


Abstract
Transplantation of progenitor cells (PCs) has been shown to improve neovascularization and left ventricular function after myocardial ischemia. The fate of transplanted PCs has been monitored by fluorescence labeling or by genetic modifications introducing reporter genes. However, these techniques are limited by the need to kill the experimental animal. The aim of this study was to radiolabel CD34+ hematopoietic PCs (HPCs) with 111In-oxine and to evaluate the feasibility of this in vivo method for monitoring myocardial homing of transplanted cells in a rat myocardial infarction model. Methods: Human HPCs were isolated from mobilized peripheral blood and labeled with 111In-oxine. Labeled HPCs were injected into the cavity of the left ventricle in nude rats 24 h after induction of myocardial infarction (n = 4) or sham operation (n = 4). Scintigraphic images were acquired up to 96 h after HPC injection. After animals were killed, tissue samples of various organs were harvested to calculate tissue-specific activity and for immunostaining. Results: Labeling efficiency of HPCs was 32% ± 11%. According to trypan-blue staining, viability of radiolabeled HPCs was impaired by 30% after 48 and 96 h in comparison with unlabeled cells, whereas proliferation and differentiation of HPCs was nullified after 7 d, as assessed by colony-forming assays. After injection of HPCs, the specific activity ratio of heart to peripheral muscle tissue increased from 1.10 ± 0.32 in sham-operated rats to 2.47 ± 0.92 (P = 0.020) in infarcted rats. However, the overall radioactivity detected in the heart was only about 1%. A transient high lung uptake of 17% ± 6% was observed within the first hour after infusion of HPCs. At 24 h after injection, the initial lung activity had shifted toward liver, kidneys, and spleen, resulting in an increase of radioactivity in these organs from 37% ± 6% to 57% ± 5%. Conclusion: Radiolabeling with 111In-oxine is a feasible in vivo method for monitoring transplanted HPCs in a rat myocardial infarction model. The potential to detect differences in myocardial homing between infarcted and normal hearts suggests that this method may provide a noninvasive imaging approach for clinical trials using transplanted HPCs in patients. Our findings, however, also demonstrated a negative effect of 111In-oxine on cellular function, which resulted in complete impairment of HPC proliferation and differentiation. For future trials in stem cell imaging with 111In-oxine, therefore, it will be mandatory to carefully check for radiation-induced cell damage.
- CD34+
- hematopoietic progenitor cells
- 111In-oxine
- rat myocardial infarction model
- cell trafficking
- radiation-induced cell damage
Novel approaches to cell therapy that may contribute to neovascularization and remodeling of acute ischemic tissue in myocardial infarction or chronic ischemic heart disease have been reported in both preclinical and clinical studies. Infusion of both ex vivo cultivated endothelial progenitor cells (EPCs) or CD34+ hematopoietic progenitor cells (HPCs) improved neovascularization and left ventricular function after myocardial ischemia (1–4). However, little quantitative data on the biodistribution and in vivo kinetics of progenitor cells (PCs) have been reported. Tissue distribution of these cells has been monitored by fluorescence labeling of transplanted PCs or by genetic modifications introducing genes for fluorochromes or metabolic enzymes (5–7). Detection of fluorescence or enzyme-produced colorimetric reactions in different tissues and organs, however, is limited by the need to kill the animal, precluding the use of this technique in humans. In addition, immunohistochemical analysis of tissue sections does not allow quantification of the homing of the transplanted autologous cells to different organs and tissues to describe distribution within the whole body. Moreover, detection by immunohistochemical methods is limited in assessing the time course of cell trafficking, because it is not possible to investigate the same animal repeatedly. Thus, alternative noninvasive monitoring methods that are also applicable in patients are necessary to get additional insights in PC trafficking and homing.
Radiolabeling of cells has been widely used to monitor the fate and tissue distribution of blood cells. Imaging of leukocyte distribution, for example, is a routine clinical procedure to localize areas of inflammation (8–10). For this purpose, commercially available and commonly used 111In-oxine has been proven a safe and easy-to-use radiolabel. 111In compounds have been applied successfully in various experimental settings to determine the biodistribution of transplanted hepatocytes (11), the migration patterns of dendritic cells (12,13), the homing of mesenchymal stem cells (14), and the physiologic recirculation of lymphocytes (15). In a first report on 111In-oxine–labeled EPCs, we showed the feasibility of this method for monitoring EPCs in rats after myocardial infarction (16).
