


Abstract
The purpose of this study was to evaluate the effects of 2 anesthetic agents on myocardial perfusion and coronary reserve in rats under resting and stress conditions with small animal PET. Methods: Twenty-four rest/stress studies were performed in 6 rats. Each animal received all 4 possible combinations of anesthetic agents (propofol, isoflurane) and pharmacologic stress agents (dobutamine, adenosine) to increase myocardial perfusion. For each stress or rest study, a 10-min dynamic acquisition was performed in list mode with 185 MBq of 13N-NH3. Data analysis was performed according to a 3-compartment myocardial blood flow model. Pharmacologic stimulation by either dobutamine or adenosine was performed to increase myocardial perfusion. Results: The perfusion values (mean ± SD) for the various experimental conditions were as follows: propofol/dobutamine, 7.8 ± 2.4 mL/g/min (rest, 3.7 ± 0.8 mL/g/min; mean ± SD); isoflurane/dobutamine, 9.3 ± 3.1 mL/g/min (rest, 4.3 ± 1.0 mL/g/min); propofol/adenosine, 6.8 ± 1.7 mL/g/min (rest, 3.2 ± 0.4 mL/g/min); and isoflurane/adenosine, 5.2 ± 1.3 mL/g/min (rest, 3.7 ± 0.7 mL/g/min). All perfusion data showed a significant increase after pharmacologic stimulation relative to baseline (P < 0.05). The coronary reserve (mean ± SD) measured by PET was slightly lower with the combination of isoflurane and adenosine (1.4 ± 0.5) than with propofol and adenosine (2.1 ± 0.5). Conclusion: Noninvasive quantitative measurements of myocardial perfusion in small animals at rest and during stress are feasible using PET. Evaluation of the coronary reserve must take into account the initial state of the anesthetized animal. The coronary reserve could be measured with both anesthetic agents using either dobutamine or adenosine stimulation.
The assessment of new therapeutic approaches for heart disease in animal models often involves complex and invasive techniques. The measurement of myocardial blood flow (MBF) can be useful to assess the impact of drugs or experimental therapies designed to treat the cardiovascular effects of diseases such as diabetes, hypertension, or coronary artery disease. Agents used in general anesthesia can influence myocardial perfusion and coronary reserve and a noninvasive approach to study this phenomenon in small animals would be useful. Interactions between anesthetic agents and new therapeutic drugs could then be tested in a controlled setting. Many of the current techniques used to measure blood flow in small animals are invasive, such as flow probes, or require the sacrifice of the animal (radiolabeled microspheres). In humans, noninvasive approaches for the investigation of myocardial perfusion have been used for many years. Techniques such as SPECT are generally qualitative, but quantitative methods are available for PET.
PET of myocardial perfusion with 13N-ammonia (13N-NH3) has been shown to be accurate in large animal models and in humans (1–3) and is suitable for the characterization and follow-up of heart disease (4). 13N-NH3 PET can assess the status of the coronary arteries and measure noninvasively the coronary perfusion reserve. Although 13N-NH3 in the blood exists mainly in its ammonium ion form 13N-NH4+, it is generally believed that 13N-NH3 crosses the cell membrane by passive diffusion as 13N-NH3. Once inside the cell, 13N-NH3 is metabolically trapped in glutamine, with an uptake relatively proportional to blood flow. This relationship is not linear, but accurate MBF values can be obtained with tracer kinetic modeling.
General anesthesia can affect almost every aspect of the animal circulatory system. Isoflurane acts by depressing the central nervous system but has only mild depressing effects on the heart. Isoflurane decreases blood pressure by reducing peripheral resistance (5–7). Propofol is another widely used anesthetic agent. This drug has a hypotensive effect, produced by an inhibition of the sympathetic nervous system and the baroreflex regulation mechanism. Propofol decreases vascular resistance and arterial pressure without affecting the heart rate (8–11). The purpose of this study was to assess the influence of isoflurane and propofol, 2 common anesthetic drugs, on 13N-NH3 myocardial perfusion at rest and during stress with 2 pharmacologic agents having different modes of action: dobutamine, a positive inotropic agent (12,13), and adenosine, a vasodilator acting on smooth muscle (8,12). Another objective was to demonstrate the feasibility of studying myocardial perfusion repeatedly in the rat in a noninvasive manner using small animal PET.
