


Abstract
To track neural stem cells transfected with reporter gene, perpetual stem cell transgene expression is required. Referring to the knowledge about epigenetic modulation, we succeeded in reversing the silencing of sodium/sodide symporter (hNIS) transgenes transfected into human neural stem (HB1.F3) cells. Methods: hNIS and hygromycin resistance gene were linked with IRES (Internal Ribosome Entry Site) under control of cytomegalovirus promoter, and this construct was transfected into HB1.F3 cells to yield the F3-NIS cell lines. hNIS transgene expression was examined by 125I uptake and reverse transcriptase polymerase chain reaction (RT-PCR). The iodide uptake of F3-NIS III cells was initially higher by up to 12.9-fold than that of nontransfected HB1.F3 cells. However, repeated passage gradually silenced hNIS expression. The recovery of hNIS transgene expression by demethylating agent (5-azacytidine) or histone deacetylase inhibitor (trichostatin A; TSA) treatment was investigated. Results: As hNIS transgene was gradually silenced in F3-NIS III cells, after the eighth passage its iodide uptake was 1.9-fold higher than that of nontransfected HB1.F3 cells. 5-Azacytidine treatment (up to 40 μmol/L) for 24 h in F3-NIS III cells increased iodide uptake and hNIS messenger RNA (mRNA) 1.8- and 1.9-fold versus nontreated F3-NIS cells, respectively. Moreover, after TSA treatment (up to 62.5 nmol/L) for 24 h, iodide uptake and hNIS mRNA in F3-NIS III cells increased 36- and 1.9-fold versus nontreated F3-NIS III cells, respectively. The synergistic effect of demethylation and histone deacetylation inhibition was significant at high-dose 5-azacytidine and low-dose of TSA treatment. After treating F3-NIS III cells in vitro for 24 h with 62.5 nmol/L TSA, the cells were implanted into BALB/c nude mice. The TSA-treated F3-NIS III cells were clearly visible on γ-camera imaging using 99mTc-pertechnetate as compared with F3-NIS III cells not treated with TSA. Conclusion: These results suggest that 2 well-known mechanisms of epigenetic modulation synergistically are involved in silencing reporter hNIS transgene in a neural stem cell line. Transgene silencing was reversed using demethylation and histone deacetylation inhibition. We conclude that silenced reporter transgenes once successfully expressed in stem cells might be awakened by pharmacologic treatment before infusion to track stem cells in vivo.
- neural stem cell
- human sodium/iodide symporter
- gene silencing
- promoter methylation
- histone deacetylation
Neural stem cell therapy has been tried to treat neurodegenerative diseases, stroke, and brain injuries in human trials and in experimental animals (1,2). Stem cell tracking is essential for studying the migration, localization, and differentiation of transplanted stem cells in vivo. Although reporter genes such as LacZ or GFP have been used to track transplanted stem cells in animal experiments, biopsy or subject sacrifice was required and sequential measurement was not possible in the same subject. To track the transplanted stem cells repeatedly, molecular imaging is required to enable noninvasive examination of the migration and possibly proliferation of stem cells in vivo.
Three methods can be used to noninvasively track stem cells in vivo: MRI, nuclear imaging, and in vivo optical imaging (3). Though MRI has the advantages of a very high spatial resolution and allows repetitive measurements in the same animal, its major shortcoming is the limited number of molecular probes. Optical imaging is relatively inexpensive and can be performed repetitively in the same animal but is largely limited to the imaging of small animals. Nuclear imaging offers the possibility of monitoring the expression of reporter genes in vivo in humans as well as in small animals. Human sodium/iodide symporter (hNIS) was proposed as a good candidate as a reporter gene (4,5) because we have already well-known radioactive probes—that is, radioiodines and 99mTc-pertechnetate. Furthermore, the metabolism and clearance of these substrates in normal subjects are well understood well.
The expression of genes in cancer can be regulated by epigenetic modulation such as DNA methylation or histone deacetylation (6). Nucleosomes, composed of 4 pairs of histones and a short length of DNA, are the sites of DNA methylation and histone deacetylation. Acetylation of the N-terminal lysine residue of histone protein opens the nucleosome structure, which favors transcription. On the contrary, if 5-methyl-cytosine binding protein binds to “methylated” CpG sites and recruits histone deacetylase (HDAC), nucleosome becomes compact to prevent transcription. Thus, DNA methylation and histone deactylation result in silencing the genes, and demethylation and HDAC inhibition can reverse the gene silencing. Reporter genes transfected to the stem cells should express their transgene activity throughout their progeny for stem cell tracking; however, transfected genes are sometimes silenced in vivo and even in vitro before injection. We need the methods to reverse the silencing of a reporter transgene such as hNIS.
In this study, we produced stable HB1.F3 human neural stem cells transfected with hNIS whose hNIS expression was prominent at first but decreased after passage. We report that this transgene silencing can be reversed by DNA demethylation and histone deacetylation inhibition, which implies that epigenetic modulations such as DNA methylation and histone deacetylation are important in reporter transgene silencing.
MATERIALS AND METHODS
Neural Stem Cell Culture
HB1.F3 cells were grown in Dulbecco’s modified Eagle medium (DMEM) (Gibco BRL) as reported earlier (7–9). HB1.F3 cells were originally isolated from embryonic human brains at 15 wk of gestation and were immortalized by retroviral transduction with v-myc oncogene.
Production of Vector and hNIS Transfected Cell Line
hNIS complementary DNA (cDNA) and the hygromycin resistance gene were inserted into plasmid pIRES (where IRES is Internal Ribosome Entry Site) (Clontech). The hNIS gene was released by Xho I and EcoRI restriction enzymes from the plasmid (kindly provided by Dr. Sissy M. Jhiang) and inserted to MCS (multicloning site) A of pIRES vector. The hygromycin resistance gene from pcDNA3.1 (Invitrogen) was prepared by polymerase chain reaction (PCR), and PCR products were released by Xba I and Not I and then inserted into the MCS B of pIRES vector to make pIRES-hNIS/HYG vector.
