


Abstract
A human-derived intrinsically nonimmunogenic reporter gene was tested for PET imaging of different molecular–genetic processes for potential clinical use. Methods: The human mitochondrial thymidine kinase type 2 (hTK2) reporter gene truncated at the N terminus (ΔhTK2), alone or fused with green fluorescent protein (GFP), was used for preclinical evaluation in a mouse model. The levels of enzymatic activity of ΔhTK2 and ΔhTK2 GFP proteins were assessed using radiotracer accumulation and prodrug activation assays in vitro and in subcutaneous tumors grown from the corresponding cell lines in nude mice. Kinetic analyses of 124I-2′-fluoro-2′-deoxy-1-β-d-β-arabinofuranosyl-5-iodouracil (FIAU), 18F-2′-fluoro-2′-deoxy-1-β-d-β-arabinofuranosyl-5-ethyluracil (FEAU), or 18F-9-(4-18F-fluoro-3-hydroxymethylbutyl)guanine (FHBG) uptake in tumors and biodistribution studies were performed. Results: ΔhTK2 was successfully expressed in the cytoplasm of transduced cells. A new anti-hTK2 monoclonal antibody 8G2 was developed. The levels of FIAU and FEAU accumulation in cells expressing ΔhTK2 and ΔhTK2 GFP were at least 10-fold higher than in wild-type cells in vitro and about 6 times higher in vivo. We determined that FEAU is a more specific reporter substrate for ΔhTK2 than FIAU, whereas FHBG is not phosphorylated by this enzyme. In addition, we showed that ΔhTK2 transduced cells can be eliminated by treatment with d-arabinofuranosyl-cytosine. Conclusion: We have tested a human-derived reporter gene that is likely to be nonimmunogenic and potentially allows for long-term monitoring of different molecular–genetic processes by nuclear imaging techniques in humans. Using 124I-FIAU, 18F-FIAU, or 18F-FEAU, it should be possible to image ΔhTK2 reporter gene expression with PET in preclinical and clinical studies.
Recent advances in noninvasive molecular imaging have provided new research tools for monitoring the expression of different genes and activities of various signal transduction pathways. Reporter systems have been developed for tracking transgene expression in vitro as well as in vivo, and in transgenic animals. Stable genetic cell labeling with various reporter genes has been used to circumvent the temporal limitations of in vitro radiolabeling or magnetic labeling of cells. The paradigm for imaging with reporter genes involves administration of a probe that is selectively bound or metabolized and is trapped by interaction with the gene product in the reporter gene transduced cell. The level of probe accumulation is proportional to the level of reporter gene product expressed (1).
Wild-type herpes simplex virus type 1 thymidine kinase (HSV1-tk) and mutant HSV1-sr39tk are the most widely used reporter genes for PET imaging, and the 2 most widely used radiolabeled reporter probes are 124I-2′-fluoro-2′-deoxy-1-β-d-β-arabinofuranosyl-5-iodouracil (FIAU) (with its derivative, 18F-2′-fluoro-2′-deoxy-1-β-d-β-arabinofuranosyl-5-ethyluracil [FEAU]) and 18F-9-(4-18F-fluoro-3-hydroxymethylbutyl)guanine (FHBG) (2,3). The initial clinical imaging studies reported by Jacobs et al. (4) showed successful 124I-FIAU PET imaging of liposome-mediated direct intratumoral HSV1-tk gene delivery and expression in glioblastoma patients (4). Yaghoubi et al. reported on the human pharmacokinetics of 18F-FHBG PET in healthy volunteers (5). A recent publication by Peñuelas et al. demonstrated that 18F-FHBG PET can monitor HSV1-tk gene expression after intratumoral injection of adenovirus in hepatocellular carcinoma patients (6).
The major limitation for translation of virus- and bacterium-derived reporter gene imaging into clinical practice is the immunogenicity of these non–human-derived reporter proteins. This limitation is especially important when repetitive administration of a reporter gene or long-term monitoring of transgene expression is required, because of the potential for compromise by an immune reaction against this foreign protein. Several human-derived reporter genes for nuclear imaging have been proposed to circumvent this limitation, including dopamine receptor type 2 (7), somatostatin receptor (8), sodium iodide symporter (9), and norepinephrine transporter (10), as well as transferrin receptor (11) and ferritin (12) for MRI. However, receptor–ligand or transporter-based approaches are hindered by a relatively low sensitivity and narrow dynamic range caused by the intrinsic saturability of receptors even at radiotracer concentrations. High levels of reporter receptor expression on the cell membrane cannot be achieved because of competition for space on the cell membrane from other receptor types, which are required for cell viability (7). Furthermore, the level of binding of imaging agents to reporter receptors is significantly influenced by levels of endogenous natural ligands, which compete for binding and determine the receptor availability or occupancy (13). An example of a transporter-based reporter gene limitation is sodium iodide symporter, which has a short duration of imageable signal because organification of the radiolabeled substrate (radioiodine) is absent (9). Ultimately, reporter gene expression should not perturb cellular homeostasis (14).
Therefore, we aimed to develop a different human-derived reporter gene encoding for a nonimmunogenic endogenous enzyme that would be suitable for molecular–genetic imaging with radionuclide imaging modalities. We used the following selection criteria: the enzyme could be expressed transiently or at low levels, be metabolically poorly accessible to a substrate without a specifically coexpressed transporter, or be expressed in only a small amount of anatomically well-defined, specific tissue.
