


Abstract
It was reported recently that 99mTc-hexakis-2-methoxyisobutyl isonitrile (MIBI) uptake is drastically reduced in cancer cells that express the multidrug resistance (MDR) product, Pgp 170 kDa (Pgp), suggesting that 99mTc-MlBI is a transport substrate for this transmembrane glycoprotein. In our study, we explored if another pump, a multidrug resistance-associated protein (MRP), could affect 99mTc-MIBI uptake. In addition, we studied the involvement of intracellular glutathione (GSH) as a modulator of 99mTc-MIBI uptake by both Pgp and MRP proteins. Methods: MDR1 and MRP gene expression in seven human tumor cell lines was determined on a transcriptional level by reverse transcriptase polymerase chain reaction and on a protein level using immunocytochemistry. Technetium-99m-MlBl uptake was quantified by measuring radioactivity retained in the cells incubated at 37°C in the presence or absence of buthionine sulfoximine (BSO), which depletes cellular GSH. The cellular GSH content was determined with Ellman’s reagent. Results: Cell lines were classified according to their phenotypic characteristics: 1/MRP-/Pgp-: breast cancer cells (MCF7), lung carcinoma cells (H69S) and mouth epidermoid tumor cells (KB 3.1), 2/MRP-/Pgp+: MCF7 mdr+, KBA.1; and 3/MRP+/Pgp-: small-cell lung carcinoma (H69 AR and A 549). Technetium-99m-MIBI uptake was significantly lower in cells expressing MRP as well as Pgp compared to MRP/Pgp cells. Depletion of GSH by BSO resulted in an increase of 99mTc-MIBI uptake in multidrug resistant cells over-expressing MRP but not expressing Pgp. Conclusion: Technetium-99m-MIBI is extruded by both Pgp and MRP efflux pumps. However, MRP action is indirect and involves intracellular GSH for a presumed interaction with the 99mTc-MIBI before its efflux. Technetium-99m-MIBI seems to be a good candidate for a noninvasive marker to diagnose MDR1 related to Pgp and MRP expression in tumors of different origin.
- technetium-99m-hexakis-2-methoxyisobutyl isonitrile
- multidrug resistance
- glutathione
- human cancer cell lines
- buthionine sulfoximine
Multidrug resistance (MDR) is the major barrier to efficient chemotherapy of cancer. Some malignancies including non-small cell lung and colon cancer have a resistant phenotype by definition and respond poorly to cytotoxic drugs. Other tumors, like advanced breast and ovarian cancers, frequently relapse after an initial response and become resistant to a broad spectrum of drugs (1).
Causes of chemotherapy failure are multifactorial including the physical inability of the drugs to reach malignant cells because of poor tumor vascularization and diverse cellular mechanisms of resistance lowering intratumoral drug concentration, or altering the ability of these drugs to affect their targets (2). Different cellular mechanisms of MDR have been identified. One of them involves the MDR gene coding for Pgp 170 kDa (Pgp), a transmembrane protein transporter that operates as a drug efflux adenosine triphosphate-dependent pump (3,4). Another transmembrane glycoprotein, multidrug resistance-associated protein (MRP), 190 kDa, was recently characterized as a drug exporter belonging to the ATP-binding superfamily (5,6).
An increasing number of biochemical and molecular alterations also have been described in cell lines resistant to multiple cytotoxic drugs. Among the increased detoxifying enzymes in these cells are the glutathione S-transferases, which catalyze the conjugation of electrophilic hydrophobic compounds to the thiol glutathione (GSH), and glutathione peroxidase, a second GSH-dependent enzyme, which catalyzes the detoxification of hydroperoxides (7). It was proposed that a decrease of topoisomerase II activity and alteration in GSH level can cause resistance to cytotoxic drugs (8,9). The tripeptide GSH is implicated in the detoxification of a wide range of xenobiotics including many of the currently used cytotoxic drugs (10). Modulation of GSH level alters the response of cells to numerous cytotoxic drugs and to ionizing radiation. Recently, it was reported that anionic and amphiphilic glutathione S-conjugates are exported by MRP (11).