The aims of this animal study, based on our previous findings for EPCs, were to radiolabel HPCs with 111In-oxine, monitor the biodistribution of transplanted HPCs in a rat myocardial infarction model, and investigate the impact of the labeling procedure on cellular function (1,3).
MATERIALS AND METHODS
Cell Preparation
CD34+ HPCs were obtained from cryopreserved samples after allogeneic transplantation of CD34-selected grafts in accordance with informed consent. Stem cells were mobilized from healthy donors with granulocyte colony-stimulating factor (CSF; 5 μg/kg/d), and peripheral blood stem cells (PBSC) were collected using a Cobe Spectra (Cobe Laboratories). Next, CD34+ cells were immunomagnetically selected from PBSC using the automated CliniMacs device (Miltenyi-Biotech) after labeling with CD34 antibodies (CD34 MicroBeads; Miltenyi-Biotech) for 30 min at room temperature as described previously (17). Enrichment of CD34+ cells resulted in a purity of >95%, as assessed by flowcytometric analysis using CD45-fluoroisothiocyanate/CD34-phycoerythrin/7-aminoactinomycin staining. After transplantation of freshly selected grafts, small pilots were cryopreserved in liquid nitrogen using X-Vivo 10 medium (Bio Whittaker) supplemented with 10% dimethyl sulfoxide for subsequent analyses and safety control. Several years after transplantation, the anonymized pilots with the CD34+ selected cells were thawed using phosphate-buffered saline (PBS) + 1 mmol/L ethylenediaminetetraacetic acid. After spinning, cells were resuspended in RPMI + 10% fetal calf serum (FCS) + 20 ng/mL stem cell factor (SCF; R&D) + 20 ng/mL interleukin-3 (IL-3; R&D) and kept at 5% CO2 and 37°C for 1 h until experimental use.
Radiolabeling
HPCs were thawed and preactivated by stimulation with 20 ng/mL SCF and IL-3 in RPMI + 10% FCS. After 2 washes with PBS, HPCs were incubated with 30 MBq 111In-oxine (37 MBq/mL; Nycomed Amersham) for 60 min at 37°C in serum-free medium. To remove excess unbound radioactivity, all cells were washed twice with serum-free medium. Labeling efficiency was measured with a dose calibrator (Atomlab 100; Biodex Medical). Before injection, cells were resuspended in 0.5 mL of serum-free medium in 1.0-mL syringes with 27-gauge needles.
Functional Assays
For evaluation of cellular effects of 111In-oxine labeling, 1 × 106/mL HPCs were labeled with 30 MBq 111In-oxine for 60 min and were then seeded into 15-mL polypropylene tubes. After 1, 24, 48, and 96 h (n = 3 each), medium was removed and radiolabeled HPCs were stained by trypan-blue for assessing cell viability. Radiolabeled HPCs were compared with untreated cells to evaluate cell death induced by the radiolabel.
To further assess the influence of radiolabeling on HPC function, migration assays were performed using transwell plates (Costar) in a modified Boyden chamber with filter membranes (5 μm pores) coated with fibronectin (20 μg/mL) in PBS for 30 min at 4°C. Before adding CD34+ cells to the upper compartment of the transwells, filter membranes were washed twice with Iscove medium supplemented with 10% FCS (complete medium; Biochrome). Then, 1 × 105 CD34+ cells were added to the upper compartment in 0.1 mL of medium, and cells were allowed to migrate through the membrane toward the lower chamber in response to 0.6 mL of medium with 100 ng/mL stromal cell–derived factor (SDF-1; R&D). The transwell plates were incubated at 37°C and 5% CO2 for 2 h. Then, the filter membranes were carefully discarded, and the cells that had migrated into the lower compartment were fixed by adding 0.6 mL of 4% formaldehyde. Cells were allowed to attach to the lower plastic surface of the plates and were scored by microscopic analysis of 5 random fields.
In addition, we performed experiments on the capacity of HPCs to produce colonies on semisolid media. HPCs can proliferate into clones of differentiated progeny. CD34+ cells (1 × 104) were inoculated into 1.5 mL of semisolid methylcellulose (MethoCult; Stem-Cell Technologies) containing FCS, human SCF, human granulocyte-macrophage CSF, human IL-3, and human erythropoietin and were allowed to proliferate and differentiate into colony-forming units (CFUs). CFUs were scored after 7 d of incubation.
