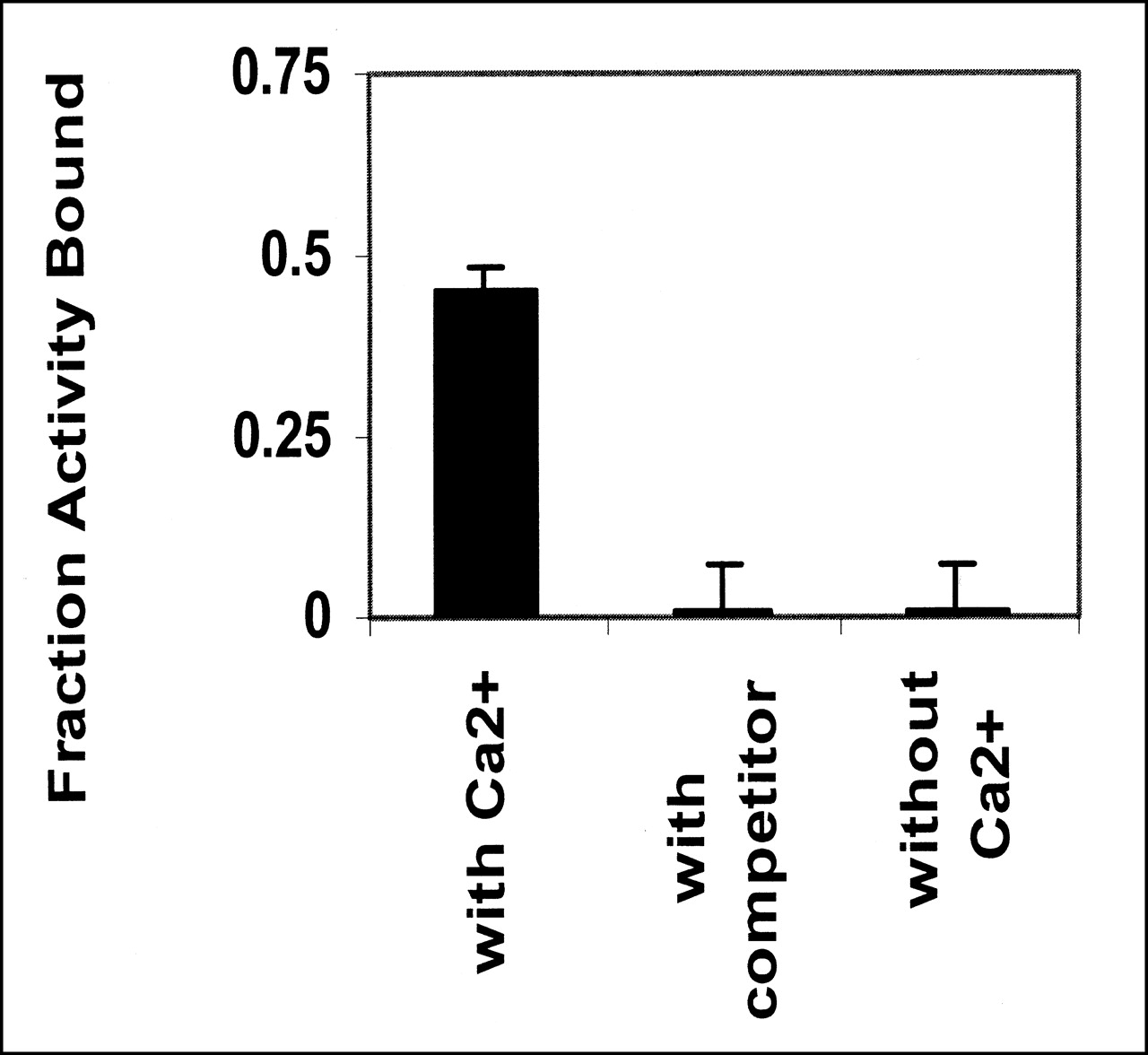
Bolton-Hunter-labeled annexin V binds to apoptotic cells only in presence of Ca2+. Binding is abolished by addition of unlabeled annexin at 100-fold excess. Data are from 2 independent experiments.