Technetium-99m-hexakis-2-methoxyisobutyl isonitrile (MIBI) is a member of the isonitrile class of coordination compounds (12) and a lipophilic cation used for myocardical perfusion imaging (13) that has been reported to be recognized by Pgp. Studies were performed in vitro on Chinese hamster resistant cells (14) and on numerous human cancer cell lines (15–17).
The aim of our study was to explore if 99mTc-MIBI could be recognized not only by Pgp but also by MRP and to assess the possible role of cellular GSH in these two efflux pump functions. The preliminary results from our laboratory were presented recently (18).
MATERIALS AND METHODS
Tracer Preparation
Technetium-99m-MIBI was prepared according to the manufacturer’s instructions (DuPont Pharma SA, Les Ulis, France). We used generator equilibrium equations to calculate the absolute concentration of total tracer (radioactive and cold) in the solutions (15). Molarity was expressed in terms of total technetium-MIBI (99 + 99m). Radiochemical purity was always greater than 96%. To obtain 1 nM solutions, 99mTc-MIBI was diluted in RPMI medium (Gibco, Grand Island, NY) or Dulbecco’s modified Eagle’s medium (DMEM, Gibco) (15).
Cell Line Culture
Seven human cell lines were studied (Table 1). Cells were grown in tissue culture flasks (Falcon, Beckton Dickinson, Lincoln Park, NJ) in DMEM supplemented with 2 mM L-glutamine (Gibco, Grand Island, NY), penicillin (50 IU/ml; Gibco, Grand Island, NY), streptomycin (50 μg/ml; Gibco, Grand Island, NY) and 10% fetal calf serum (Biological Industrie Kibbutz, Beth Haemek, Israel). MCF7mdr+ and H69 AR cells were cultured in RPMI supplemented with doxorubicin (Farmitalia Carlo, Erba, Italy) 5.8 and 0.5 μg/ml, respectively. For KBA. l culture, the DMEM supplemented with doxorubicin (1 μg/ml) was used.
Phenotype of Studied Human Cancer Cell Lines
Technetium-99m-MIBI Uptake
Technetium-99m-MIBI uptake was determined, as previously described (15), in cells pretreated, or not, with an intracellular glutathione-depleting agent, 25 μM buthionine sulfoximine (BSO) for 24 hr. Briefly, adherent cells were harvested with 0.05% trypsine (Gibco, Grand Island, NY) and washed twice with RPMI or DMEM medium. The cells were counted in a hemocytometer (Coultronics, Margency, France) and suspended in the medium at a concentration of 1 × 107 cells/ml. To minimize nonspecific binding of 99mTc-MlBI to plastic tubes, we presaturated them for 1 hr with a phosphate-buffered saline (PBS) solution containing 1% of fetal calf scrum followed by three washes with PBS. Technetium-99m-MIBl uptake was initiated by adding an equal volume of tracer solution to the cell suspension. At various time intervals, 50-μl aliquots were transferred to 400-μl microfuge tubes (Beckman Instruments, Palo Alto, CA) containing 200 μ1 of fetal calf scrum and centrifuged for 1 min in an Eppendorf microfuge (15,000 × g). After freezing the tubes, their tips were cut off and the pellet and the supernatant were separated. Radioactivity was determined in a gamma counter. Nonspecific binding was determined with the same method but without cells. The results were expressed as an accumulation ratio calculated by dividing the total specific cell uptake (without nonspecific binding) by the total radioactivity (pellet and supernatant). All datapoints represented the mean of at least two independant experiments performed in triplicate. The ratio of tracer uptake in the cells treated and not treated with BSO was named the multiplying factor.