HB1.F3 cells were transfected with pIRES-hNIS/HYG vector using a lipofectamine plus reagent (Invitrogen). Selection was performed using 150–250 μg/mL hygromycin B in DMEM containing 5% fetal bovine serum (FBS) and 5% horse serum (HS) for 2 wk beginning the day after transfection. Three colonies were selected with hygromycin B, which are referred to as F3-NIS I, II, and III.
In Vitro 125I Uptake Assay in F3-hNIS Neural Stem Cells
F3-NIS cells were plated at a cell density of 1 × 105 cells per well in a 24-well plate and cultured with DMEM containing 5% FBS and 5% HS. The 125I uptake assay was performed using the methods reported by Min et al. (4) and Shin et al. (5). Briefly, 125I uptake was determined by incubating cells with 500 μL of Hanks’ balanced salt solution containing 0.5% bovine serum albumin and 10 mmol/L 2-(4-[2-hydroxyethyl]-1-piperazinyl)ethanesulfonic acid-NaOH with 3.7 kBq (0.1 μCi) carrier-free Na125I and 10 μmol/L NaI (specific activity, 740 MBq/mmol) for 30 min. After incubation the cells were then detached with 500 μL of 0.2% sodium dodecyl sulfate, and radioactivity was measured using a Cobra II γ-counter (Canbera Packard). The radioactivity of cells was normalized with total proteins, which was measured using a bicinchoninic acid Protein Assay Kit (Pierce). For the inhibition assay, radioiodine uptake was inhibited by pretreatment with 50 μmol/L potassium perchlorate for 30 min.
Treatment Effect of 5-Azacytidine and Trichostatin A (TSA)
After a 24-h incubation, cells were treated in quadruplicate with 5-azacytidine (Sigma) at 2.5, 5, 10, 20, or 40 μmol/L. Cells were also treated in quadruplicate with TSA (Sigma) at 25, 37.5, 50, or 62.5 nmol/L. Twenty-four hours later, the clones were assessed for NIS activity using the same procedure of 125I uptake per milligram of protein.
To measure the effect of the combined treatment of 5-azacytidine and TSA of hNIS in F3-NIS III cells, 5-azacytidine (0, 2.5, 5, 10, or 20 μmol/L) and TSA (12.5 and 25 nmol/L) treatment lasted for 24 h.
Reverse Transcriptase PCR (RT-PCR) Analysis for hNIS