In this paper, we report on one such suitable human-derived reporter gene, human mitochondrial thymidine kinase type 2 (hTK2). Two of the 4 human deoxyribonucleoside kinases, deoxyguanosine kinase and hTK2, are expressed in mitochondria (15). hTK2 phosphorylates deoxythymidine, deoxycytidine, and deoxyuridine, as well as several antiviral and anticancer nucleoside analogs (16). The efficient phosphorylation of FIAU and other nucleoside analogs by hTK2 has also been reported as the cause of mitochondrial toxicity (16). Previously, we reported very low (not able to be imaged) levels of FIAU accumulation in tumor cells that do not express HSV1-tk and have low proliferative activity (2). The major reason why FIAU is not efficiently entrapped by hTK2-mediated phosphorylation in naïve tumor cells is that the hTK2 is poorly accessible to FIAU. In mitochondria, the deoxyribonucleotide pool is separated from the cytosolic pool because the mitochondrial inner membrane is impermeable to charged molecules. In contrast, human thymidine kinase type 1 (hTK1) is expressed in the cytoplasm of rapidly proliferating cells and can be imaged noninvasively with PET using 11C-thymidine or other radiolabeled nucleoside analogs: 18F-fluorothymidine (17), 2′-fluoro-5-11C-methyl-1-beta-d-arabinofuranosyluracil (11C-FMAU) (18) or 18F-FMAU (19), and 124I-iodeoxyuridine (20).
In this paper, we describe a human-derived reporter gene, mitochondrial thymidine kinase type 2, that has been truncated in the N terminus to eliminate the nuclear localization signal (ΔhTK2) and, as a result, is expressed throughout the cell cytoplasm. Noninvasive imaging of retrovirus-mediated expression of ΔhTK2 in tumor xenografts in vivo was achieved using 124I-FIAU and 18F-FEAU PET.
MATERIALS AND METHODS
Human Thymidine Kinase/Green Fluorescent Protein (GFP) Retroviral Vectors
The schematic structures of retroviral vectors used in this study are presented in Figure 1. hTK2 complementary DNA (cDNA) truncated at 18Ser (ΔhTK2) and cloned into pET-14b vector with the N-terminal fusion 6xHis-tag (Gene Bank Y10498) was kindly provided by Liya Wang (Swedish University of Agricultural Sciences). The ΔhTK2 cDNA fragment lacking stop codon was amplified using 2 primers (5′-CGTACGTCACGTGCCGGCCATCGATCATATGGCATCAGTGATCTGTGTCG-3′ and 5′-ACGTACGTGGATCCTTACACGTGGGCCGGCCCTGGGCAATGCTTCCG-3′), digested with PmlI and HpaI, and subcloned into corresponding restriction sites of the retroviral vector SFG-wt-HSV1-tk/GFP described by us previously (21), replacing the HSV1-tk cDNA. The resulting vector was termed SFG-ΔhTK2 GFP (Fig. 1A). With the same primer pair, the ΔhTK2 cDNA fragment including a stop codon was amplified, digested with PmlI and BamHI, and placed into the retroviral vector SFG-HSV1-tk internal ribosomal entry site (IRES) GFP, replacing the HSV1-tk cDNA. The resulting vector was termed SFG-ΔhTK2-IRES GFP (Fig. 1A).

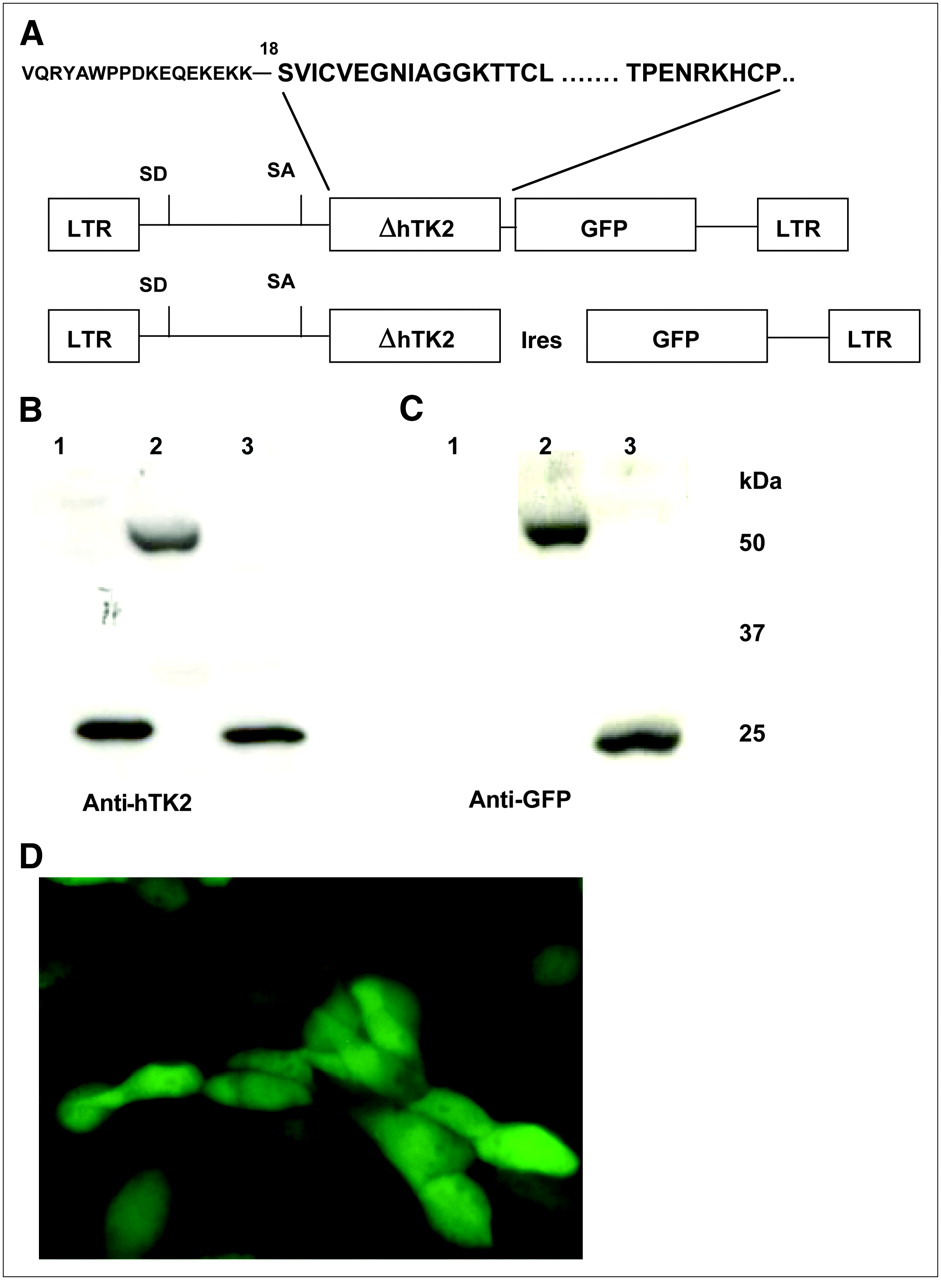