Reverse Transcriptase Polymerase Chain Reaction
Total ribonucleic acid (RNA) was extracted from cells treated, or not, with 25 μM BSO using a mRNA isolation kit (RNA plus, Bioprobe, Montreuil, France) according to the procedure of Chomczynski and Sacchi (19). Synthesis of cDNA was performed with 0.25 U/μl of reverse transcriptase (Promega, Woods Hollow Road, Wl). This cDNA was used as a template for potymerase chain reaction (PCR) amplifications using 0.25U Taq polymerase (ATGC, Noisy le Grand, France). Thirty-five cycles were performed on a Perkin-Elmer thermocycler at 94°C, 50°C (positive control, β2-microglobulin) or 55°C (PgP and MRP) for 1 min each, at 72°C for 1.5 min with a final elongation step at 72°C for 10 min. Forward and reverse oligonucleotide primer (Genset, Paris, France) nomenclature and sequences are listed in Table 2. Fifteen micro-liters of each PCR-amplified mixture were used for electrophoretic analysis of PCR products on 3% agarose gel with DNA size markers and stained with ethidium bromide (BET). The BET staining, revealed in ultraviolet, was quantified by an image analyser (NIH Image 1.44 Macintosh, Paris, France). The results are presented as ratio of MDR or MRP and β2-microglobulin signals.
Position and Sequence of Sense* and Antisense† Oligonucleotides Used as Primers in Poymerase Chain Reaction Amplification
Detection of Pgp170 and MRP by Immunocytochemistry
Cytocentrifuged preparations of tumor cell lines were air dried 1 hr, fixed in acetone for 10 min at room temperature and frozen (−80°C). For every staining, a positive control was used (MCF7 mdr+ for Pgp170 and H69AR for MRP). All washes were performed in PBS. Negative controls for each sample were performed as described below but without primary antibody.
Pgp Detection
Cytocentrifuged preparations were incubated 30 min at room temperature with the monoclonal antibody (MAb) UIC2, mouse IgG2a (Immunotech, Marseille, France) at a final concentration of 2 μg/ml. The MAb was diluted in 0.05 M Tris (pH 7.4) containing 1% bovine serum albumin and 5% (v/v) goat serum (Dako, Carpinteria, CA). MAb binding was detected using rabbit biotinylated antimouse immunoglobulins in PBS (15 min of incubation) and streptavidin conjugated to horseradish peroxidase in PBS (15 min of incubation). Bound peroxidase was revealed with amino-ethyl-carbazole and H2O2 in N, N-dimethylformamide. Then the slides were counterstained with hematoxilin.
MRP Detection
We have used the MON9018 antibody (rat IgG2a) (Genzyme, Paris, France) in dilution of 1:20. The slides were incubated 1 hr at room temperature. MAb binding was detected using a rabbit peroxidase-conjugated antirat antibody in a dilution of 1:50. The incubation was performed for 1 hr at room temperature. Bound peroxidase was developed with diaminoben-zidin and H2O2 in Tris. Then, the slides were counterstained with hematoxilin.
When more than 10% of cells were positive, the tumor cell line was considered strongly positive (+ +). When less than 10% of the cells were positive, the tumor cell line was considered weakly positive (+). When no cells were positive, the tumor cells were designated negative (-).
Determination of Reduced Glutathione Content in Cells
To deplete the level of intracellular GSH, the cells were treated with 25 μM BSO for 24 hr. The viability of cells was determined using trypan blue staining after every BSO treatment. When more than 90% of cells were viable, the experiment was maintained. It is noteworthy that 50 μM BSO induced the death of more than 50% of the treated cells. Proteins from cells treated, or not, with BSO (2–8 106 per sample) were precipitated with 30% sulfosalycilic acid. After centrifugation, the cellular reduced GSH content was determined in the supernatant with Ellman’s reagent, 5,5-dithiobis (2-nitrobenzoic acid) and absorbance was measured at 412 nm (20). The results are expressed as μmol GSH/106 cells.
Statistics
Values are presented as mean ± s.d. Statistical significance was determined by unpaired two-tailed Student’s t-test; p < 0.05 considered significant.
RESULTS
Immunohistochemical and Reverse Transcriptase Polymerase Chain Reaction Analysis of MDR and MRP Expression
All cell lines were assessed to estimate the level of MDR1 and MRP expression on mRNA and protein level. Immunohistochemical (Table 3) and reverse transcriptase polymerase chain reaction (RT-PCR) (Table 4) results concordantly showed that only two cell lines expressed Pgp. RT-PCR indicated that KBA. l cells produced more Pgp mRNA than MCF7mdr+ cells. Expression of MRP was observed in all studied cell lines using RT-PCR, but only two lines, A549 and H69AR, were positive when immunohistochemical analysis was applied. The highest level of MRP was found in H69AR cells as determined by two methods.