Animals and Study Design
Human cells were transplanted into a xenogeneic rat model using immunodeficient athymic rnu:rnu rats (5- to 7-wk-old females, 120 ± 30 g body weight; Charles River). Animals were anesthetized with intramuscular ketamine (100 mg/kg; Curamed) and midazolam (2 mg/kg; Hoffmann-LaRoche). To prevent arrhythmias after cardiac surgery, intramuscular amiodarone (5 mg/kg; Sanofi-Synthelabo) was given prophylactically. Furthermore, to reduce intra- and postoperative pain, piritramide (2 mg/kg; Janssen-Cilag) was administered. A 17-gauge endotracheal tube was inserted for volume-controlled ventilation of the animals. Open-chest cardiac surgery was performed after left thoracotomy to occlude the left anterior descending coronary artery (LAD) by passing a 5-0 suture around the LAD just under the tip of the left auricle (n = 4). Sham-treated animals underwent left thoracotomy and incision of the pericardium only (n = 4). After the chest wound was closed, rats were allowed to recover for 24 h before injection of HPCs.
111In-Oxine–labeled HPCs with a mean activity of 9.3 ± 3.4 MBq were administered using an x-ray–assisted transdiaphragmatic approach for intracavitary administration into the left ventricle of either sham-treated (n = 4) or infarcted (n = 4) rats. Because intracoronary application cannot be performed in rats, intracavitary administration of HPCs was considered the closest approach possible to the intracoronary cell administration in patient studies. During this procedure, contrast medium (Isovist; Schering) was injected to confirm proper left ventricular position of the needle using x-ray control.
Three additional animals received pure 111In-oxine as controls to assess the distribution of free 111In-oxine. In another 3 animals with no surgical pretreatment, radiolabeled HPCs were administered to exclude the surgical procedure as a reason for cardiac homing.
For scintigraphic imaging, animals were anesthetized with ketamine and midazolam. Planar whole-body pinhole images and spot images of the thorax were acquired over 30 min at 1, 24, 48, and 96 h after cell administration using a double-head γ-camera (ECAM; Siemens) equipped with a pinhole collimator with an insert for medium energy. The energy windows were centered at 171 keV ± 10% and 245 keV ± 10%, and images were stored in a 512 × 512 matrix.
After the last image acquisition, animals were killed. All animal experiments were performed with approved consent by the local Animal Research Committee in accordance with both federal animal protection and radiation protection laws (V 252-72241.122-17).
Tissue Preparation and Immunostaining
To detect CD34+ cells in histologic sections, we used the green cell tracker 5-chloromethylfluoresceindiacetate (CMFDA; Molecular Probes) for colabeling. After the rats were killed, transverse slices of the base, midregion, and apex of the hearts as well as tissue samples from other organs, such as skeletal muscles, lungs, kidneys, liver, bones, and spleen, were obtained. All specimens were weighed, and the counting rate was measured in a lead-shielded and calibrated well counter (LB 5310; Berthold) to calculate the specific activity per gram of tissue sample per megabecquerel of injected activity after correction for radioactive decay. Organ samples were mounted in TissueTek OCT compound freezing medium (Sakura) and snap frozen in 2-methylbutane prechilled by liquid nitrogen. Sections of 5 μm were cut and examined for human cells. Destruction of cardiomyocytes as a result of myocardial infarction was visualized by staining of myocytes with α-sarcomeric actinin (Sigma), followed by goat-antimouse Cy3 (red; Dianova). Nuclei were stained with TO-PRO-3 (blue; Molecular Probes), and sections were analyzed using a confocal microscope (LSM 510; Zeiss).
Statistical Analysis
Data presented in the results represent mean ± SD. The Student t test for unpaired data was used to evaluate statistical differences between rat subgroups, with P < 0.05 considered to be statistically significant.
RESULTS
HPC Labeling with 111In-Oxine
To test the efficiency of the labeling procedure, HPCs were incubated with 30 MBq 111In-oxine in serum-free medium for 60 min. Incorporation of 111In-oxine in percentage of added activity yielded a mean labeling efficiency of 31.9% ± 11.0% after 60 min.