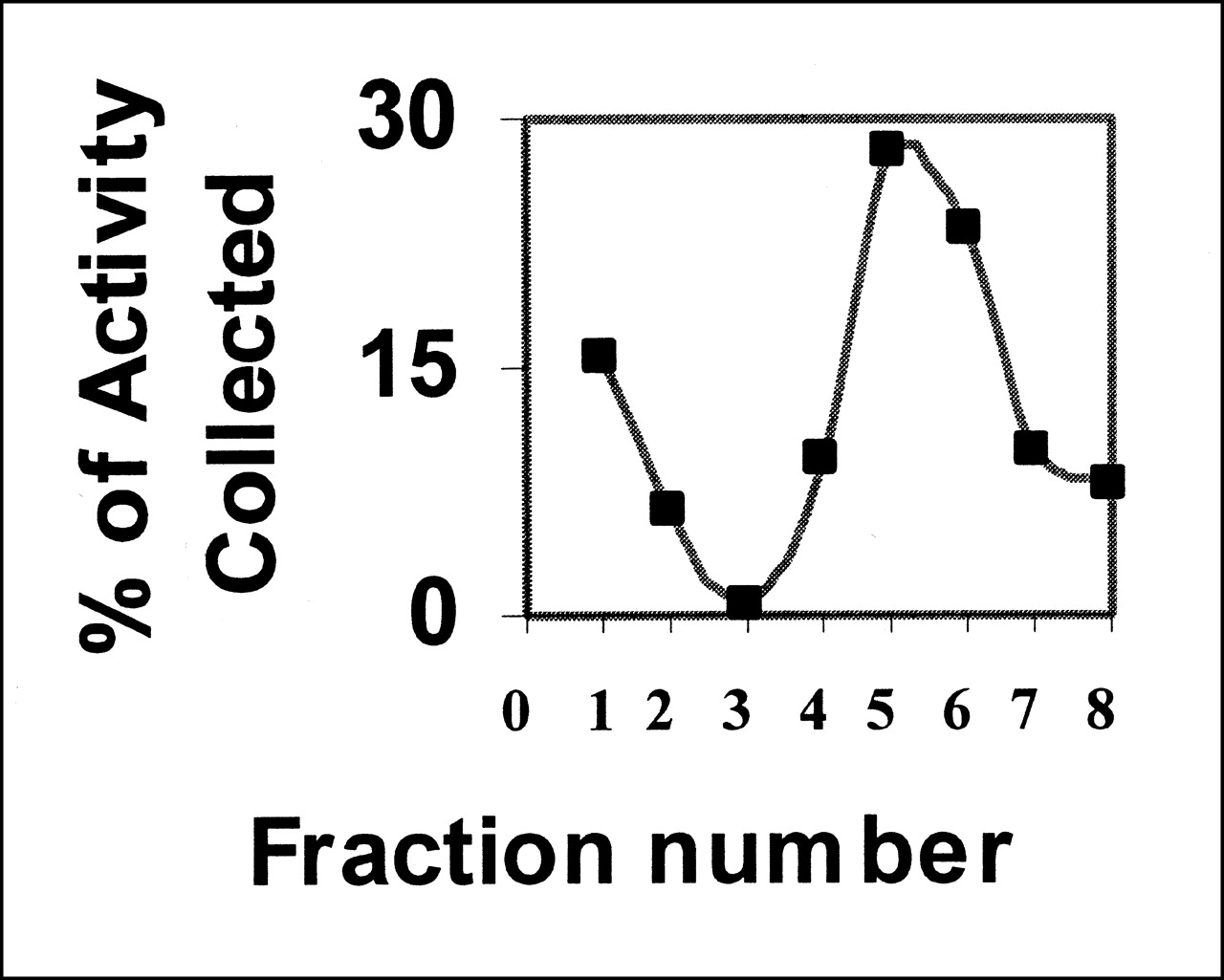
Biologically active fraction of Bolton-Hunter-iodinated annexin V can be purified by affinity chromatography. Labeled annexin was loaded onto column in calcium-containing buffer; this buffer was used to elute unbound activity (fractions 1–3). Column was washed with buffer containing EDTA to elute bound activity (fractions 4–8).