(A) Schematic representation of ΔhTK2-bearing retroviral vectors encoding ΔhTK2 GFP fusion gene and ΔhTK2 and GFP genes linked by encephalomyocarditis virus IRES element. (B and C) Western blots of ΔhTK2-expressing cell lysates with anti-hTK2 (B) and anti-GFP (C) monoclonal antibodies. (D) Fluorescent microscopy of U87/ΔhTK2 GFP cells using GFP filter set. LTR = long-terminal repeat.
Transduction of Tumor Cells
The U87 human glioma cell line was obtained from American Type Culture Collection and grown as cell monolayers in minimal essential medium supplemented with 10% fetal calf serum. The U87 cells were transduced in vitro with retroviral vectors by exposing U87 cell monolayers to a filtered (0.45-μm) culture medium obtained from the vector producer cells for 8 h in the presence of polybrene (8 μg/mL), as previously described (21).
Flow Cytometry and Fluorescent Microscopy
Retroviraly transduced U87 cells were grown as bulk cultures for 48 h and subsequently sorted for positive GFP expression using fluorescence-activated cell sorting (FACS) (FACSVantage; Becton Dickinson); the 488-nm excitation beam and 510-nm emission filter were used. Subcellular localization of the ΔhTK2 GFP in transduced tumor cells was visualized by fluorescence microscopy (Nikon) using similar excitation and emission parameters.
Production and Purification of ΔhTK2 Protein and ΔhTK2-Specific Monoclonal Antibody
The ΔhTK2 cDNA with N-terminal fusion 6xHis-tag in the pET-14b plasmid (Gene Bank Y10498) was transformed into the BL-21p(LYS)ES strain of Escherichia coli (Novogen). Inoculated medium (800 mL, with 100 μg/mL of carbenicillin) was incubated at 37°C and vigorously shaken until an optical density of 0.6 was reached. Expression was induced by adding isopropyl-β-thiogalactopyranoside (Sigma Chemical Co.) to a final concentration of 1 mM. Incubation was continued until an optical density of 1.2 was reached, and cells were harvested by centrifugation at 15,000g for 20 min at +4°C (Sorvall RC-5B). The bacterial pellet was frozen at −80°C and stored. Then, the bacterial pellet was thawed on ice and homogenized in 35 mL of lysis buffer (100 mM NaH2PO4, 0.5 M NaCl, 8 M Urea, pH 8.0) in a glass–glass homogenizer. Lysozyme (1 mg/mL, Sigma Chemical Co.) was added, and the suspension was incubated at +4°C for 30 min with shaking, followed by sonication (6 × 10 s, 80 W, on ice.). Benzoate (1 μL/mL, Novagen) and Protease Inhibitor Cocktail III (2 μL/mL, Novagen) were added, and the lysate was incubated at room temperature for 3 h and then centrifuged at 15,000g for 30 min at +4°C. The supernatant was passed through a 0.45-μm filter (Pall Corp.). ΔhTK2 protein was purified using fast protein liquid chromatography (BioLogic LP; Bio-Rad), and HiTrap affinity columns were loaded with Ni2+ (Amersham Pharmacia Biotech). The bacterial lysate was applied to a column with a flow rate of 1 mL/min and then eluted with a lysis buffer under a pH gradient of 8.0 to 4.0. Multiple fractions were collected and analyzed by sodium dodecylsulfate (SDS)–polyacrylamide gel electrophoresis (PAGE) to obtain the pure ΔhTK2 protein.