Next, we investigated whether the labeling affects cell viability. We counted the number of dead cells by trypan-blue staining in both radiolabeled and unlabeled cells at 1, 24, 48, and 96 h (Fig. 1). Up to 24 h after radiolabeling, no significant differences were observed, whereas at 48 (P = 0.030) and 96 h (P = 0.011), an increased number of dead cells was found after radiolabeling of HPCs (n = 3 each).

Trypan-blue staining. Values are given as percentages of dead cells of controls as determined by counting 5 microscopic random fields of view (n = 3 each). Significant differences were found at 48 and 96 h after radiolabeling.
To check specific cellular functions, we performed migration studies with HPCs in a modified Boyden chamber in response to the chemotactic SDF-1. A significant reduction of SDF-1–induced migration was already detected 24 h after radiolabeling, resulting in a decrease in the number of migrated radiolabeled HPCs to 74.2% of the number of unlabeled control cells (P < 0.001; n = 3 each). At 48 h, SDF-1–induced migration was nullified in radiolabeled cells and reduced to 59.8% of the initial value in unlabeled cells.
In addition, we performed experiments on the capacity of 111In-oxine–labeled HPCs to produce colonies on semisolid media. HPCs were labeled for 1 h and then allowed to proliferate and differentiate into CFUs, which were scored after 7 d. Although untreated cells showed 164 ± 55 CFUs per 10,000 inoculated HPCs, no CFUs were detected in radiolabeled cells (n = 3 each).
Finally, we checked the activity of 111In in the supernatants as well as in the cells to determine the loss of 111In from the cells into the supernatant. We found that 33.5% ± 10.3%, 63.9% ± 0.9%, and 74.5% ± 10.2% of 111In initially incorporated into HPCs was released into the supernatant after 24, 48, and 96 h, respectively.
Distribution of Radiolabeled HPCs
After 111In-oxine–labeled human HPCs were administered directly into the left ventricular cavity of infarcted or sham-operated rats, a high tracer accumulation of 37% ± 6% of the injected activity was found in the liver, kidneys, and spleen at 1 h after injection (Fig. 2). A transient high lung uptake of 17% ± 6% was observed within the first hour after infusion of HPCs (Fig. 2). At 24 h after injection, the initial lung activity was no longer detectable. It had been shifted to liver, kidneys, and spleen, resulting in an increase of radioactivity in these organs to 57% ± 5% during the first 24 h. Tracer distribution then remained stable up to 96 h after injection. At all time points in rats with myocardial infarction, a diffuse heart uptake could be detected, whereas no activity accumulation was found within the heart in sham-operated controls (Fig. 2). The pinhole collimator provided a sufficiently high resolution to discriminate the heart from liver and spleen, depicting ischemia-induced heart uptake in infarcted rats.
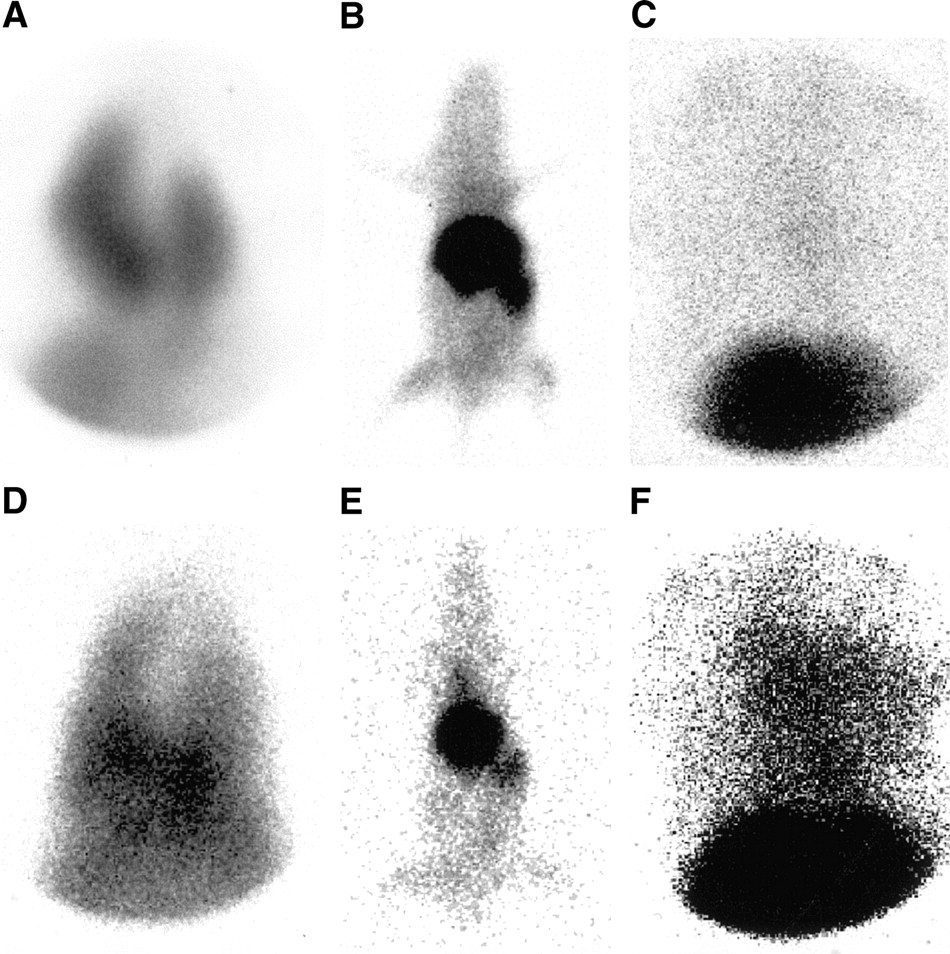