Results of Immunocytochemical AnaIysis*
RT-PCR Analysis of Multidrug Resistance and Multidrug Resistance Associated Protein Expression in Human Cancer Cell Lines
Lack of Technetium-99m-MIBI Uptake Related to Pgp and MRP Expression
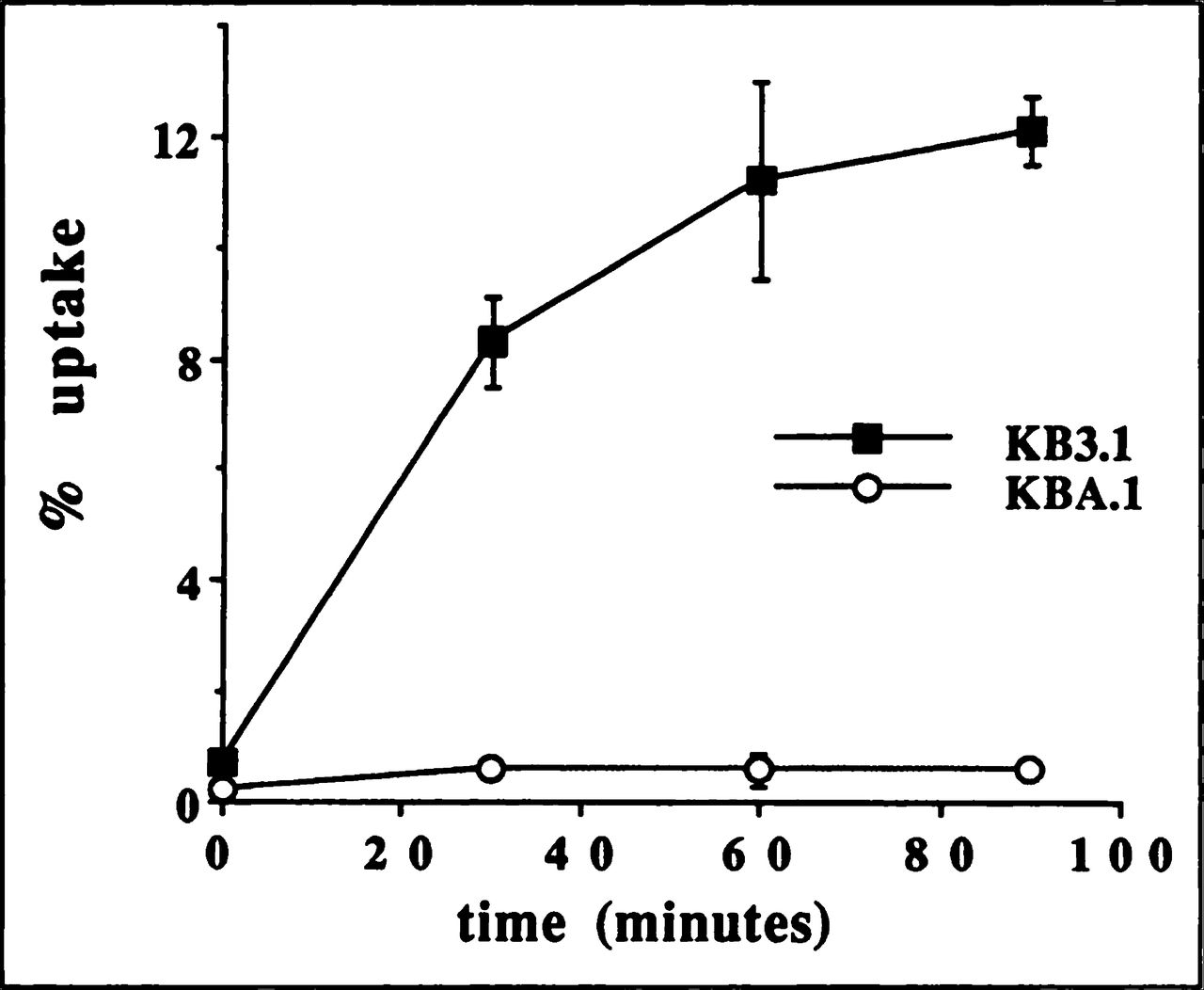
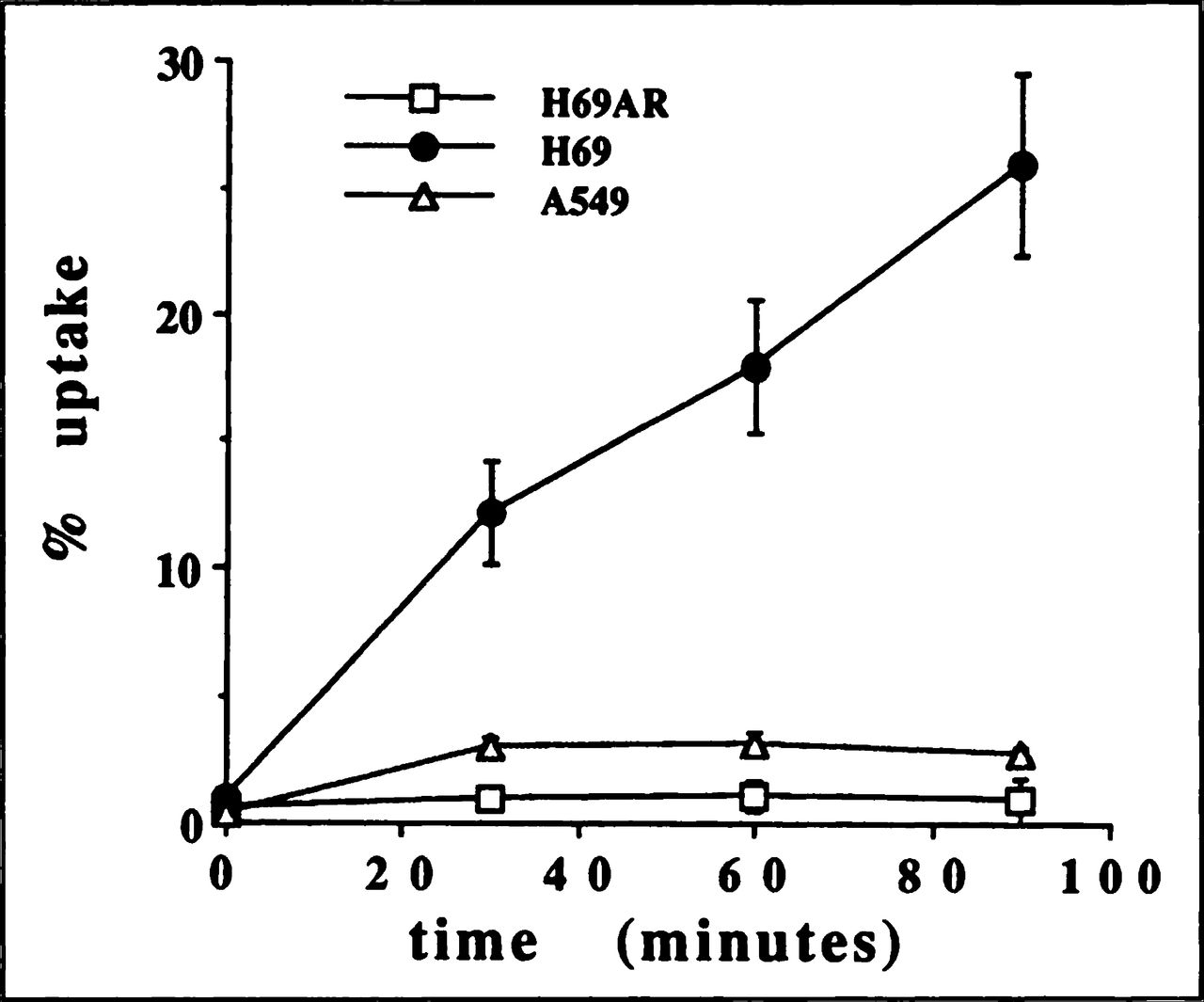
The kinetics of 99mTc-MIBI uptake for nonresistant cell lines KB3.1 (Fig. 1) and H69 (Fig. 2) were similar to the curve observed for MCF7 previously published by our laboratory (15). Thus, the tracer is retained not only in breast cancer cells but also in mouth and lung cancer cells. The expression of Pgp in KBA. 1 and MRP in A549 and H69AR drastically decreased the 99mTc-MIBI uptake (Figs. 1 and 2) as we reported for MCF7mdr+ (15). After 90 min of incubation, the tracer uptake was significantly (p < 0.05) higher (2.8%) in A549 cells that weakly express MRP than H69AR (0.7%) producing more MRP (Figs. 2 and 4). There was no difference in tracer uptake in MCF7mdr+ and KBA. 1 cells. It is possible that Pgp expression was high enough in both cell lines to completely suppress tracer accumulation.