The anti-ΔhTK2 monoclonal antibody clone ΔhHTK-8G2-AF11-1 was generated at the Monoclonal Antibody Facility of Memorial Sloan-Kettering Cancer Center. Briefly, BALB/c female mice were immunized 5 times intraperitoneally with 1–6 μg of purified ΔhTK2 protein in urea emulsified in 50 μL of TiterMax (TiterMax USA, Inc.). After having been allowed to rest for more than 1 mo, the best responder was boosted with 9 μg of purified ΔhTK2 protein intraperitoneally 5 d before the splenocytes were fused with SP2/0-Ag14 (American Type Culture Collection) using polyethylene glycol-1500 (Roche). The fusion was plated 5 h later into 25- to 96-well plates and cultured for 7 d in Hybridoma-SFM (Invitrogen) supplemented with 15% fetal bovine serum, 1% hybridoma cloning factor (BioVeris), 10 mg/mL gentamycin (Invitrogen), and 1× hypoxanthine, aminopterin, thymidine (Sigma). The fusion was performed, and wells greater than 50% confluent were selected, expanded twice, and frozen over the next 2 wk. Screening involved an initial enzyme-linked immunosorbent assay using purified ΔhTK2 protein and whole-cell lysates from U87/ΔhTK2 GFP and U87/ΔhTK2-IRES GFP cells; thereafter, the enzyme-inked immunosorbent assay–positive hybridoma clones were tested by Western blot.
Western Blot of ΔhTK2, GFP, and ΔhTK2 GFP
The cells were washed with ice-cold phosphate-buffered saline and mechanically lysed in a buffer containing 10 mM TriszHCl (pH 7.5), 1 mM dithiothreitol, 20% (v/v) glycerol, 1 mM ethylenediaminetetraacetic acid, and protease inhibitor mixture (Sigma; 100 mL/100 mL buffer). The samples were centrifuged at 4°C for 5 min, and the supernatant was mixed with SDS sample buffer 1:2 and boiled for 5 min. A 10-mL sample was subjected to SDS-PAGE (10%) and transferred to Immobilon polyvinylidene difluoride membrane (Millipore). GFP and ΔhTk2 GFP were assessed using anti-GFP antibody (catalog number 8362; Clontech) at a 1:500 dilution. ΔhTK2 recombinant protein, ΔhTK2, and ΔhTK2 GFP from U87 cell lysates were assayed using the murine serum described above (dilution, 1:500) and purified monoclonal antibodies (dilution, 1:5,000). Immunochemical detection of these proteins was performed using chemiluminescence (ECL kit; GE Healthcare). The protein concentration in samples was assayed using a protein assay kit (Bio-Rad).
FIAU, FEAU, and Penciclovir Accumulation Assay
The radiotracer accumulation assays were performed as previously described (1). The cells were seeded in 150 × 25 mm tissue culture plates (Nunc) at a concentration of 1 × 106 cells per plate and grown until 50%–60% confluent. The incubation medium was replaced with 14 mL of medium containing [2-14C]FIAU, 370 Bq/mL (2,072 MBq/mmol), and [met-3H]thymidine, 7,400 Bq/mL (2,200 GBq/mmol), or [2-14C]thymidine, 370 Bq/mL (2,072 MBq/mmol), and [met-3H]FEAU or [met-3H]penciclovir, 7,400 Bq/mL (2,200 GBq/mmol) (Moravek Biochemicals). The radiochemical purity of each compound was checked in our laboratory using high-pressure liquid chromatography and found to be more than 98%. The cells were harvested by scraping after various periods of incubation (10, 30, 60, 90, and 120 min) and were centrifuged, and the cell pellets were weighed and assayed for radioactivity concentration on a TriCarb 1600 β-spectrometer (Packard) using standard 3H and 14C dual-channel counting techniques. The medium was also counted before and after incubation. The data were expressed as a harvested cell–to–medium concentration ratio (dpm/g of cells)/(dpm/mL of medium). The rates of accumulation (Ki) for FIAU, FEAU, penciclovir, and thymidine were determined from the slope of the cell-to-medium ratios versus incubation time plots and have units of tracer clearance from the medium (milliliters of medium/min/g of cells). The ratio of Ki values (the FIAU-to-thymidine, FEAU-to-thymidine, and penciclovir-to-thymidine ratios) is a measure of ΔhTK2 activity and correlates with independent measures of gene expression (1).
Assessment of Drug Sensitivity of ΔhTK2 GFP and ΔhTK2-IRES GFP Cells to Different Nucleoside Analogs
The wild-type U87 cells or U87/ΔhTK2 GFP and U87/ΔhTK2-IRES GFP cells were seeded into 96-well plates (Nunc) at 2,000 cells per well and treated with various concentrations of different nucleoside analogs for 4 d. Cell viability was assessed using a WST-1 assay (Roche) according to the manufacturer's protocol with a microplate spectrophotometer (Spectra Count; Packard BioScience Co.).