In Vivo Application of Bolton-Hunter-Iodinated Annexin V
To show that Bolton-Hunter-labeled annexin V is active in vivo, mice were exposed to 5-Gy irradiation (whole body) and injected 6 h later with 125I-annexin. 131I-BSA, also labeled with the Bolton-Hunter reagent, was co-injected to serve as an internal standard. Two hours later the mice were killed and radioactivity was counted in the spleen, liver, and kidney. The results are shown in Figure 5A and 5B. There was a 4-fold increase in 125I uptake in the spleens of the irradiated mice, with P = 0.004 (2-tailed t test), implying annexin binding. The spleens showed significant TUNEL staining relative to that of the control animals (Fig. 5B).
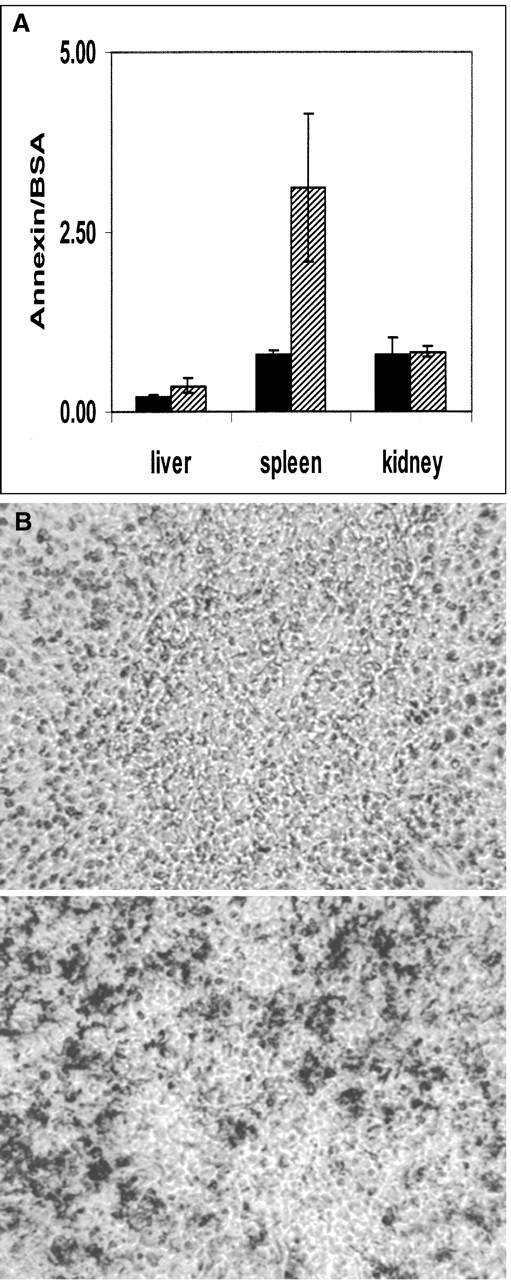
(A) 125I-Annexin V binding in 4 untreated animals (black bars) compared with irradiated animals (hatched bars). Annexin V uptake is expressed relative to 131I-BSA uptake. Error bars represent 95% confidence intervals. (B) TUNEL staining of spleen sections: (Top) control; (Bottom) irradiated.
To image annexin V binding, mice were co-injected with Bolton-Hunter-labeled 125I-annexin V and 5 μg of anti-FAS antibody to induce liver apoptosis. The results are shown in Figure 6A and 6B. Figure 6A shows the control animals; the mice in Figure 6B received the anti-FAS antibody. The anti-FAS treatment led to a 5-fold increase in the 125I activity in the liver (calculated as counts/min/g of tissue/kBq injected); this difference was highly significant (P = 0.003; 2-tailed t test). Over a period of 2 h, the mice did not show any evidence of activity accumulating in the thyroid. Figure 6C shows 125I activity in mice, imaged 24 h after an injection of Bolton-Hunter-labeled 125I-annexin V. At this time, the mice had excreted >95% of the injected activity. Of the retained activity, a significant fraction was found in the thyroid region, indicating that the Bolton-Hunter-labeled annexin V is eventually subject to deiodination.