Pinhole images of nude rats without (A–C) or with (D–F) myocardial infarction after injection of 111In-oxine–labeled HPCs into the left ventricular cavity. At 1 h after injection, a high lung uptake was visible on the spot images of the chest and abdomen (A and D), but was no longer detectable on images at 24 (B and E) and 96 (C and F) h after injection. The spot images of the chest and abdomen after 96 h clearly showed significant differences in cardiac uptake between infarcted (F) and sham-operated (C) animals.
After 96 h, animals were killed, and the specific radioactivity per gram of tissue per megabecquerel of injected activity was measured in different tissues. Consistent with the in vivo images, liver, spleen, and kidneys revealed the highest specific tissue activities (Fig. 3). Because the specific activity of skeletal muscle was a constant parameter, we calculated the heart-to-muscle specific activity ratio (i.e., the ratio of specific radioactivity of the heart compared with peripheral skeletal muscle tissue) as a reference parameter for comparison. In sham-operated animals, the heart-to-muscle specific activity ratio was 1.10 ± 0.32. It significantly increased to 2.47 ± 0.92 (P = 0.020) when HPCs were infused in rats with previous infarction. After correction for radioactive decay, the amount of radioactivity in the whole heart was 1% of the injected activity in infarcted rats, which correlated to 1 × 104 HPCs.

Distribution pattern of specific activity (megabecquerel per gram of tissue per megabecquerel of injected activity) in different tissues after left intraventricular administration of 111In-oxine–labeled HPCs in infarcted or sham-operated nude rats. Data are given as mean ± SD (n = 4 each). Significant differences between organs of sham-operated and infarcted animals were observed only in the heart.
In 3 animals treated with 111In-oxine alone, a high blood-pool activity was observed up to 96 h after injection as a result of the strong affinity of oxine to serum transferrin. According to the scintigrams, blood samples at 96 h showed a more than 25-fold higher specific blood activity compared with animals treated with 111In-oxine–labeled HPCs. In 3 rats with no surgical pretreatment, no significant differences were found for specific heart activity when compared with sham-operated animals. These findings exclude the thoracotomy itself as a reason for myocardial homing of HPCs.
Fluorescence Microscopy and Immunostaining
To confirm results obtained by measuring the tissue specific radioactivity, tissue sections were analyzed by immunostaining and fluorescence microscopy (Fig. 4). The human HPCs could be identified within the rat tissue by labeling with the green cell tracker CMFDA. Although HPCs were rarely found in a scattered distribution pattern in the heart of sham-operated animals, the hearts of infarcted rats revealed human cells predominantly located in the border zone of the infarction, as evidenced by visualizing neighboring cardiac myocytes with α-sarcomeric actinin. In addition, tissue sections of other organs in which most of the radioactivity was detected were analyzed. HPCs were predominantly found in spleen and liver, whereas few HPCs were seen in the kidneys. In sections of thigh muscles, the same scattered distribution pattern of HPCs was found as in myocardial sections of sham-operated rats.