Subcutaneous Tumors and Study Groups
The experimental protocol involving animals was approved by the Institutional Animal Care and Use Committee of the Memorial Sloan-Kettering Cancer Center. The wild-type U87 cells and transduced U87 cells expressing ΔhTK2 GFP were released from the culture plates by treatment with 0.5% trypsin in phosphate-buffered saline for 5–10 min, resuspended in the growth medium to neutralize the trypsin, centrifuged to obtain the cellular pellet, and then resuspended in minimal essential medium (without fetal calf serum) at a concentration of 106 viable cells in 100 μL. Tumor cells (106 cells in 100 μL) were injected subcutaneously into 18 nu/nu mice (Taconic) weighing 20–25 g. In each mouse, 2 subcutaneous tumors—the wild-type U87 tumor and the ΔhTK2 GFP tumor—were produced by injection of 106 cells subcutaneously into the dorsal aspects of the left and right shoulders, respectively.
PET with 124I-FIAU, 18F-FEAU, and 18F-FHBG
No-carrier-added 18F-FHBG, 124I-FIAU, and 18F-FEAU were prepared as previously described (22–24). The mice were anesthetized (a 2:98 isoflurane:oxygen gas mixture). The mice were pretreated with 1 mL of 0.9% NaI solution intraperitoneally 24 h before radiotracer injection when required to block thyroid accumulation of small amounts of radiolysed 124I-iodide. In the first group of animals (n = 6), PET was performed 2 h after intravenous administration of 7.4 MBq of 18F-FEAU per animal on the first day. On the next day (24 h later), the same animal was injected intravenously with 7.4 MBq of 18F-FHBG and imaged at 2 h. On the third day, the same animal was injected intravenously with 7.4 MBq of 124I-FIAU, and PET images were acquired 24 h afterward, followed by tissue sampling. PET was performed using a dedicated small-animal PET scanner (microPET; Concorde). Images were reconstructed using an ordered-subsets expectation maximization iterative reconstruction algorithm. To estimate the tumor dosimetry of 124I-FIAU and to confirm the PET radioactivity measurements, we sampled, weighed, and assayed the individual tumors, muscle, venous blood, and liver for 124I radioactivity using a 5500 γ-spectrometer (Packard).
Two additional groups of animals (n = 6 each) were injected with 7.4 MBq of 18F-FEAU or 7.4 MBq of 18F-FHBG per animal, and PET images were acquired 2 h after tracer administration; thereafter, the animals were sacrificed, and individual tumors, muscle, and venous blood were sampled, weighed, and assayed for 18F radioactivity.
Statistics
Descriptive statistics of group data were obtained using univariate analysis. Group data were compared using ANOVA analysis, regression analysis, and the Student t test; a P value of less than 0.05 was considered significant. Statistical analysis of data was performed using StatView 4.57 (Abacus Concepts) and KaleidaGraph 3.5 (Synergy).
RESULTS
Expression and Assessment of ΔhTK2 Protein In Vitro
Western blot analysis detected highly immunoreactive bands of purified proteins at approximately 29 kDa for ΔhTK2 protein, approximately 55 kDa for ΔhTK2 GFP, and approximately 27 kDa for ΔhTK2 fusion GFP (Fig. 1B). The lower sensitivity threshold for the ΔhHTK-8G2-AF11-1 monoclonal antibody was determined to be a 1:50,000 dilution (data not shown). Western blot with anti-GFP antibodies showed a GFP-immunoreactive band for the ΔhTK2 fusion GFP at approximately 55 kDa and for the IRES-mediated coexpressed GFP at approximately 27 kDa, as predicted (Fig. 1C).
Fluorescent microscopic analysis of U87 tumor cells transduced with SFG-ΔhTK2 GFP retroviral vector demonstrated a pancellular distribution of ΔhTK2 GFP (Fig. 1D).
The levels of enzymatic activity of ΔhTK2 and ΔhTK2 GFP were assessed using an FIAU accumulation assay in 12 different populations of transduced U87 cells with 6 significantly different levels of expression of ΔhTK2 and ΔhTK2 GFP. These populations of cells with different levels of fluorescence were obtained using FACS, expanded in vitro, and reanalyzed by FACS for the levels of fluorescence of GFP and ΔhTK2 GFP.
The slopes of the relationship between GFP fluorescence and the FIAU-to-thymidine accumulation ratio (enzymatic activity of the ΔhTK2 subunit) reflects a constant relationship between the levels of expression of these reporter proteins in different transduced tumor cell populations. The magnitude of the FIAU-to-thymidine accumulation ratio (the y-intercept of the slope in Fig. 2A) in various populations of tumor cells expressing ΔhTK2 protein was about 2-fold higher than that measured in populations of cells expressing ΔhTK2 GFP when normalized by the GFP expression levels (GFP fluorescence) in corresponding populations. Overall, the levels of FIAU accumulation in cells expressing ΔhTK2 and ΔhTK2 GFP were about 10-fold higher than in wild-type cells.

(A) Comparison of tracer accumulation and GFP fluorescence in populations expressing different levels of ΔhTK2 GFP (hTG) and ΔhTK2 coexpressed with GFP (hTIG) reporter genes. (B) Radiotracer uptake assay of FIAU, FEAU, and penciclovir in wild-type and ΔhTK2 GFP transduced U87 cells in vitro. (C). Radiotracer accumulation ratio in ΔhTK2 GFP transduced/wild-type U87 cells for FIAU, FEAU, and penciclovir in vitro. PCV = penciclovir; TdR = thymidine.