MATERIALS AND METHODS
In Vivo Studies
The animal experiments were conducted in accordance with the recommendations of the Canadian Council on Animal Care and the in-house Ethics Committee for Animal Experiments. The study was performed on a group of 6 male Sprague-Dawley rats (Charles River Canada) weighing 350–400 g. The experimental protocol was designed in such a way that each rat received all 4 myocardial stress and anesthesia combinations. A separate group of 6 rats was used to study the fraction of 13N-NH3 metabolized at various time intervals.
Metabolite Analysis
The proportion of 13N radioactivity in plasma present as 13N-NH3 was measured in 6 rats with a catheter inserted in a femoral artery. 13N-NH3 (185 MBq) was injected over a period of 25 s in a tail vein, and blood samples were collected (400 μL) at 30, 60, 120, 180, and 300 s from the femoral artery catheter. After deproteinization and centrifugation, the supernatant was sequentially applied to 3 ion-exchange resins (Dowex 1 acetate form, Dowex 50 tris-acetate, and Dowex 50 hydrogen; Sigma-Aldrich) to calculate the fraction of various metabolites using a γ-counter (Cobra II Auto-Gamma; Packard Instruments), as described by Rosenspire et al. (14).
Imaging
Rats were prepared for imaging while under isoflurane (Abbott Laboratories) anesthesia delivered through a nose cone at a concentration of 2.0% volume and 2 L/min oxygen flow. Two venous cannulas (dead volume = 150 μL) were placed in the tail veins, one for administration of 13N-NH3 and the other for injection of the pharmacologic stress agent. In the isoflurane group, the gas was continuously administered at the same dose rate for the remainder of the rest/stress study. For the rats in the propofol group, an infusion of propofol (Abbott Laboratories) was initiated at 0.8 mg/kg/min and isoflurane was progressively discontinued over a period of 5 min. In this group, a minimal waiting period of 15 min occurred after the discontinuation of isoflurane and the beginning of the rest study. For propofol anesthesia, the same vein was used for both anesthesia and administration of 13N-NH3. The physiologic parameters (body temperature, oxygen saturation, heart rate) were monitored and recorded for the entire length of the experiment (Vet/ox 4404; Heska).
Imaging experiments were performed on the Sherbrooke Small Animal PET Scanner (15). This scanner is designed with individual coupling of bismuth germanate crystals to avalanche photodiodes and achieves an in-plane resolution of 2.1 mm, with 2 detector rings and 3 imaging planes at an average distance of 2.75 mm. With a tightly collimated axial field of view, which limits the effects of out-of-field scatter and random coincidences, a high counting rate capability, and good image contrast, this instrument is well suited for dynamic cardiac imaging in small animals.
Three automatic injection pumps (model 210; Lomir Biomedical Inc.) were used for delivery of radiopharmaceuticals and drugs. Before imaging, the heart position was localized with a Doppler probe (0.64 cm [¼ in], 9 MHz; Parks Medical Electronics). During imaging, the animals rested supine on the scanner bed and were kept warm with a heating pad. An interval of 2 wk was introduced between each rest/stress session.
The rest study was initiated once the anesthetized rat was stabilized. The 10-min list-mode dynamic acquisition was begun by intravenous administration of 185 MBq 13N-NH3 over a period of 25 s using an automated injection system. There was a waiting period of ≥10 min after the end of the rest study to allow for 13N decay, after which the stress studies were initiated without moving the animal, using either dobutamine (Novopharm) or adenosine (Fujisawa) stimulation. Dobutamine administration was performed with 3 stepwise increases in the infusion rate to progressively increase the heart rate (5 μg/kg/min for 2 min, 10 μg/kg/min for 2 min, 20 μg/kg/min for 9 min). The injection of 13N-NH3 (185 MBq) occurred 5 min after the last dobutamine plateau was reached. The infusion of dobutamine was stopped 5 min after the beginning of the 10-min acquisition. Adenosine stimulation was effected by a constant infusion rate (140 μg/kg/min), which started 4 min before the injection of 13N-NH3 and was stopped 2 min later.
Data Analysis
A dynamic series of 22 frames was sorted out from the list-mode data, using the following sequence: 12 × 5 s, 8 × 30 s, 2 × 150 s. Image planes were reconstructed on a 128 × 128 matrix with a 0.475 × 0.475 mm pixel size using the maximum likelihood expectation maximization algorithm with 15 iterations (16). Tissue time-activity curves were generated from a region of interest (ROI) drawn manually on the myocardium of the best frame of the rest study and then copied to the remaining frames of the whole rest/stress images. The input curve was obtained from a ROI drawn on the left ventricular cavity and was used with and without corrections for metabolites. This correction was applied using a monoexponential equation fitted to the fraction of 13N present as metabolites at each time interval (Table 1). The blood-to-myocardium spillover component was modeled using the noncorrected curve.