Total RNA was prepared from F3-NIS III cells using Trozol reagent (Invitrogen) following the manufacturer’s protocol. cDNA was produced using Moloney murine leukemia virus (M-MLV) reverse transcriptase (Invitrogen). PCR was then performed with the hNIS forward and reverse primers using a GeneAmp PCR system (Applied Biosystems). β-Actin was amplified as a control using the same reaction solution but with β-actin primer. The amplified products obtained were analyzed by ethidium bromide–stained agarose gel electrophoresis. Results were interpreted using Tina 2.1 (Raytest). The expression value of the NIS gene was calculated by dividing the intensity of bands for NIS with that of β-actin for normalization.
Scintigraphic Imaging of hNIS Gene in TSA-Treated F3-NIS III Cells Implanted into Nude Mice
Stem cell transplantation was performed in male BALB/c nu/nu mice by subcutaneous injection as follows: 1 × 108 F3-NIS III cells treated with 62.5 nmol/L TSA in vitro, left thigh; and 1 × 108 nontreated F3-NIS III cells, right thigh. Three hours later, mice were anesthetized with an intraperitoneal injection of ketamine (53 mg/kg) and xylazine (12 mg/kg). Thirty minutes after injecting 11.1 MBq (300 μCi) of 99mTc-pertechnetate intraperitoneally per animal, the mice were placed in a spread prone position and scanned with an ON 410 γ-camera (Ohio Nuclear) with a pinhole collimator.
RESULTS
Changes (Silencing) of hNIS Gene Expression in F3-NIS III Cells by Cell Passage
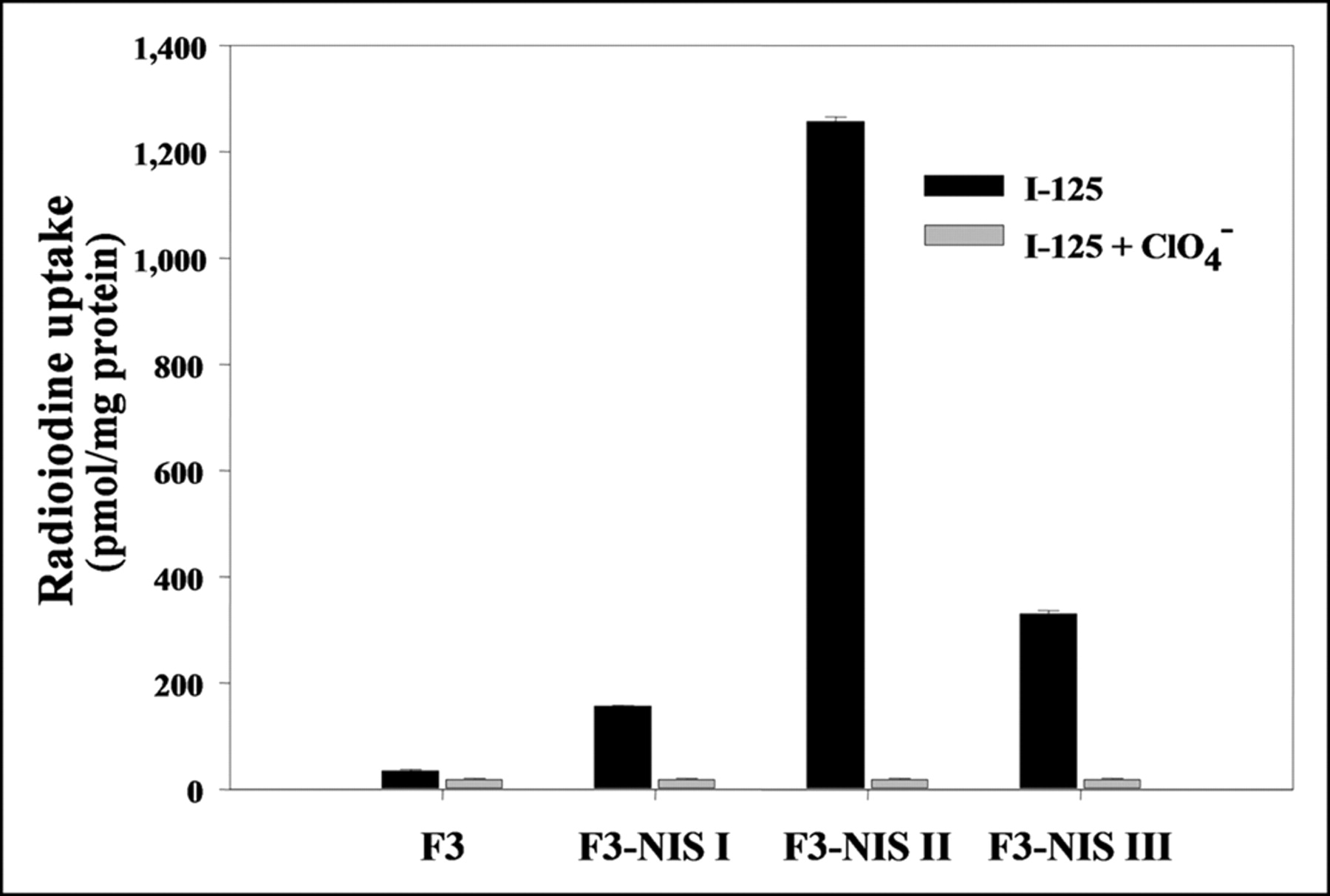
After transfecting HB1.F3 cells with pIRES-hNIS/HYG reporter vector using liposomes and selecting with hygromycin B, we obtained 3 colonies that stably expressed the hNIS gene. 125I uptake in HB1.F3 I, II, and III cells were 4.5-fold (156.6 ± 1.4 pmol/mg protein), 36.5-fold (1,257.2 ± 8.2 pmol/mg protein), and 9.5-fold (330.6 ± 5.9 pmol/mg protein) higher than that in HB1.F3 cells (34.8 ± 2.9 pmol/mg protein), respectively (Fig. 1). In all cell lines, 125I uptake was completely inhibited by 30 μmol/L potassium perchlorate, a competitive inhibitor of NIS.
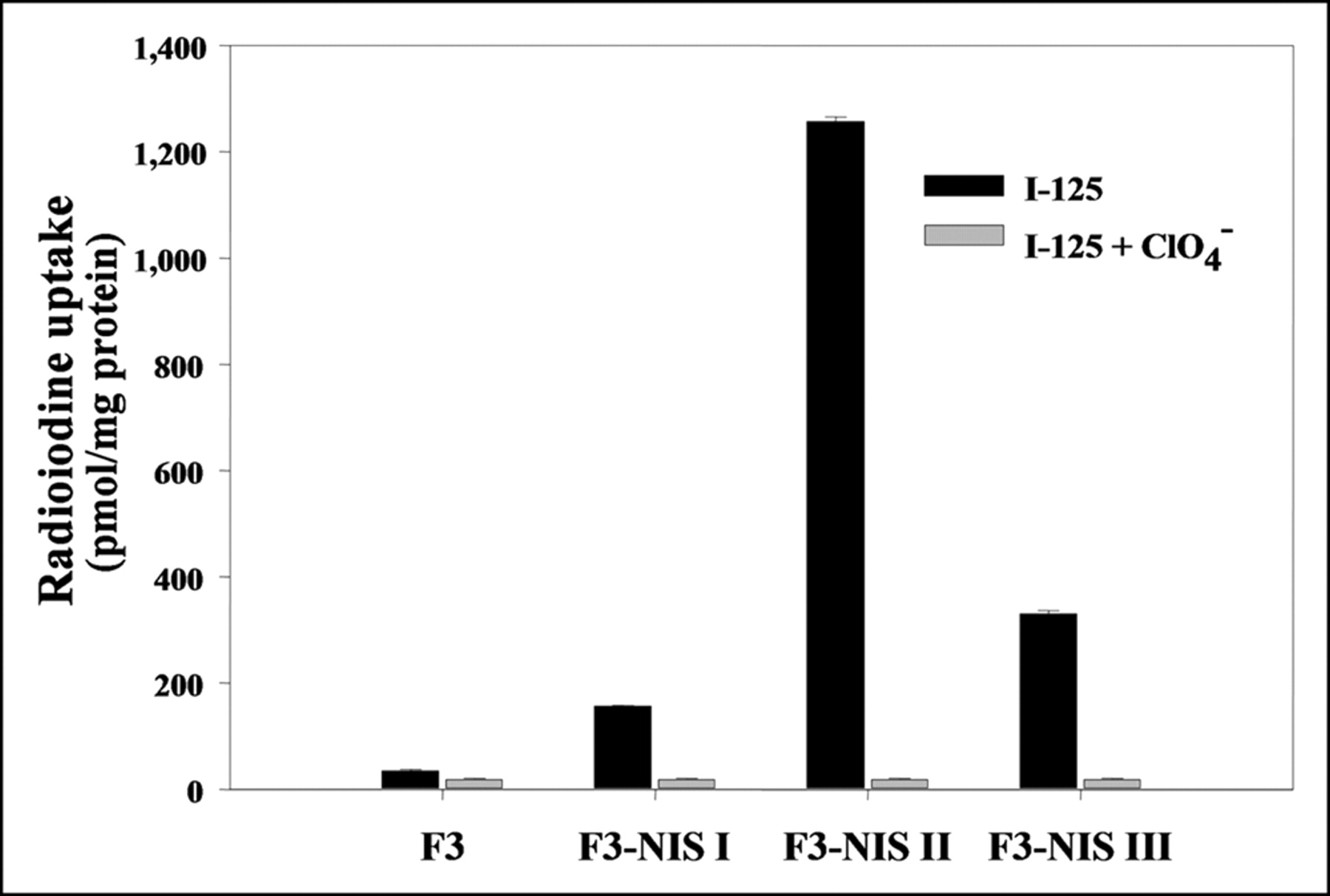
Results of 125I uptake assay in 3 clones of hNIS transfected HB1.F3 neuronal stem cell lines. 125I uptake increased up to 36-fold and was completely inhibited by KClO4. Results are means ± SD (n = 4).