The levels of enzymatic activity of ΔhTK2 GFP were assessed using FIAU, FEAU, and penciclovir accumulation assays. FIAU and FEAU were efficiently phosphorylated by ΔhTK2 GFP enzyme. In contrast, no penciclovir phosphorylation was observed for ΔhTK2 GFP enzyme (Fig. 2B). In wild-type U87 cells, background levels of FIAU, FEAU, and penciclovir accumulation were detected. The specificity of ΔhTK2 GFP enzyme phosphorylation was assessed by calculating the ratio of radiotracer accumulation in ΔhTK2 GFP transduced cells to that in nontransduced cells, as represented in Figure 2C.
To explore the possibility of killing the ΔhTK2 transduced cells using prodrug activation suicide therapy, the wild-type, ΔhTK2 GFP, and ΔhTK2-IRES GFP transduced U87 cells were tested for in vitro sensitivity to clinically relevant nucleoside analogs such as FIAU, d-arabinofuranosyl-cytosine (ara-C), FUdR, d-arabinofuranosyl-thymidine (ara-T), gemcitabine, and ganciclovir at different concentrations of these drugs. Among the tested drugs, ara-C showed an inhibitory concentration of 50% of as low as 0.04 μmol for U87-ΔhTK2 GFP cells, compared with 0.260 μmol for wild-type U87 cells (Fig. 3).

Assessment of drug sensitivity of ΔhTK2 GFP and ΔhTK2-IRES GFP transduced U87 cells to different nucleoside analogs in vitro. FUdR = fluorodeoxyuridine; GCV = ganciclovir; IC50 = inhibitory concentration of 50%.
In Vivo Imaging of ΔhTK2 GFP Gene Expression with PET
Small-animal PET imaging studies were performed on mice bearing subcutaneous tumor xenografts grown from ΔhTK2 GFP transduced U87 cell populations and wild-type U87 cells, which served as a negative control. Both transduced and wild-type tumors reached 5 mm in diameter, and no difference was observed between transduced and wild-type tumor growth. PET was performed on 3 consecutive days; therefore, there was no difference in the interval between tumor inoculation and PET with different tracers. A highly specific localization of 124I-FIAU– and 18F-FEAU–derived radioactivity was revealed in tumor xenografts expressing the ΔhTK2 GFP reporter gene (Figs. 4A and 4C). The 18F-FHBG accumulation in tumors expressing the ΔhTK2 GFP reporter gene was low and comparable with the level of radioactivity in muscle (Fig. 4B). The levels of 124I-FIAU, 18F-FEAU, and 18F-FHBG radioactivity in control (nontransduced) U87 tumors were similar to muscle activity, which was very low (Fig. 4). Tissue sampling and biodistribution studies confirmed highly specific accumulations of 124I-FIAU– and 18F-FEAU–derived radioactivity in U87-ΔhTK2 GFP tumors. The wild-type U87 tumors and other organs and tissues showed only low background levels of radioactivity (Fig. 5). Overall, the levels of 124I-FIAU and 18F-FEAU accumulation were about 6-fold higher in U87 tumors expressing ΔhTK2 GFP reporter gene than in wild-type U87 tumors.
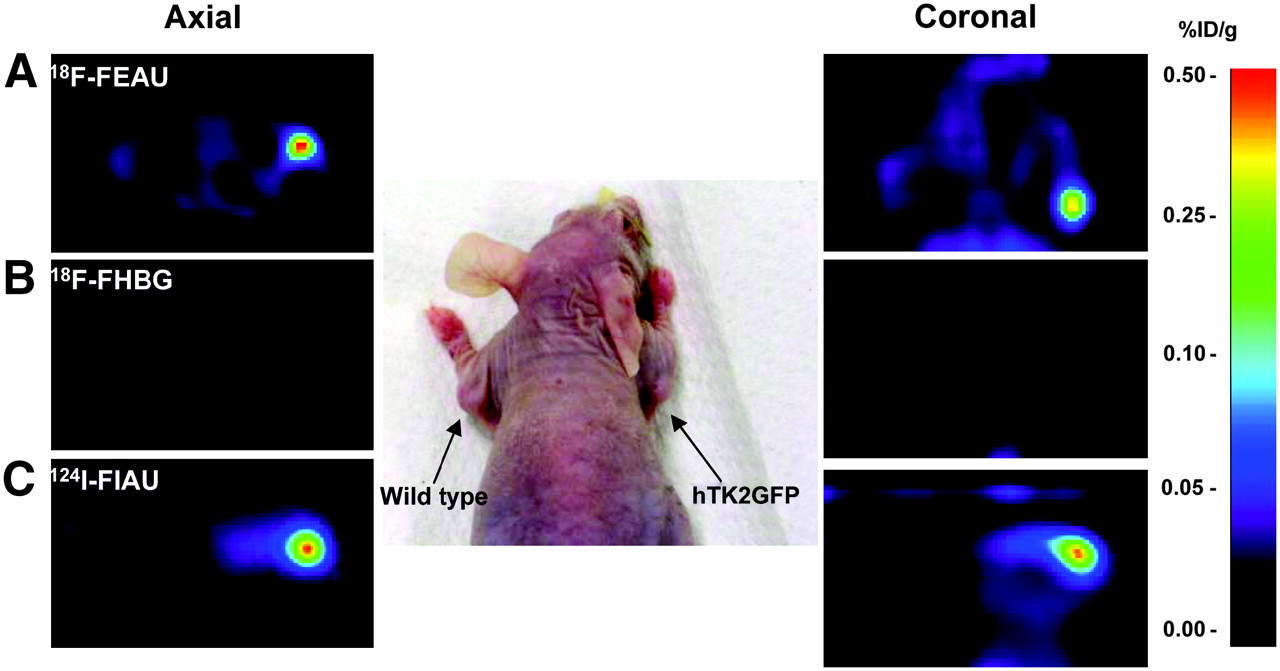
Small-animal PET imaging of ΔhTK2 GFP expression with 18F-FEAU (A) and 18F-FHBG (B) at 2 h and 124I-FIAU (C) at 24 h after radiotracer administration in same animal. U87/ΔhTK2 GFP tumor is in right shoulder; control U87 wild-type tumor is in left shoulder. %ID/g = percentage injected dose per gram of tissue.