Fraction of 13N Activity Present as 13N-NH3 in Rat Blood
A 3-compartment kinetic model was used for the computation of blood flow as shown in the equations below, using the blood and tissue curves derived from PET data (Fig. 1) (17,18):



 where Ct and Cp are the tissue and plasma time-activity curves; K1, k2, and k3 are the rate constants between the 3 compartments (blood, nonmetabolized, and metabolized tissue activity); v is the blood volume that represents the fraction of blood in the tissue ROI; and ρ = 1.04 g/mL is the tissue density. The MBF or perfusion is given by K1 (mL/g/min).
where Ct and Cp are the tissue and plasma time-activity curves; K1, k2, and k3 are the rate constants between the 3 compartments (blood, nonmetabolized, and metabolized tissue activity); v is the blood volume that represents the fraction of blood in the tissue ROI; and ρ = 1.04 g/mL is the tissue density. The MBF or perfusion is given by K1 (mL/g/min).

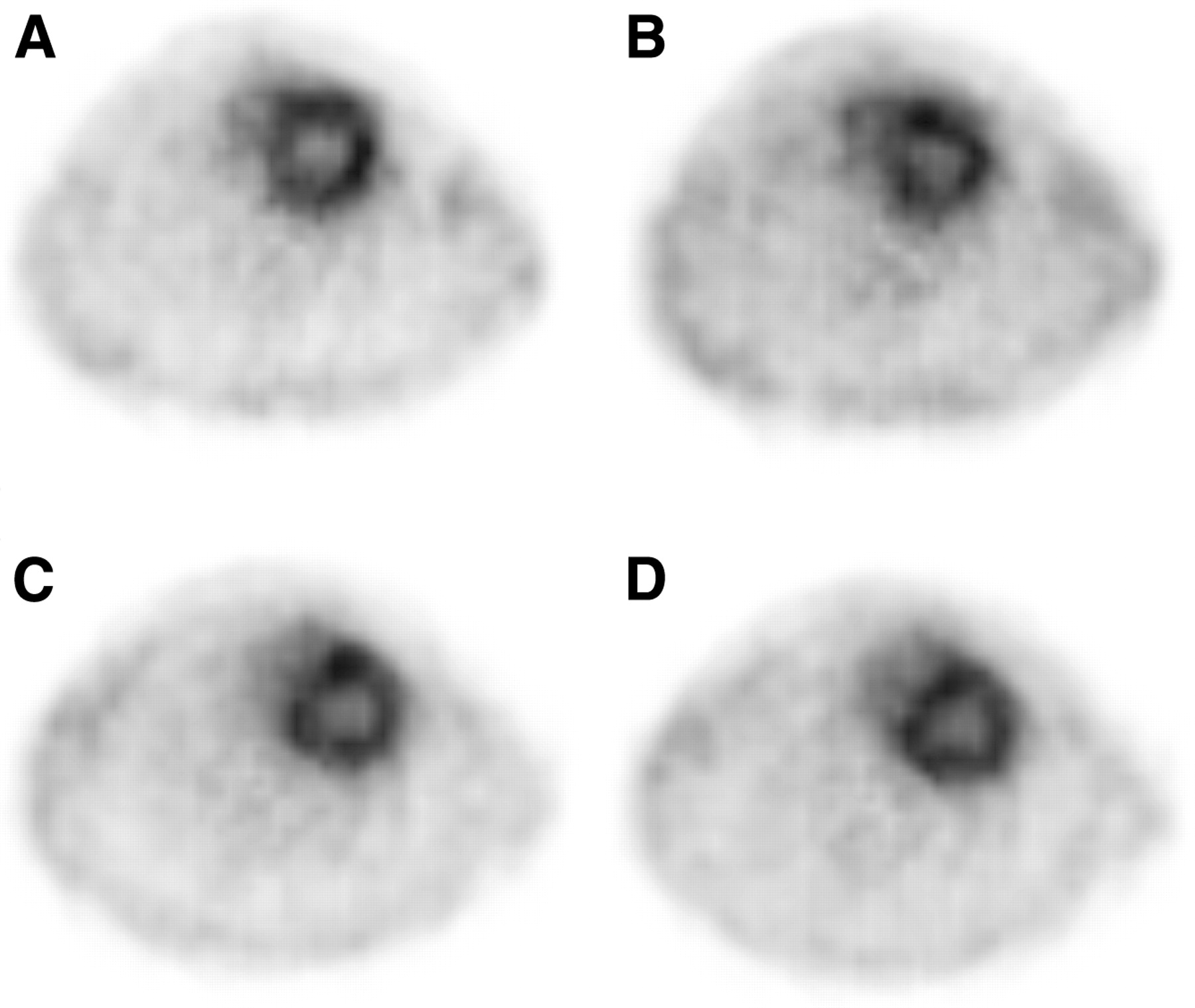
Perfusion images obtained at rest and stress with 13N-NH3. Top images were obtained at rest (A) and after dobutamine pharmacologic stimulation (B). Bottom images were obtained at rest (C) and after adenosine administration (D). There is a clear separation of left ventricular cavity from myocardium.