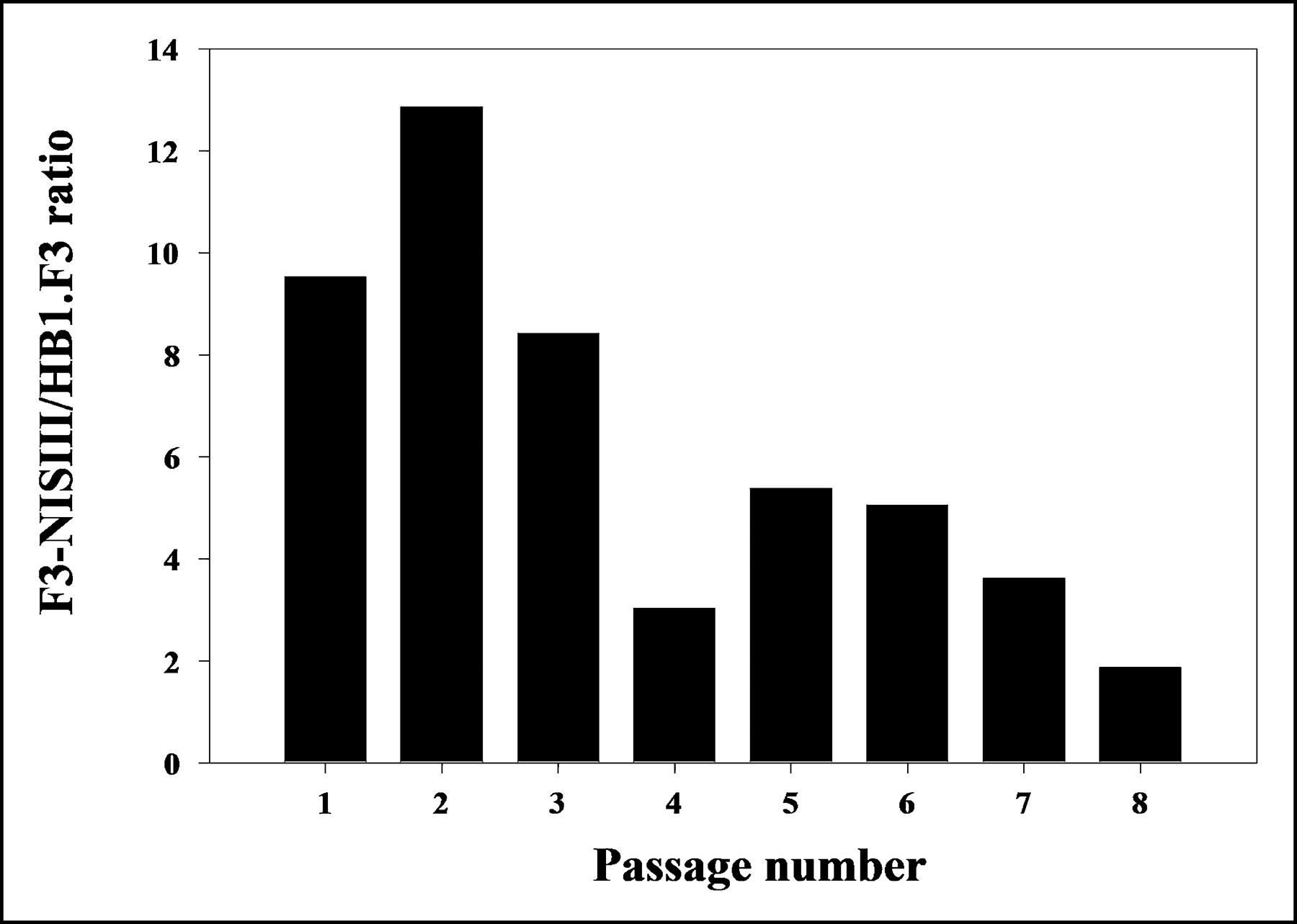
In the cell-passage hNIS activity experiment, 125I uptake in F3-NIS III cells was initially 12.9-fold higher than that in HB1.F3 cells. 125I uptake in F3-NIS III cells gradually decreased with cell passage. At the eighth passage, the 125I uptake of F3-NIS III cells was only 1.9-fold higher than that of HB1.F3 cells (Fig. 2).