Mice injected with Bolton-Hunter-labeled 125I-annexin V. (A and B) Mice were injected with 925 kBq 125I-annexin V and imaged 2 h later. (A) Control animals. (B) Mice were injected with anti-FAS antibody. (C) Mice received 370 kBq 125I-annexin V and were imaged 24 h after injection.
After killing the animals, activity was also counted in the kidneys. In the control animals, kidney uptake was 3–4 times higher per gram of tissue in the kidneys than that in the liver.
DISCUSSION
We have shown that when annexin V is iodinated on its tyrosine residues with IODO-GEN or IODO-BEADS, the label is prone to deiodination in vivo. We did not attempt iodination with chloramine-T because of the risk of protein denaturation associated with chloramine-T. Proteins iodinated with the Bolton-Hunter reagent are known to be substantially less prone to in vivo deiodination (14), and we have taken advantage of this fact to produce a compound that is not deiodinated substantially in vivo, at least over the first 2 h. During this period, the Bolton-Hunter-labeled compound can bind apoptotic tissue in vivo.
Annexin V was labeled to a specific activity of 110–220 kBq/μg, which is equivalent to 1 iodine per 10 molecules of annexin V. Annexin V and the Bolton-Hunter reagent were present at equimolar amounts (i.e., 1.5 nmol per reaction), whereas the iodine (37 MBq 125I) was present at 0.5 nmol. Increasing the iodine in the reaction could clearly raise the specific activity. Because the Bolton-Hunter reagent can accept 2 iodine atoms, the activity could be raised to 222 MBq, before a corresponding increase in Bolton-Hunter reagent would also be necessary.
The labeled annexin retains its biologic activity. We measured the binding of labeled annexin V using apoptotic cells and PS immobilized in polyacrylamide. Surprisingly, the estimate of the active fraction was always lower when the measurement was conducted on cells (50% vs. 75%), as can be seen by comparing Figures 2B and 3. We are not sure why this should be so: Possibly, in the cell preparation, the annexin V is subject to proteolytic attack or some other form of inhibition. Regardless of which substrate was used, labeled annexin V binding was dependent on Ca2+, indicating that the protein had retained its biologic activity. We were able to further enhance the active fraction of the labeled protein by applying it to a column containing PS.
The labeled annexin was active in vivo. After irradiation, we found a 4-fold increase in 125I uptake in the spleen relative to that of the control animals. We believe that this finding was associated with annexin V binding apoptotic cells because the TUNEL assay showed the induction of apoptosis in the irradiated spleen. No increase in the BSA uptake in the irradiated spleen was found, ruling out the possibility that increased annexin V uptake was caused by some nonspecific change in protein retention in the irradiated animals.
99mTc-Annexin V localizes in the kidneys of control animals, to such an extent that the kidneys are clearly visible on scans, with minimal activity elsewhere (7). With Bolton-Hunter-labeled annexin V, we did not observe such clear-cut localization to the kidneys (Fig. 6). Instead, a more diffuse pattern of uptake in the visceral region was found. We measured uptake for liver and kidney in terms of activity per gram. Although we found approximately 4 times more activity in the kidney than in the liver, with 99mTc-labeled annexin V, the ratio of kidney-to-liver activity was >10 (7). These differences may be associated with the different linkers used. The Bolton-Hunter reagent will tend to make the complex quite lipid soluble, leading to higher liver uptake and, subsequently, increased gut activity. In contrast, the hydrazinonicotinamide linker used by Blankenberg et al (7) is less lipid soluble and has been shown to significantly reduce liver uptake compared with that of other 99mTc linkers (15).
CONCLUSION
Bolton-Hunter chemistry represents a good way to iodinate annexin V and may prove useful in adding 124I to annexin V for use as a PET tracer. However, its tendency to accumulate in the liver may restrict its use for measuring apoptosis in tumor deposits within the liver.
Acknowledgments
The authors thank Dr. Pat Zanzonico for his thoughtful discussion and advice.
Footnotes
Received Aug. 27, 2001; revision accepted Jan. 29, 2002.
For correspondence contact: James Russell, PhD, Department of Medical Physics, Memorial Sloan Kettering Cancer Center, S1136, 1275 York Ave., New York, NY 10021.
E-mail russellj{at}mskcc.org




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}