Statistical Analysis
Statistical analyses were performed with the PRISM 3.0 software package (GraphPad Software). The curves representing the proportion of 13N present in the blood as 13N-NH3 were fitted with a monoexponential equation using the data points from all animals at each time interval. The half-life and the SE were calculated as ln(2)/K from the rate constant K provided by the PRISM curve-fitting package. For each time point, the mean ± SEM of the fraction of radioactivity remaining as 13N-NH3 were also calculated (Table 1). Comparisons between stress and rest under the different conditions of anesthesia were performed using a paired 1-tailed Student t test. The mean ± SD were calculated for MBF and heart rate values for each experimental group (Table 2). The comparison between resting flow values was conducted with a 2-tailed paired Student t test.
Hemodynamic Responses Under Anesthesia
RESULTS
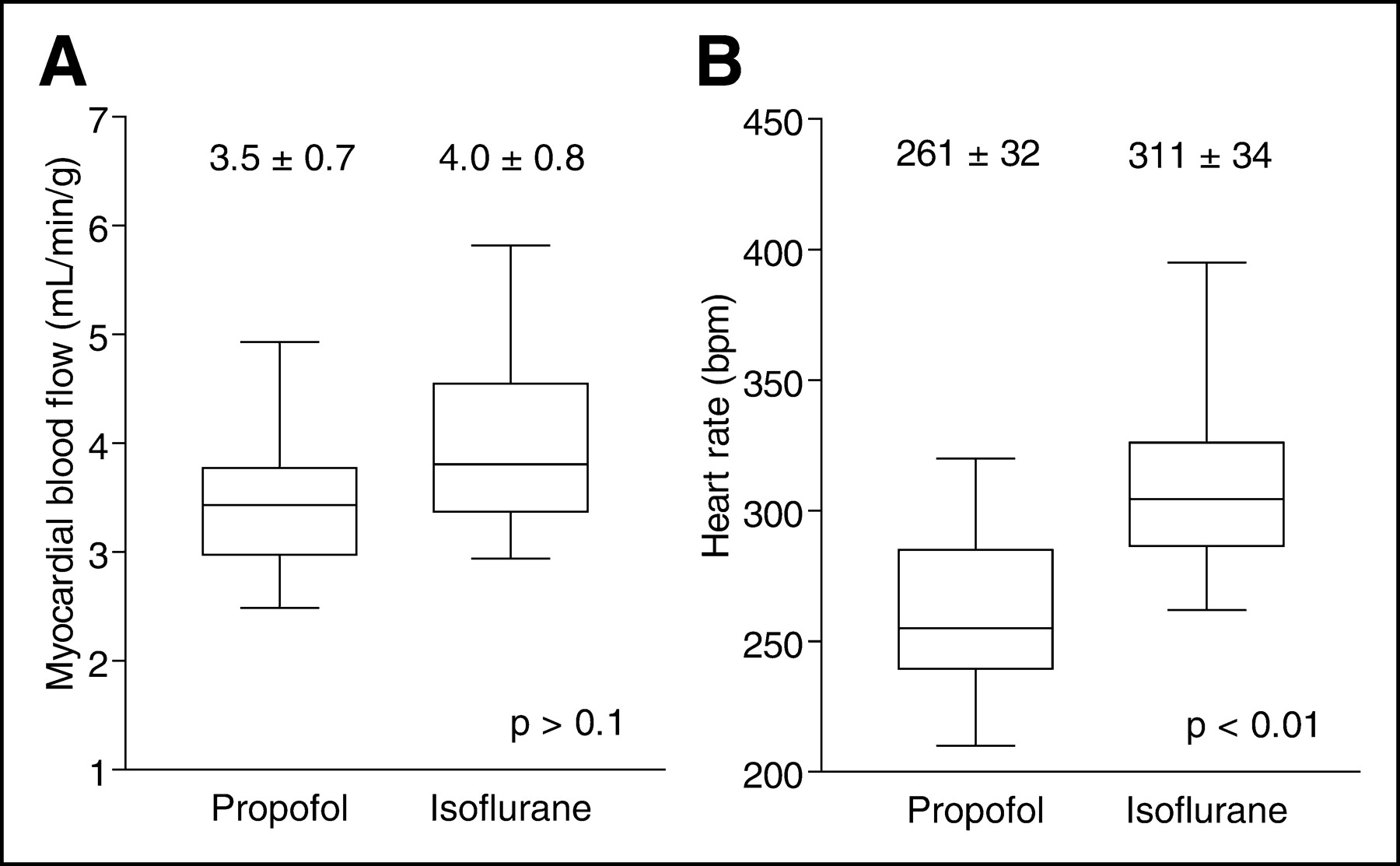
The animal PET scanner resolution allowed for a clear delineation of the blood cavity from the myocardial wall (Fig. 1). The model used to calculate perfusion included a blood activity curve (ammonia + metabolites) and a myocardial wall activity curve (Fig. 2). 13N-NH3 was metabolized rapidly (Table 1), with a plasma half-life of 59 ± 9 s. In general, correction for metabolites in rat blood increased myocardial blood perfusion values slightly (Table 2). The perfusion values obtained in 6 rats before dobutamine and adenosine stress were averaged for each anesthetic agent, providing 12 resting