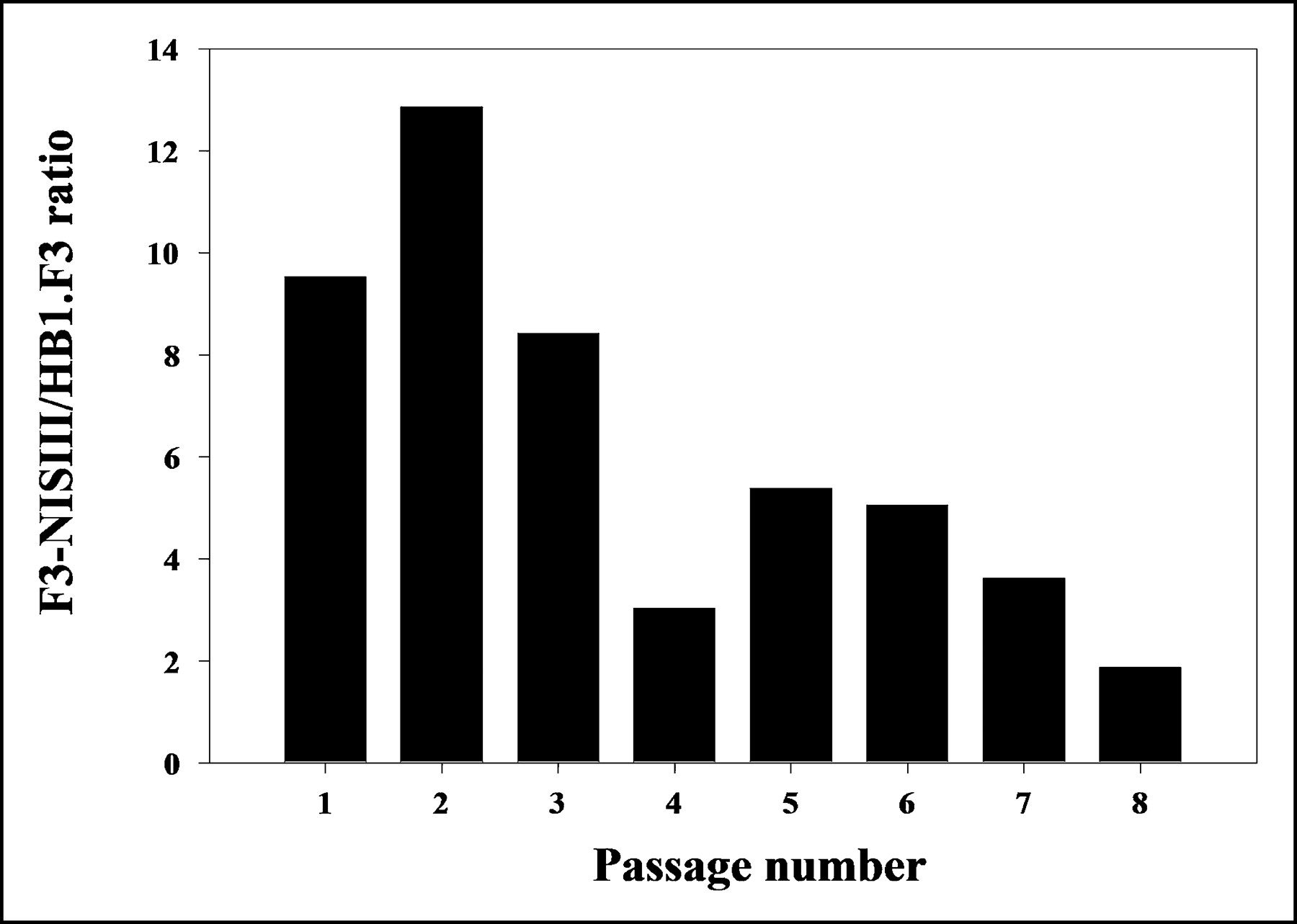
125I uptake in F3-NIS III cells (expressed as F3-NIS III/HB1.F3 ratio) according to passage number. Results are means ± SD (n = 4).
Effect of Demethylation (5-Azacytidine) or HDAC Inhibition (TSA) on 125I Uptake by hNIS Gene Expression
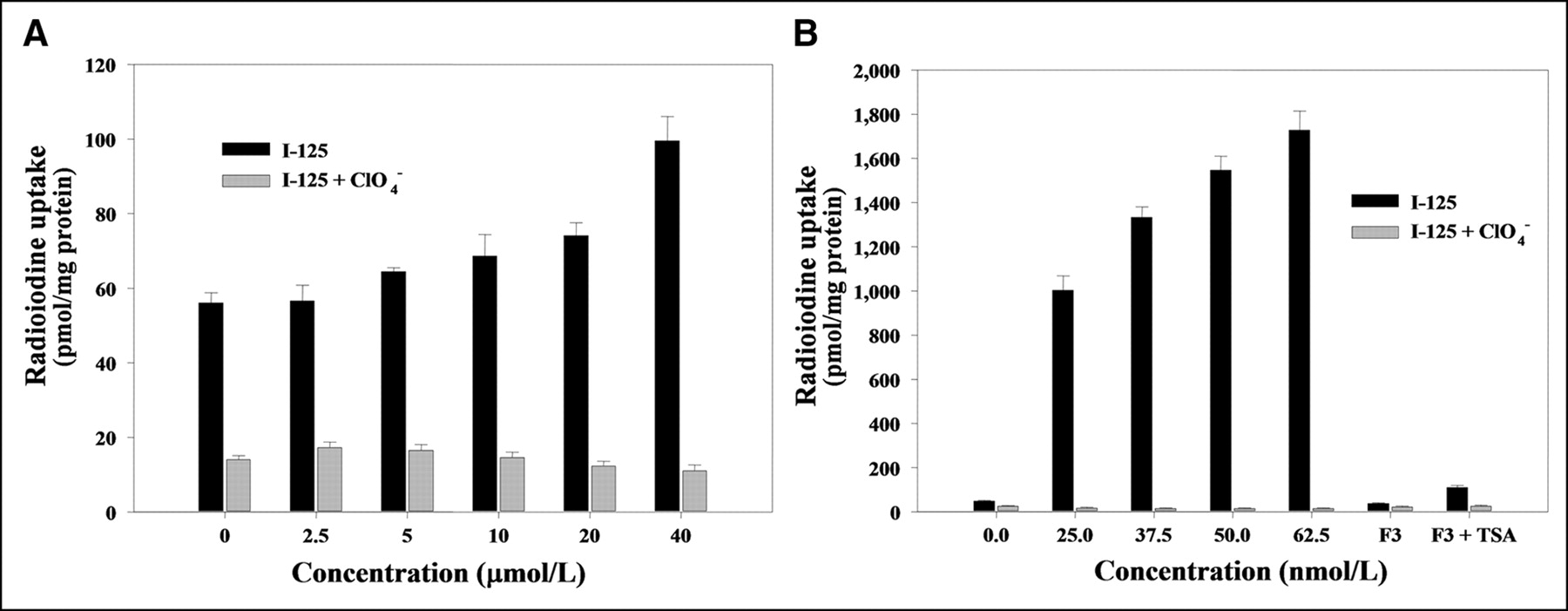
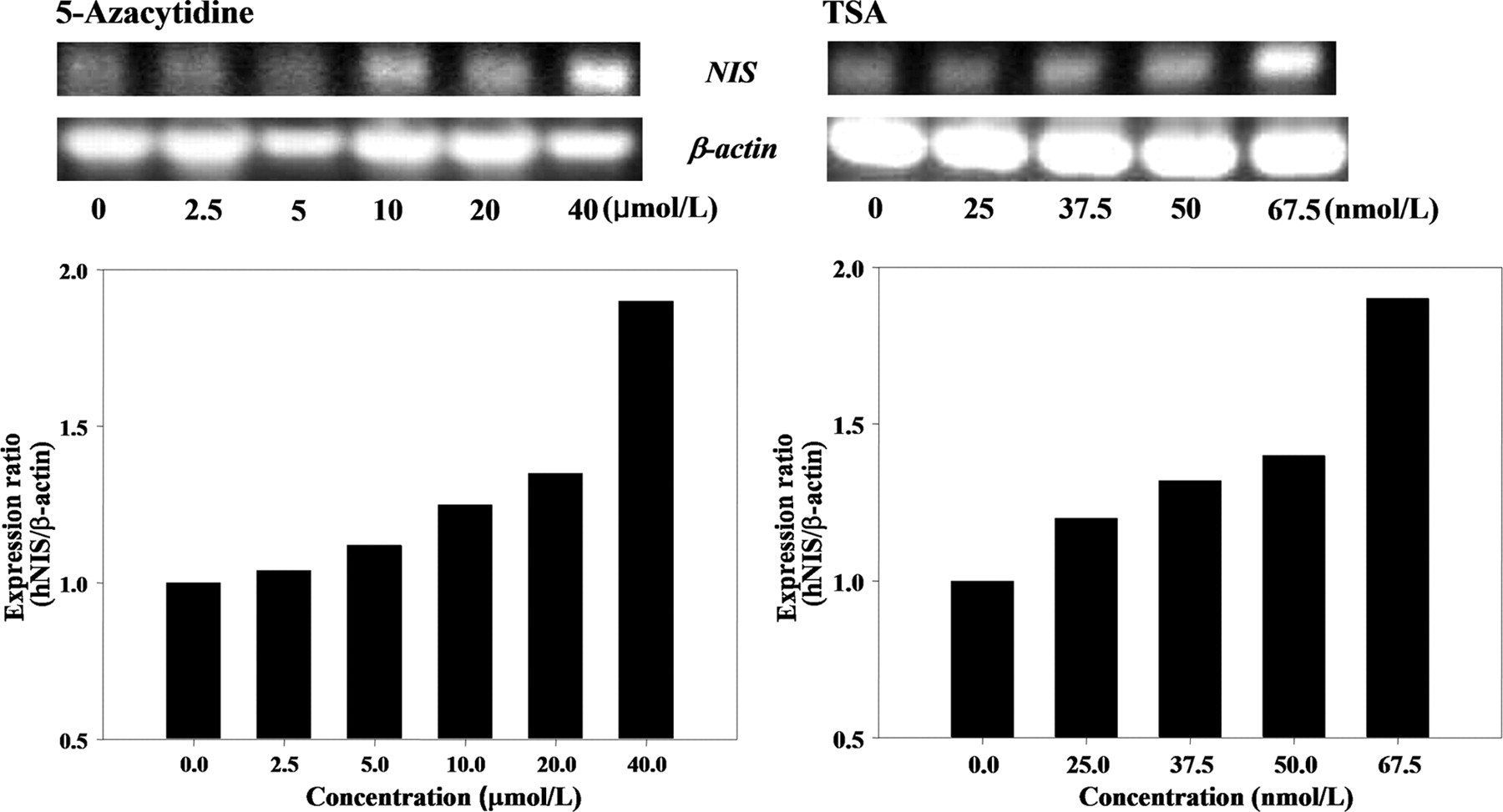
When F3-NIS III cells were treated with different concentrations of 5-azacytidine (2.5, 5, 10, 20, or 40 μmol/L), 125I uptake was dependent on the 5-azacytidine concentration. 125I uptake increased up to 1.8-fold (99.5 ± 6.5 pmol/mg protein) higher uptake with 40 μmol/L of 5-azacytidine than nontreated F3-NIS III cells (56.1 ± 2.7 pmol/mg protein) (Fig. 3A). When F3-NIS III cells were treated with different concentrations