Time-course of 99mTc-MIBI uptake in human mouth carcinoma cell lines: sensitive KB3.1 (■) and resistant (MDR1 +) KBA.1 (○).

Time-course of 99mTc-MIBI uptake in human small-cell lung cancer cell lines: a sensitive H69 (●) and resistant (MRP+) H69 AR (□) and A549 (∆).
Effect of Glutathione Depletion on Technetium-99m-MIBI Uptake
Concerning the basal GSH level in cells, we have not found a significant systemic tendency between sensitive and resistant cells (Table 5). In all studied cell lines, sensitive and resistant, BSO decreased the GSH level by at least 75% as compared to the basal level (Table 5).
Glutathione Concentration in Cancer Cells
Depletion of intracellular GSH resulted in a highly significant (p < 0.001) increase of 99mTc-MIBI uptake in MRP but not Pgp170-expressing cancer cell lines (Fig. 4). In two of three studied sensitive cell lines, both epithelial, GSH depletion resulted in weaker but significant (p < 0.05) augmentation of tracer accumulation (Fig. 3). No alteration of 99mTc-MIBI uptake by BSO was observed in small-cell lung carcinoma H69 (Fig. 3). Interestingly, BSO treatment increased the MRP expression (Table 4) in sensitive as well as in all resistant cells.

Technetium-99m-MIBI uptake after 90 min incubation in sensitive cancer cell lines pretreated (■), or not, (□) with 25 μM BSO for 24 hr.

Technetium-99m-MIBI uptake after 90 min incubation in resistant cancer cell lines pretreated (■), or not, (□) with 25 μM BSO for 24 hr.
DISCUSSION
In our study, we showed that human cancer cells of different origins are able to accumulate 99mTc-MIBI. Development of MDR related not only to Pgp (14,15,25) but also to MRP led to a sharp decrease of 99mTc-MIBI uptake. These observations agree with recent reports from Crankshaw and Piwnica-Worms (26), de Vries’ et al. (27) and Franssen’s laboratories (28) and suggest that 99mTc-MIBI may be a transport substrate of both Pgp and MRP. However, the mechanisms of 99mTc-MIBI efflux by these two pumps seem to be different. Indeed, BSO depletion of intracellular GSH content in cells overexpressing MRP but not in Pgp-positive cells restored 99mTc-MIBI accumulation. This phenomenon was described recently for another lung carcinoma cell line GCL4/ADR (28). We found that exposition of sensitive epithelial cells (MRP, Pgp) to BSO increased the amount of 99mTc-MIBI retained in the cells, which argues for other possible side effects of BSO or/and GSH depletion in MCF7 and KB3.1 cells. As sensitive lung carcinoma cells H69 were not affected by BSO treatment, it also is probable that these differences between uptake in sensitive cell lines may reflect tissue-specific variations in complex GSH-producing and GSH-using pathways (29). Finally, this phenomenon could be due to the depletion of GSH-decreasing protection of the plasma membrane. This low protection might be an oxidative stress damaging the membrane and resulting in an increased drug influx, a reversal of resistance and a greater intracellular concentration of 99mTc-MIBI. Cellular GSH depletion to 20%–30% of total glutathione concentration can impair the cell’s defense against the toxic actions of such compounds and may lead to cell injury and death (30). However, the last hypothesis seems to be less probable as in our experimental conditions the majority of cells remained viable. We also observed that exposure of sensitive as well as resistant cells to BSO increased the MRP gene expression. Recently, it was reported that the depletion of GSH as a result of BSO treatment alters the binding of transcriptional factors, including AP-1 and NFkB to DNA (31), suggesting the possibility of regulation of gene expression.