studies to compare perfusion values at rest under isoflurane anesthesia with those obtained under propofol. At rest, there was a trend for slightly higher average myocardial perfusion values with isoflurane (4.0 ± 0.8 mL/g/min [mean ± SD]) compared with propofol (3.5 ± 0.7 mL/g/min), but this difference did not reach statistical significance (P = 0.1; Fig. 3A). The heart rates observed at rest with isoflurane (311 ± 34 beats per minute [bpm]) were higher than those obtained under propofol anesthesia (261 ± 32 bpm; P < 0.001; Fig. 3B). Overall, the blood volume (v) parameter estimated from the model of Musik et al. (17) was 0.59 ± 0.09 (mean ± SD).

Tissue and blood activity curves from 13N-NH3 study in rat. Blood 13N represents curve obtained without correction for metabolites. Blood 13N-ammonia represents curve after correction for metabolites.
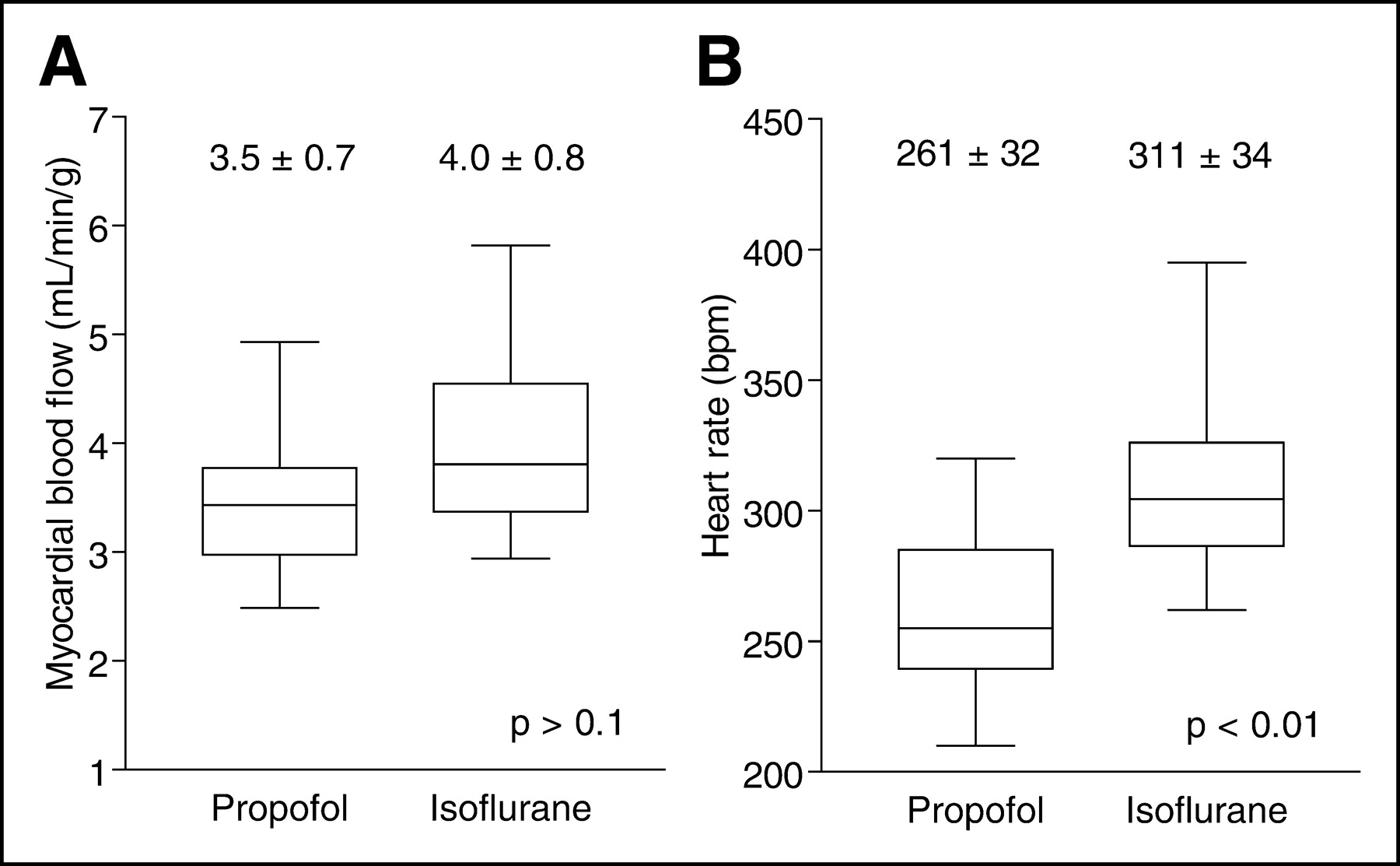
Box and whisker plots of effect of anesthesia on rat heart at rest. Values are mean ± SD. (A) Coronary perfusion. Paired Student t test, 2 tails, n = 12, t = 1.61, P = 0.13. (B) Heart rate in beats per minute (bpm). Paired Student t test, 2 tails, n = 12, t = 5.58, P < 0.01.
The increase in MBF after pharmacologic stress relative to the resting state under anesthesia was clearly demonstrated (Fig. 4), with an average increase of MBF by 3.5 mL/g/min during stress. The mean rest and stress heart rate, MBF, and coronary reserve are shown on Table 1. Both agents induced a significant increase in MBF. The greatest difference in heart rate between rest and