of TSA (25, 37.5, 50, or 62.5 nmol/L), 125I uptake increased, dependent on the TSA concentration. With 62.5 nmol/L of TSA, 125I uptake increased up to 36.2-fold (1,727.7 ± 85.8 pmol/mg protein) higher than that of nontreated F3-NIS III cells (47.8 ± 3.5 pmol/mg protein) (Fig. 3B). Moreover, the increased 125I uptake of F3-NIS III cells by 5-azacytidine or TSA treatment was completely inhibited by potassium perchlorate.
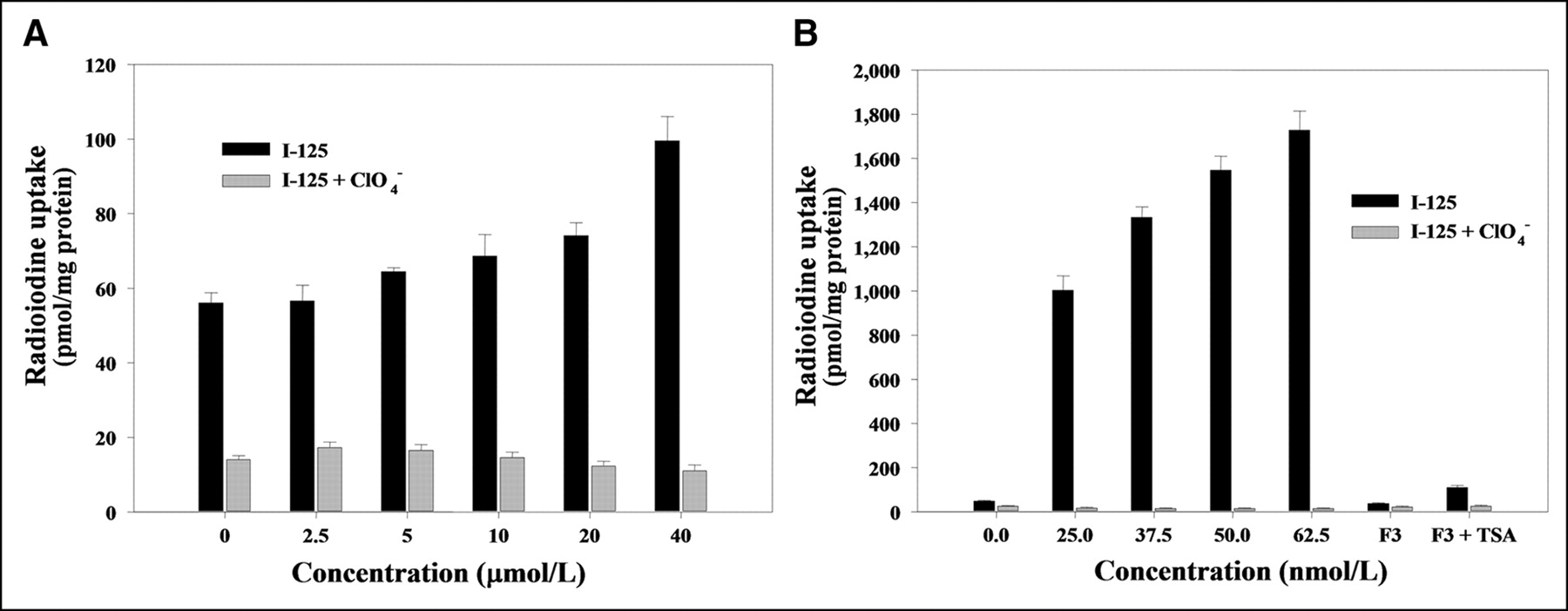
Increase of 125I uptake by DNA demethylation by 5-azacytidine (A) and histone deacetylase inhibition by TSA (B) in F3-NIS III cells. Increases of 125I uptake were completely inhibited by KClO4. Treatment with 62.5 nmol/L TSA in NIS nontransfected HB1.F3 cells also increased 125I uptake. Results are means ± SD (n = 4).