Determination of reduced GSH is important for our study. Intracellular concentration of GSH is maintained mainly as reduced GSH due to thiol redox status produced by intracellular GSH reductase and the reduced form of nicotinamide-adenine dinucleotide phosphate. Detoxification or inactivation of electrophilic alkylating agents and platinum compounds may occur as a consequence of their reactions with thiol-containing compounds, such as GSH and metallothionine. For example, GSH conjugates are formed with a variety of alkylating agents in nonenzymatic and GST-catalyzed reactions. Associations have been reported between increased GST levels or specific GST isoenzymes and resistance to nitrosureas, chlorambucil, and other nitrogen mustards. Increased GSH levels also have been reported with resistance to alkylating agents and cisplatin (32). However, we did not observe significant alteration in the GSH level between sensitive and resistant cells.
Positively charged, as well as neutral, drugs such as MK 571, etoposide and, presently, 99mTc-MIBl may be transported as are other negatively charged groups (11,33). Mitochondrial GSH is important in regulating inner membrane permeability by maintaining intramitochondrial thiols in the reduced state. Of the cellular content of GSH, 10%–15% is located in the mitochondria (34). MRP mediates ATP-dependent membrane transport of the endogenous glutathione conjugate leukotriene LTC 4 and of structurally related anionic conjugates of lipophilic compounds (35,36). MRP acts as a multispecific organic anion transporter as demonstrated for fluorescent calcein extruded independently of intracellular GSH concentration in a direct way (37). Further study is necessary to determine the manner of GSH involvement in MRP-dependent 99mTc-MIBI efflux from cancer cells.
CONCLUSION
Our study demonstrates that loss of 99mTc-MIBI uptake in cancer cells of different origins is related not only to Pgp but also to MRP pump expression. Thus, 99mTc-MIBI is not specific for Pgp. The mechanism of 99mTc-MIBI efflux by MRP pump is unknown, but the involvement of GSH, glucuronate or sulfate conjugates negatively charged is possible. Our data suggest that 99mTc-MIBI could be a good candidate to diagnose the presence of both Pgp, 170 kDa, and MRP, 190 kDa, in tumors by a noninvasive method in vivo and to monitor the efficiency of reversal products including BSO. The last molecule is now in clinical Phase I study. To validate the use of imaging with 99mTc-MIBI to monitor the development of resistance in vivo, it is necessary to correlate the loss of tracer accumulation in cancer tissue with the detection of Pgp and MRP expression.
Acknowledgments
This work was supported by grants from the Association pour la Recherche sur le Cancer (ARC), the Midy Foundation and Cancer Leagues 89 and 93. We are grateful to SCP Cole from CRL Kingston, Ontario, Canada for providing the H69 AR cells. We also thank Dr. G. Baillet for editorial assistance.
- © 1998 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication August 5, 1997.
- Accepted for publication October 13, 1997.
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Usefulness of 99mTc-Sestamibi Scintigraphy in Suggesting the Therapeutic Effect of Chemotherapy against Gastric Cancer
- 99mTc-MIBI Imaging as a Predictor of Therapy Response in Osteosarcoma Compared with Multidrug Resistance-Associated Protein and P-Glycoprotein Expression
- What Can We Expect from MDR Breast Cancer Imaging with Sestamibi?
- Expression of Multidrug Resistance Protein and Messenger RNA Correlate with 99mTc-MIBI Imaging in Patients with Lung Cancer
- P-Glycoprotein and Multidrug Resistance-related Protein Expressions in Relation to Technetium-99m Methoxyisobutylisonitrile Scintimammography Findings