CMFDA cell-tracker–labeled human CD34+ HPCs (green) were identified in the infarct border zone delineated by damaged α-sarcomeric actinin-positive cardiac myocytes (red). Nuclei were stained with TO-PRO-3 (blue).
DISCUSSION
111In-Oxine is a well-known, safe, and commercially available tracer for radiolabeling and monitoring of blood cells. Inflammation scintigraphy with 111In-oxine–labeled leukocytes, for example, is a standard nuclear medicine procedure (8). In an earlier study on 111In-oxine–labeled EPCs, we demonstrated the feasibility of this in vivo method for monitoring EPCs in rats, finding an increased homing of these cells in the heart after myocardial infarction (16). The results of the present study show that radiolabeling of transplanted human CD34+ HPCs (a precursor of EPCs) with 111In-oxine is also feasible for monitoring myocardial homing and biodistribution of these cells in rats over a somewhat limited period of 24–48 h. General limitations in dealing with 111In-oxine in experimental settings must be addressed. Inherent to this radiopharmaceutical are the high energy of 111In (171 and 245 keV), requiring medium-energy collimators with a restricted spatial resolution; limited cellular retention of 111In; and irradiation of radiolabeled cells, which can result in impaired cellular viability and function.
The physical 2.8-d half-life of 111In allows us to monitor cell distribution for about 1 wk, which we considered a clear advantage over 99mTc-labeled radiopharmaceuticals such as 99mTc-hexamethylpropyleneamine (with a 6-h half-life). The limitation of 111In, however, is the need for medium-energy collimators. Although the spatial resolution of these collimators is rather poor, it is sufficient for clinical application in patients. However, in nude rats with a mean weight of 120 g and a heart size (about 10 mm) just in the range of or below collimator resolution, the relatively low uptake of HPCs in the heart (only 1% of injected acitvity) could not be distinguished from high tracer accumulation in the adjacent liver and spleen. An alternative to increasing resolution is the use of a pinhole collimator. We showed that pinhole collimators are appropriate tools to obtain a sufficiently high resolution to reliably discriminate differences between heart activities in rats with and without myocardial infarction. For an optimal resolution, however, only 1 animal can be scanned at a time, and long acquisition times of 30 min are required. For studying small animals, such as mice or rats, PET with a high spatial resolution would be the ideal imaging modality. However, when we tested the widely available 18F-FDG as a radiolabel, an insufficient labeling efficiency of less than 10% was obtained for both EPCs and HPCs, with a high efflux rate within the first hour. In addition, the short physical half-life of 110 min for 18F-FDG does not seem adequate for monitoring cell trafficking. 64Cu-Pyruvaldehyde bis(N4-methylthiosemicarbazone) has been shown to be an alternative PET tracer for cell labeling with a potentially suitable half-life of 12.7 h (18), but this tracer is not routinely available and therefore limited to a few PET centers. Thus, to extend PC imaging from the experimental setting to clinical application, we decided to use 111In-labeled PCs for our studies despite the limited resolution in rats and to combine scintigraphic imaging with in vitro measurement of tissue-specific activities in the heart and other organs.
Up to 96 h after injection of radiolabeled HPCs, radioactivity was predominantly located in spleen, liver, and kidneys. Immunostaining revealed many intact HPCs in the spleen and liver. These organs have been identified as the major site for homing of immunocompetent cells (19–21). In contrast, although specific tissue activity was high in the kidneys, HPCs were rarely detectable in these organs. Loss of 111In from labeled cells is a well-known phenomenon and has been described as a reason for high uptake of radioactivity in liver and kidneys (22). Because the binding of this compound to intracellular structures is reversible, 111In is released from the labeled cells over time. For lymphocytes, a 70% loss is reported within 24 h after cell labeling (22). Lower release rates were observed in our study, with a loss of 34% of the initially cell-bound activity within 24 h into the supernatant of cultured HPCs. We consider this well-known efflux of 111In as the main reason for the high radioactivity uptake in the kidneys. According to this assumption, radioactivity in the kidneys was not correlated to the number of HPCs as demonstrated by immunostaining. This nonspecific tracer signal caused by the in vivo efflux of 111In, however, does not seem to be a major concern outside liver and kidneys, because the number of HPCs were correlated with the tissue-specific activity in the heart, muscles, lungs, and spleen. The significant difference in specific heart radioactivity between sham-operated and infarcted rats did represent the different myocardial accumulation of engrafted HPCs as shown by immunostaining.