(A) In vivo radiotracer accumulation measurements for 18F-FEAU, 18F-FHBG at 2 h, and 124I-FIAU at 24 h after radiotracer administration. (B and C) Calculated tissue-to-muscle (B) and transduced tumor–to–nontransduced tumor (C) tracer accumulation ratios in vivo. %ID/g = percentage injected dose per gram of tissue; Pl = plasma; Ms = muscle (n = 6; P < 0.05).
DISCUSSION
The effectiveness of cell- or vector-mediated gene therapy depends largely on the specificity and efficacy of gene delivery into target cells (and tissues) and the specificity, magnitude, and persistence of gene expression required for an optimum therapeutic effect. Recently, we and others demonstrated the feasibility of PET reporter gene imaging with HSV1-tk for monitoring the efficacy and durability of various cell therapies (25–29). In this paper, we describe a novel human-derived reporter gene, ΔhTK2, that is suitable for PET of genetic and adoptive cell therapies with 124I-FIAU or 18F-FEAU. We developed the ΔhTK2 reporter gene to enable studies that require long-term monitoring of therapeutic gene coexpression in normal and neoplastic tissues, including adoptive immunotherapies, stem cell therapies, and other transplantation therapies. The ΔhTK2 reporter gene was developed because the HSV1-tk reporter may induce immune-mediated rejection of gene-transfected cells and prevent long term PET-based monitoring of gene- and cell-based therapies (30). In the study reported by Riddell et al., immune responses specific for the HSV1-tk protein developed in 5 of 6 patients who had received anti-HIV cytotoxic HSV1-tk transduced T lymphocytes, and transduced T cells were eliminated (31). The rejection of genetically modified cells by these immunocompromised hosts suggests that strategies to render gene-modified cells less susceptible to host immune surveillance are required for successful gene therapy of immunocompetent hosts. Furthermore, administration of ganciclovir, which is routinely used for antiviral therapy in immunocompromised patients who have received transplanted bone marrow (stem cells), will eliminate the adoptively transferred HSV1-tk transduced stem cells or T cells. In contrast, hTK2 is intrinsically nonimmunogenic and can sensitize transduced cells to certain prodrugs (e.g., ara-C) for elimination of any undesired consequences of genetic and transplantation therapies (e.g., neoplastic transformation) while allowing patients receiving stem-cell or T-cell transplants to be treated with ganciclovir.
Several biochemical features of hTK2 make it an attractive PET reporter gene. In nonreplicating cells, where cytosolic deoxyribonucleotide synthesis is downregulated, mitochondrial DNA synthesis depends solely on the mitochondrial salvage pathway enzymes, the deoxyribonucleoside kinases, including hTK2, which is 1 of 4 human deoxyribonucleoside kinases that are expressed in mitochondria (15). The paradigm of applying ΔhTK2 as a clinically useful reporter gene is based on overexpression and retargeting of subcellular localization of ΔhTK2 protein from mitochondria into the cell cytoplasm, where it will readily be metabolically accessible to radiolabeled nucleoside analogs (124I-FIAU and 18F-FEAU). Previously, we and others have observed that proper subcellular localization of a reporter protein plays an important role in the context of apparent enzymatic activity (21). In the current study, we tested a truncated version of hTK2 (ΔhTK2) that lacks 18 N-terminal amino acids responsible for mitochondrial localization of this protein (32). When overexpressed in fusion with GFP, ΔhTK2 GFP exhibited a pancellular distribution in transduced cells, resulting in a significant increase in accumulation of the pyrimidine nucleoside analogs FIAU and FEAU in tumor cells expressing high levels of ΔhTK2 or ΔhTK2 GFP (Fig. 1). These data support our previous conclusions that cytoplasmic retargeting improves the bioavailability of reporter enzymes for corresponding reporter substrates (21). In rapidly proliferating cells, hTK1, being naturally expressed in the cell cytoplasm, phosphorylates thymidine and can phosphorylate FIAU (16), albeit to a significantly lesser degree. However, as we have demonstrated in this work (Figs. 2B and 2C), hTK1 cannot phosphorylate FEAU because of the steric clash between the larger-than-iodine 5-ethyl group of FEAU and amino acid Thr163 in the active site of hTK1 enzyme (33). We determined in vitro that FEAU is a more specific reporter substrate for ΔhTK2 than for hTK1, as compared with FIAU (transduced-to-nontransduced ratios in vitro, 75 for FEAU and 9.7 for FIAU).