stress was observed with dobutamine, with an increase of 57 bpm under propofol and 53 bpm under isoflurane anesthesia (P < 0.05). Adenosine administered during propofol anesthesia also increased the heart rate but to a lesser extent than dobutamine. The combination of adenosine and isoflurane decreased the heart rate relative to the resting state (Fig. 4B).

Box and whisker plots of effect of stimulating agent on MBF and heart rate while animals were under anesthesia. (A) Dobutamine stimulation. (B) Adenosine stimulation. *Paired Student t test, 1 tail, n = 6 for each group, P < 0.05 rest vs. stress.
The coronary vasodilatation reserve (CVR = MBF at stress/MBF at rest) for the rats was 2.2 ± 0.7 (n = 12) with propofol and 1.8 ± 0.7 with isoflurane (n = 12). Using data from both propofol and isoflurane anesthesia, the perfusion reserve was 2.2 ± 0.8 (n = 12) with dobutamine and 1.8 ± 0.6 (n = 12) with adenosine (Fig. 5). The differences between CVR values did not reach statistical significance (2-way ANOVA, P > 0.10).

Box and whisker plots of CVR in rat heart after stress stimulation under anesthesia.
DISCUSSION
This study demonstrates that measurement of myocardial perfusion under conditions of rest and stress, assessed by a small animal PET scanner using 13N-NH3 in the rat, is feasible and provides consistent, reproducible results. Although rats degrade NH3 much more rapidly than dogs and humans (14), the corrections applied to take into account the presence of metabolites had a relatively limited impact on absolute MBF measurements.