When hNIS nontransfected HB1.F3 cells were treated with 62.5 nmol/L of TSA, 125I uptake increased by 3-fold (110.8 ± 8.6 pmol/mg protein) compared with that in nontreated HB1.F3 cells (36.7 ± 3.9 pmol/mg protein). TSA itself increased endogenous hNIS expression in nontransfected hNIS cells but the increase was much less than that in hNIS transfected F3-NIS III cells (Fig. 3B).
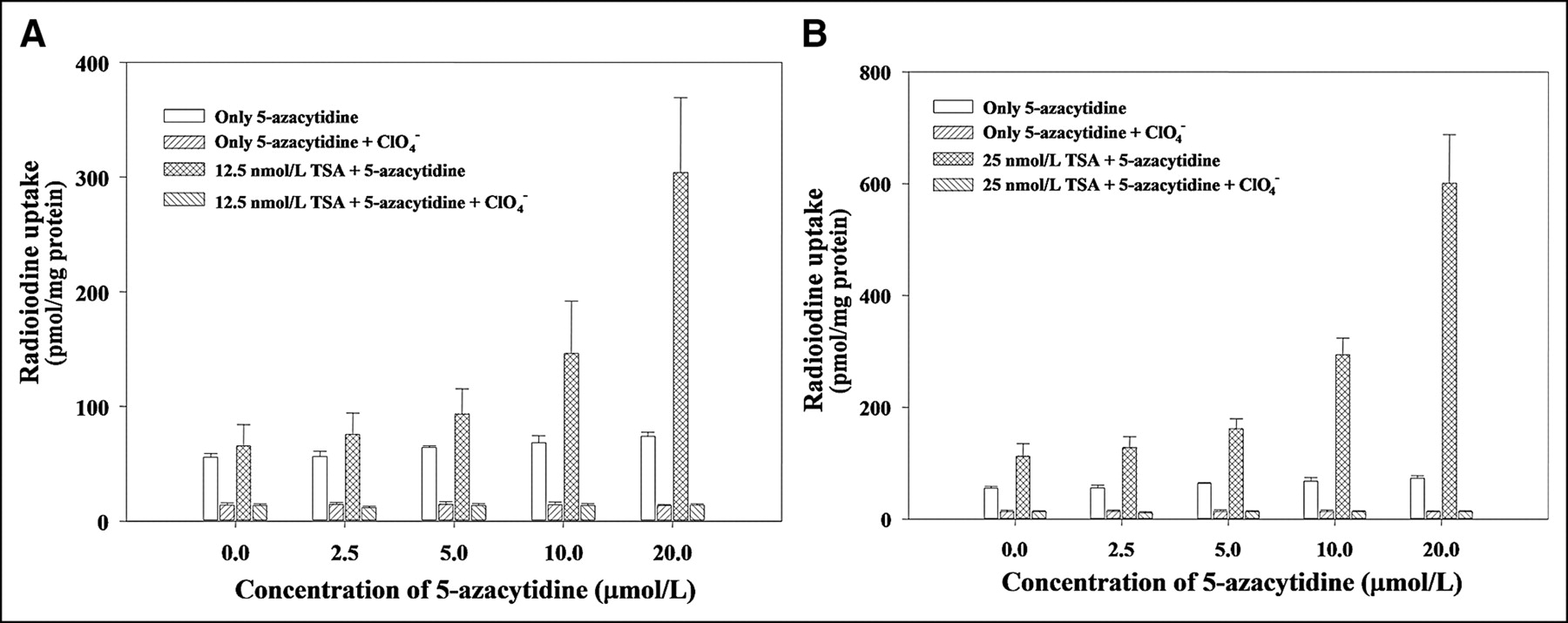
To find the synergistic effect of 5-azacytidine and TSA on hNIS gene expression in F3-NIS III cells, cells were treated with 12.5 or 25 nmol/L of TSA and different concentrations of 5-azacytidine (2.5, 5, 10, or 20 μmol/L) for 24 h. When F3-NIS III cells were treated with 12.5 nmol/L of TSA and different concentrations of 5-azacytidine (2.5, 5, 10, or 20 μmol/L) for 24 h, 125I uptake increased in an 5-azacytidine dose-dependent manner up to 4.6-fold (304.2 ± 14.1 pmol/mg protein) higher than that of 5-azacytidine nontreated F3-NIS III cells (65.8 ± 18.3 pmol/mg protein) (Fig. 4A). Also, F3-NIS III cells were treated with 25 nmol/L of TSA and different concentrations of 5-azacytidine (2.5, 5, 10, or 20 μmol/L) for 24 h. 125I uptake of F3-NIS III cells increased, in an 5-azacytidine dose-dependent manner, up to 5.3-fold (601.1 ± 86.5 pmol/mg protein) higher than that of 5-azacytidine nontreated F3-NIS III cells (113.3 ± 21.6 pmol/mg protein) (Fig. 4B). A synergistic effect was found with a high dose of 5-azacytidine and a low dose of TSA.