Although FIAU phosphorylation was approximately 4 times lower by ΔhTK2 than by the HSV1-tk enzyme expressed at similar levels (FIAU-to-thymidine ratios, 0.068 and 0.3, respectively (21)), the accumulation of 18F-FEAU and 124I-FIAU in ΔhTK2 transduced tumors in vivo was more than sufficient for successful imaging with PET (transduced-to-nontransduced ratio, approximately 6). Others have reported similar or even lower values for different human-derived reporter genes suitable for nuclear imaging (7–9,34). Although FIAU is usually labeled with 124I for PET studies (or with 123I or 131I for γ-camera scintigraphy) and “late” imaging is usually performed, the feasibility of “early” FIAU imaging has been demonstrated by our group. Good-quality images can be obtained within the first 30 min to 2 h after radiolabeled FIAU administration (2). Our current data demonstrate that 18F-FEAU can be used for imaging of ΔhTK2 reporter gene expression at 2 h after tracer administration. Interestingly, although FEAU was shown to be superior to FIAU in vitro (transduced-to-nontransduced ratios, 75 and 9.7, respectively; Fig. 2C), “late” imaging of 124FIAU due to the longer 124I half-life (∼4 d) makes these tracers equally effective for PET in vivo (transduced-to-nontransduced ratios, 6.67 for FIAU at 24 h and 5.58 for FEAU at 2 h; Fig. 5B). Radiolabeling of FIAU and FEAU with 11C is an alternative strategy and would provide additional positron-emitting probes for PET of ΔhTK2 with high sensitivity and dynamic range. In addition, the very short half-life of 11C (20 min) should facilitate more frequent repetitive PET and would allow for sequential PET of ΔhTK2 expression with 11C-FEAU, followed 4 h later by PET with a different tracer in the same subject.
CONCLUSION
We have developed, characterized, and validated in an animal tumor model in vivo a novel human-derived reporter gene, ΔhTK2, that is likely to be nonimmunogenic for molecular genetic imaging with PET using specific radiolabeled nucleoside analogs (FIAU and FEAU) as reporter probes. ΔhTK2 can be used as a reporter gene and as the pro-drug activation gene for therapy with routinely used anticancer nucleoside analogs, such as ara-C. We expect that ΔhTK2 can rapidly be translated into clinical applications that require imaging for effective monitoring of various genetic and cellular therapies.
Acknowledgments
This work was supported by several grants: NIH P50 CA86438-01, R01 CA102352, DOE DE-FG02-02ER63481, and DOE DE DE-FG02-86ER-60407. We thank Dr. Ronald Blasberg for helpful discussions and advice on preparation of the manuscript, Drs. William Bornmann and Ronald Finn for help with radiotracer production and advice on preparation of the manuscript, and Vilia Tourkova for excellent assistance with the in vitro studies.
Footnotes
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication October 5, 2006.
- Accepted for publication January 20, 2007.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Imaging of T-cell Responses in the Context of Cancer Immunotherapy
- Targeted molecular-genetic imaging and ligand-directed therapy in aggressive variant prostate cancer
- Comparative Analysis of T Cell Imaging with Human Nuclear Reporter Genes
- Evaluation of Prostate-Specific Membrane Antigen as an Imaging Reporter
- Noninvasive Molecular Imaging Using Reporter Genes
- Long-term in vivo monitoring of mouse and human hematopoietic stem cell engraftment with a human positron emission tomography reporter gene
- Early Stem Cell Engraftment Predicts Late Cardiac Functional Recovery: Preclinical Insights From Molecular Imaging
- Structure-guided Engineering of Human Thymidine Kinase 2 as a Positron Emission Tomography Reporter Gene for Enhanced Phosphorylation of Non-natural Thymidine Analog Reporter Probe
- Cardiovascular Molecular Imaging: Focus on Clinical Translation
- A New Pyrimidine-Specific Reporter Gene: A Mutated Human Deoxycytidine Kinase Suitable for PET During Treatment with Acycloguanosine-Based Cytotoxic Drugs
- Tracking Cell Fate With Noninvasive Imaging
- Molecular-Genetic PET Imaging Using an HSV1-tk Mutant Reporter Gene with Enhanced Specificity to Acycloguanosine Nucleoside Analogs
- Molecular Imaging: Antidote to Cardiac Stem Cell Controversy
- Monitoring the Efficacy of Adoptively Transferred Prostate Cancer-Targeted Human T Lymphocytes with PET and Bioluminescence Imaging
- Noninvasive Imaging of Cell-Mediated Therapy for Treatment of Cancer
- Molecular-Genetic Imaging Based on Reporter Gene Expression
- PET Imaging of Cancer Immunotherapy
- A New Acycloguanosine-Specific Supermutant of Herpes Simplex Virus Type 1 Thymidine Kinase Suitable for PET Imaging and Suicide Gene Therapy for Potential Use in Patients Treated with Pyrimidine-Based Cytotoxic Drugs