The perfusion measurements were performed using 1 of 2 possible anesthetic agents at rest and after the administration of either adenosine or dobutamine. Our results show similar myocardial perfusion values at rest under isoflurane or propofol anesthesia, although there was a trend for slightly higher values with isoflurane. Both anesthetics are known to produce a mild coronary vasodilatation (19–21). Isoflurane induces a decrease in peripheral resistance and blood pressure. To compensate for the drop in systolic volume the heart rate is increased, resulting in an unchanged cardiac output (5–7). In vivo, when the hemodynamic conditions are controlled, isoflurane is able to increase the coronary blood flow but to a lesser extent than the vasodilator adenosine (19). Propofol acts on the cardiovascular system by decreasing both total vascular resistance and cardiac output, resulting in a decrease in arterial blood pressure (8–11,21,22). At rest, propofol does not seem to interfere with the heart rate, whereas isoflurane increases it. Due to concomitant changes in preload, systemic vascular resistance, baroreflex activity, and central nervous system activity, the precise effects of propofol on intrinsic myocardial contractility are difficult to assess in vivo. With propofol, changes in heart rate and cardiac output are usually transient and insignificant in healthy patients (8–11,23). Among reported cardiovascular adverse reactions with propofol, bradycardia is seldom observed, occurring in <3% of subjects (24). Apart from this potential side effect, a reduction in heart rate is not usually observed after propofol administration. With isoflurane, there is a dose-dependent relationship with the increase in heart rate (25). Isoflurane depresses deep breathing, so increased doses of isoflurane may reduce the tidal volume without affecting the respiration rate. This reduction in breathing efficiency brings about a decrease in oxygenation. The most likely hypothesis is that increased heart rate compensates for the drop in systolic volume and hypercapnia to maintain the cardiac output and tissue oxygenation at various levels of anesthesia (5–7,19,26). There was a lower heart rate in animals anesthetized with propofol compared with isoflurane. This could be due to bradycardia induced by propofol or, more likely, to an increase in the baseline heart rate induced by isoflurane.
Our rest/stress model demonstrates that dobutamine- and adenosine-mediated vasodilatation of the coronary arteries can be detected noninvasively by PET in small animals. These 2 compounds act by different mechanisms. Dobutamine is a positive inotropic agent stimulating the β-adrenergic receptors, which increases heart rate and strength of contraction (27). Adenosine, on the other hand, initiates vasodilatation by acting on the adenosine A2A receptors, which are found on the smooth muscle and endothelial cells of the coronary walls (28). The differences observed between the rest and the stress conditions were statistically significant for each of the pharmacologic stress-anesthetic combinations.
During the rest/stress study with the 2 anesthetic agents, stimulation with dobutamine increased the heart rate and the coronary perfusion, as expected (13). A similar response was observed when propofol was used in conjunction with adenosine (8). However, when isoflurane was combined with adenosine we observed a decrease in heart rate despite an increase in myocardial perfusion. A possible explanation for the effects of adenosine in these conditions would be an inhibition of the atrioventricular nodal conduction, bringing about a decrease in heart rate (29). With propofol, the heart rate was lower and this effect was not observed.
In healthy humans, the CVR is around 4 (30). In the rat, however, a lower value of 2.1 was obtained. The CVR values we measured in the rat were similar to what has been reported by others (31), using simple pulsing contrast echocardiography and radiolabeled microspheres in a stress/rest dypiridamole study. Three possible hypotheses are suggested to explain this difference in CVR:
The perfusion measurements underestimate MBF at high values.
The vasodilatation induced by the pharmacologic stress was not maximal or the rat was under anesthesia, increasing MBF at rest.
MBF in the rat cannot be increased >2.0 × baseline.
We tend to favor the second hypothesis, although whether anesthesia increases baseline blood flow in this model is debatable, given the absence of a nonsedated control group. At rest, the MBF tended to be slightly higher under isoflurane compared with propofol anesthesia, suggesting increased perfusion compared with a resting state obtained without anesthesia in a nonstimulated rat. For obvious reasons, PET cannot be reliably conducted in rats without sufficient sedation or anesthesia. Even with immobilization devices, the severe stress induced by such a procedure would probably increase blood flow relative to a true resting state. The resting perfusion values observed in this study are in agreement with those obtained with microspheres, which are in the range of 3.83–7.89 mL/min/g in awake animals (32–34). The underestimation of myocardial activity in the tissue ROI due to partial-volume averaging effects is unlikely to be significantly different between rest and stress and this effect is negated when calculating the CVR. The blood-to-tissue spillover fraction, another consequence of partial-volume averaging, is taken into consideration by the perfusion model (17,18). In the later time frames, when the myocardial wall activity is high, the tissue-to-blood spillover could affect the input curve. However, corrections for the presence of metabolites, which have a much more significant impact on the latter portion of the input curve (Fig. 2), have a relatively minor impact on MBF values, as shown on Table 2. The effect of myocardium-to-blood spillover, which also occurs only at later time frames, is therefore probably not significant.