Administration of HPCs after myocardial infarction increased the specific activity in the heart significantly. It was 2.3-fold higher in rats with infarction than in sham-operated rats. This is in accordance with previous studies, which demonstrated that tissue ischemia is a major stimulus for incorporation of circulating PCs (6,23,24). Even after infarction, however, the activity detected in the heart was quite low. Our data revealed that the absolute level of HPCs homing to the heart after myocardial infarction was about 1% of the injected activity, which corresponded to 1 × 104 HPCs. Thus, a significantly lower myocardial uptake of HPCs was observed than the 3% for EPCs (16). Measuring the tissue-specific activity, however, results in a relative underestimation of myocardial cell incorporation at 96 h because of the leakage of 111In from labeled cells. Thus, the number of incorporated cells as determined by tissue-specific activity represents the lower limit of true cell accumulation in the heart. Based on the in vitro leakage rate of 75% within 96 h, the true number of accumulated cells could be in the range of 4%.
HPCs showed a transient high lung uptake of 17% ± 6% immediately after intracardial infusion, a phenomenon not observed after administration of EPCs (16). As a result of this considerable lung uptake, HPC uptake in liver, spleen, and kidneys at 1 h after injection was about 50% lower than the uptake of EPCs (37% and 72%, respectively). Up to 24 h after injection, activity shifted from the lungs toward liver, spleen, and kidneys, reaching activity levels in these organs comparable to those of EPCs.
The main obstacles in dealing with 111In-oxine, however, are potential radiation-induced alterations of specific cellular features that have been reported for various cell types. These alterations may be caused by either irradiation or toxic effects of oxine. In activated T-lymphocytes, for example, a significant loss of cytotoxic activity and proliferation has been described with no alteration of cell viability or phenotype (25). In 111In-oxine–labeled activated neutrophils, upregulated adhesion and phagocytosis rates in addition to a decreased chemotaxis were found in comparison with unlabeled cells (26). Therefore, in our previous study it was an important finding that in EPCs 111In-oxine did not significantly affect cell viability, proliferation, migration capacity, or receptor-mediated 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine–labeled acetylated low-density lipoprotein (DiLDL) uptake (16). These findings suggested that 111In-oxine–labeled human EPCs maintain their characteristic features. Different results, however, were obtained in HPCs, which are less differentiated than EPCs. Although the in vitro viability of radiolabeled HPCs, as measured by trypan-blue staining, was impaired by only about 30% in comparison with unlabeled cells after 48 and 96 h, proliferation and differentiation, as assessed by colony-forming assays, were nullified after 7 d of incubation. The migratory capacity in radiolabeled HPCs was reduced after 24 h and completely suppressed after 48 h. Thus, EPCs seem to be much more resistant to radiation-induced damage than are immature multipotent HPCs. Also contributing to the increased radiosensitivity of HPCs may be the fact that in this study we used HPCs that were frozen until clinical use. Nevertheless, HPCs were incorporated into the infarcted myocardium as shown by measurement of the heart-specific activity and by immunostaining. Moreover, HPC trafficking with a significant shift of activity from the lungs to liver and spleen within the first 24 h after injection could be demonstrated. Thus, 111In-oxine–labeled human HPCs can be used to monitor cell trafficking at least up to 24 h after injection. For cell therapy studies, however, radiolabeled HPCs do not seem suitable because of complete impairment of cell proliferation and differentiation. Only a small portion of HPCs may be radiolabeled with 111In-oxine if this imaging approach is planned for clinical studies on HPC therapy to document the myocardial homing of transplanted cells. In summary, our results clearly demonstrated the impact of 111In-oxine on cell proliferation and cellular function, which can vary substantially between different PC types. In future patient trials on stem cell imaging it will be mandatory to carefully screen for radiation-induced damage and impairment of cellular function.
CONCLUSION
The results of this animal study demonstrated that 111In-oxine is a usable tracer for in vivo monitoring of transplanted CD34+ HPCs in a rat model of myocardial infarction. The potential of this method to detect differences in myocardial homing between infarcted and normal rat hearts, as proven by immunohistochemical staining, suggests that radiolabeling of transplanted HPCs with 111In-oxine may provide a noninvasive imaging approach in patients. This study, however, also demonstrated the negative impact of 111In-oxine, resulting in complete impairment of HPC proliferation and differentiation, which is in contrast to our previous experience with 111In-oxine in more differentiated EPCs. Thus, the results of this study made clear that for future trials on stem cell imaging it will be mandatory to carefully check for radiation-induced cell damage.