We did not establish a full dose-response relationship between the dose of the pharmacologic agents and the measured flow increase by PET. Thus, it is possible that higher perfusion values could have been obtained using higher doses of dobutamine or adenosine. We used doses that were extrapolated from typical human doses during pharmacologic stress testing. The dose of dobutamine of 20 μg/kg/min was slightly lower than the maximal dose of 30 μg/kg/min commonly used in pharmacologic stress testing. The response to adenosine, though comparable to intracoronary papaverine in humans at a dose of 0.14 mg/kg/min (35), could be more pronounced at higher doses in rats. Thus, interspecies differences and submaximal doses could account for the lower CVR values observed in this study.
Our MBF values are in the same range as published results obtained with microspheres. The measurement of rat MBF by the microsphere method yields variable results depending on the protocol used: conscious MBF (3.83–7.89 mL/min/g) (32–34), under isoflurane MBF (5.04–8.58 mL/min/g) (33,36), with ketamine/xylazine MBF (1.52–5.17 mL/min/g) (33,37), and with pentobarbital MBF (2.79–6.64 mL/min/g) (38,39). Many other factors influence the MBF when measured by microspheres, such as the size of the microspheres, the rate of infusion, and the presence or absence of ventilation during anesthesia. To correlate the 13N-NH3 PET and microsphere blood flow measurements, the experimental conditions should be the same for both methods. The lack of a gold standard to validate the absolute flow measurements obtained with PET is a limitation of the current study.
The high flow values observed in rats at baseline and after pharmacologic stimulation are in the range where the extraction of many perfusion tracers is reduced. Though 13N-NH3 uptake continues to increase significantly at flow values >4 mL/min/g (2), the uptake of 99mTc-sestamibi, for example, reaches a plateau at flow levels in the range of 2–2.5 mL/min/g (40). This may limit the applicability of current 99mTc tracers to study alterations in blood flow in small animals with micro-SPECT instruments. For this purpose, small animal PET has the advantage of allowing absolute flow measurements using tracer kinetic modeling to correct for the nonlinearity between 13N-NH3 uptake and blood flow.
CONCLUSION
Noninvasive small animal PET rest/stress studies in the rat are feasible and reproducible. Our results suggest that propofol and isoflurane anesthesia produce similar perfusion values under “resting” conditions with a trend toward slightly higher values with isoflurane. It is possible to measure noninvasively the increase in perfusion induced by pharmacologic stress testing with either dobutamine or adenosine in rats using small animal PET. Several repeated experiments can be performed in the same animal. Although the accuracy of absolute flow measurements obtained with small animal PET remains to be validated against a gold standard, the measured values were reproducible and a significant increase was measured using pharmacologic stimulation.
Noninvasive PET rest/stress studies in small animals open new possibilities for the investigation of innovative therapeutic approaches designed to improve myocardial perfusion in disease conditions such as hypertension, diabetes, and coronary artery disease. The ability to study the effects of new drugs and disease states on MBF using the same animal under various experimental conditions may offer interesting insights, as long as the effects of anesthesia on myocardial perfusion are taken into consideration.
Acknowledgments
The authors thank the technical and cyclotron staff of the Metabolic and Functional Imaging Center for their assistance. This work was supported by Canadian Institutes of Health Research (CIHR) grant MOP-15348 and a scholarship from the Clinician-Scientist Award Program of the CIHR.
Footnotes
Received Feb. 2004; revision accepted July 14, 2004.
For correspondence or reprints contact: François Bénard, MD, Département de médecine nucléaire et radiobiologie, Faculté de médecine, Université de Sherbrooke, 3001 12th Ave. N., Sherbrooke, Québec, Canada, J1H 5N4.
E-mail: francois.benard{at}usherbrooke.ca
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Comparison of 18F-Labeled Fluoroalkylphosphonium Cations with 13N-NH3 for PET Myocardial Perfusion Imaging
- Initial Characterization of an 18F-Labeled Myocardial Perfusion Tracer
- In Vivo Characterization of Murine Myocardial Perfusion With Myocardial Contrast Echocardiography: Validation and Application in Nitric Oxide Synthase 3 Deficient Mice
- Noninvasive Measurement of Cardiovascular Function in Mice with High-Temporal-Resolution Small-Animal PET
- Assessment of Myocardial Blood Flow Using 15O-Water and 1-11C-Acetate in Rats with Small-Animal PET
- Voluntary physical exercise and coronary flow velocity reserve: a transthoracic colour Doppler echocardiography study in spontaneously hypertensive rats
- Functional and Morphologic Imaging of Coronary Atherosclerosis in Living Mice Using High-Resolution Color Doppler Echocardiography and Ultrasound Biomicroscopy