Acknowledgments
We thank Christiane Mildner-Rihm and Marion Muhly-Reinholz (Molecular Cardiology, University of Frankfurt, Germany), and Kerstin Brötzmann (Department of General and Thoracic Surgery, University of Kiel, Germany), for excellent technical assistance. We also thank Dr. Hans-Ulrich Wottge and his team from the Animal Center at the University Hospital Kiel for providing excellent animal care service. We thank Rosemarie Grams (Department of Obstetrics and Gynecology, University of Kiel) for her support. We are grateful for the use of the animal laboratory facilities of the Department of General and Thoracic Surgery and the laboratory facilities of the Department of Obstetrics and Gynecology, both at the University Hospital Kiel, and the radioiosotope laboratory of the Department of Nuclear Medicine at the University of Frankfurt. This work was supported by a research grant from the University of Kiel, by a young investigator grant from the University of Frankfurt, and by a research grant (Di 600/4-1) from the Deutsche Forschungsgemeinschaft.
Footnotes
Received Jun. 18, 2003; revision accepted Nov. 24, 2003.
For correspondence or reprints contact: Winfried Brenner, MD, University of Washington Medical Center, Division of Nuclear Medicine, 1959 NE Pacific St., Box 356113, Seattle, WA 98195-6113.
E-mail: winbren_2000{at}yahoo.com
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Tadalafil, a long acting phosphodiesterase inhibitor, promotes bone marrow stem cell survival and their homing into ischemic myocardium for cardiac repair
- Bone Marrow Cell Trafficking Analyzed by 89Zr-oxine Positron Emission Tomography in a Murine Transplantation Model
- Comparison of 18F-Labeled Fluoroalkylphosphonium Cations with 13N-NH3 for PET Myocardial Perfusion Imaging
- 18F-FDG Labeling of Mesenchymal Stem Cells and Multipotent Adult Progenitor Cells for PET Imaging: Effects on Ultrastructure and Differentiation Capacity
- CXC-Chemokine Receptor 4 Antagonist AMD3100 Promotes Cardiac Functional Recovery After Ischemia/Reperfusion Injury via Endothelial Nitric Oxide Synthase-Dependent Mechanism
- CXCR4 blockade augments bone marrow progenitor cell recruitment to the neovasculature and reduces mortality after myocardial infarction
- Challenges in the Translation of Cardiovascular Cell Therapy
- Assessment and Optimization of Cell Engraftment After Transplantation Into the Heart
- Noninvasive Quantification and Optimization of Acute Cell Retention by In Vivo Positron Emission Tomography After Intramyocardial Cardiac-Derived Stem Cell Delivery
- In Vivo Imaging of Stem Cells and Beta Cells Using Direct Cell Labeling and Reporter Gene Methods
- Pilot Trial on Determinants of Progenitor Cell Recruitment to the Infarcted Human Myocardium
- An Imperfect Syllogism: Granulocyte Colony-Stimulating Factor Mobilization and Cardiac Regeneration
- Activation of Epac stimulates integrin-dependent homing of progenitor cells
- Imaging of Gene Expression in Live Pancreatic Islet Cell Lines Using Dual-Isotope SPECT
- Comparison of Imaging Techniques for Tracking Cardiac Stem Cell Therapy
- Dynamic Tracking During Intracoronary Injection of 18F-FDG-Labeled Progenitor Cell Therapy for Acute Myocardial Infarction
- Intramyocardial Implantation of Bone Marrow-Derived Stem Cells Enhances Perfusion in Chronic Myocardial Infarction: Dependency on Initial Perfusion Depth and Follow-up Assessed by Gated Pinhole SPECT
- Imaging Stem Cells Implanted in Infarcted Myocardium
- Tissue Distribution of 18F-FDG-Labeled Peripheral Hematopoietic Stem Cells After Intracoronary Administration in Patients with Myocardial Infarction
- Dynamic Imaging of Allogeneic Mesenchymal Stem Cells Trafficking to Myocardial Infarction
- Monitoring of Bone Marrow Cell Homing Into the Infarcted Human Myocardium