Synergistic effect of TSA (A, 12.5 nmol/L; B, 25 nmol/L) and various concentrations of 5-azacytidine (0, 2.5, 5, 10, or 20 μmol/L) in F3-NIS III cells. At a low concentration of 5-azacytidine (0–5 μmol/L), 125I uptake in F3-NIS III cells was not influenced by combined treatment. However, at a high concentration of 5-azacytidine (20 μmol/L), 125I uptake in F3-NIS III cells was higher than additive uptakes of 5-azacytidine and TSA. Results are means ± SD (n = 4).
Effect of Demethylation (5-Azacytidine) or HDAC Inhibition (TSA) on Messenger RNA (mRNA) Expression of hNIS Genes
When F3-NIS III cells were treated with 5-azacytidine, the mRNA level of the hNIS gene was 1.9-fold higher with 40 μmol/L of 5-azacytidine than that of nontreated F3-NIS III cells. When F3-NIS III cells were treated with TSA, the mRNA level of the hNIS gene was 1.9-fold higher with 62.5 nmol/L of TSA than that of nontreated F3-NIS III cells (Fig. 5).

Recovery of hNIS gene transcription after 5-azacytidine and TSA treatment. mRNA level of hNIS increased according to 5-azacytidine and TSA treatment.
Scintigraphic Imaging of hNIS Genes in F3-NIS III Cells Implanted into Nude Mice
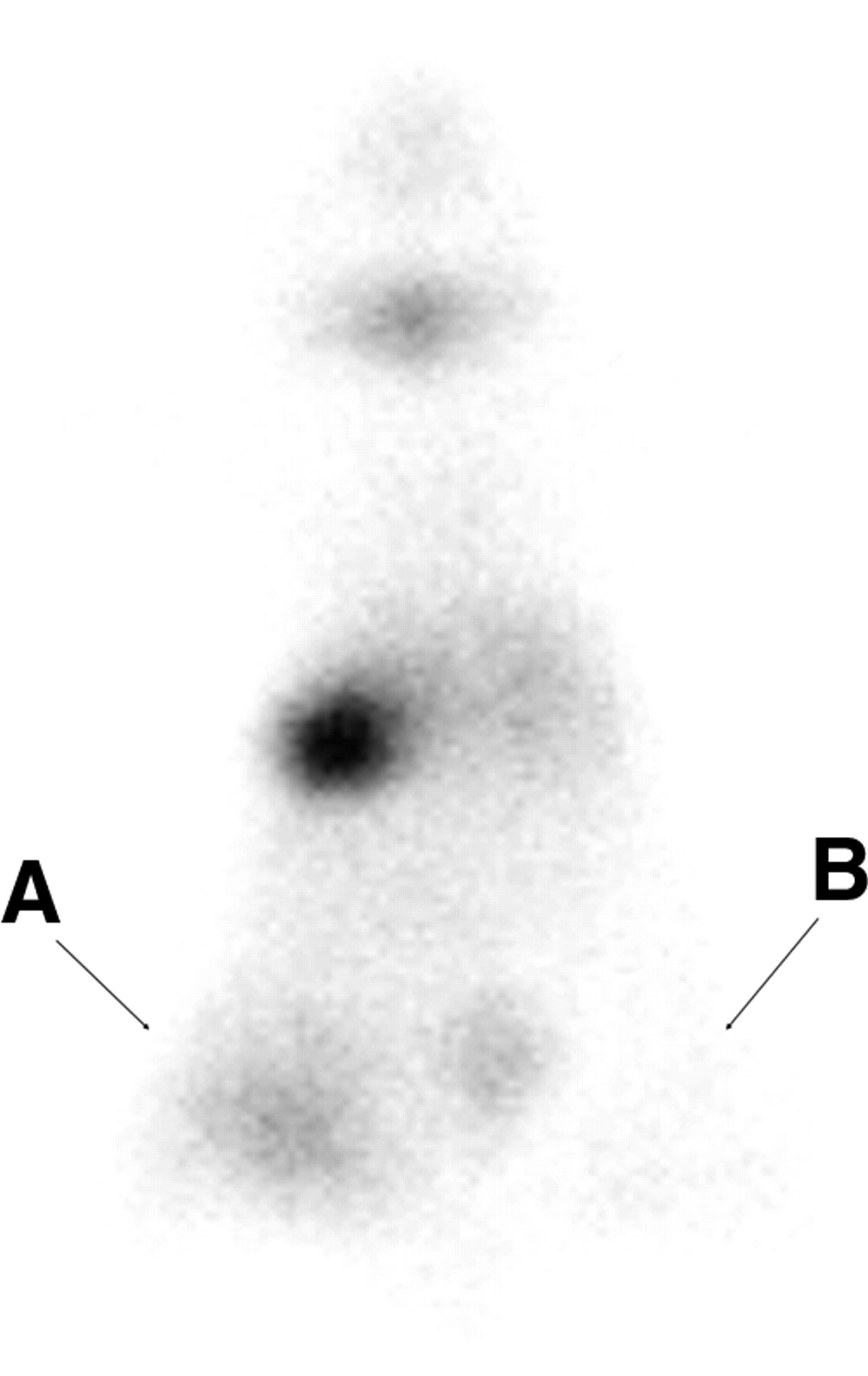
F3-NIS III cells were treated with 62.5 nmol/L of TSA in vitro for 24 h and then implanted into BALB/c nu/nu mice. Three hours later, the animals were injected intraperitoneally with 11.1 MBq (300 μCi) of 99mTc pertechnetate and images were taken 30 min later. The TSA-treated F3-NIS III cells were clearly visible, whereas nontreated F3-NIS III xenografts were not (Fig. 6).
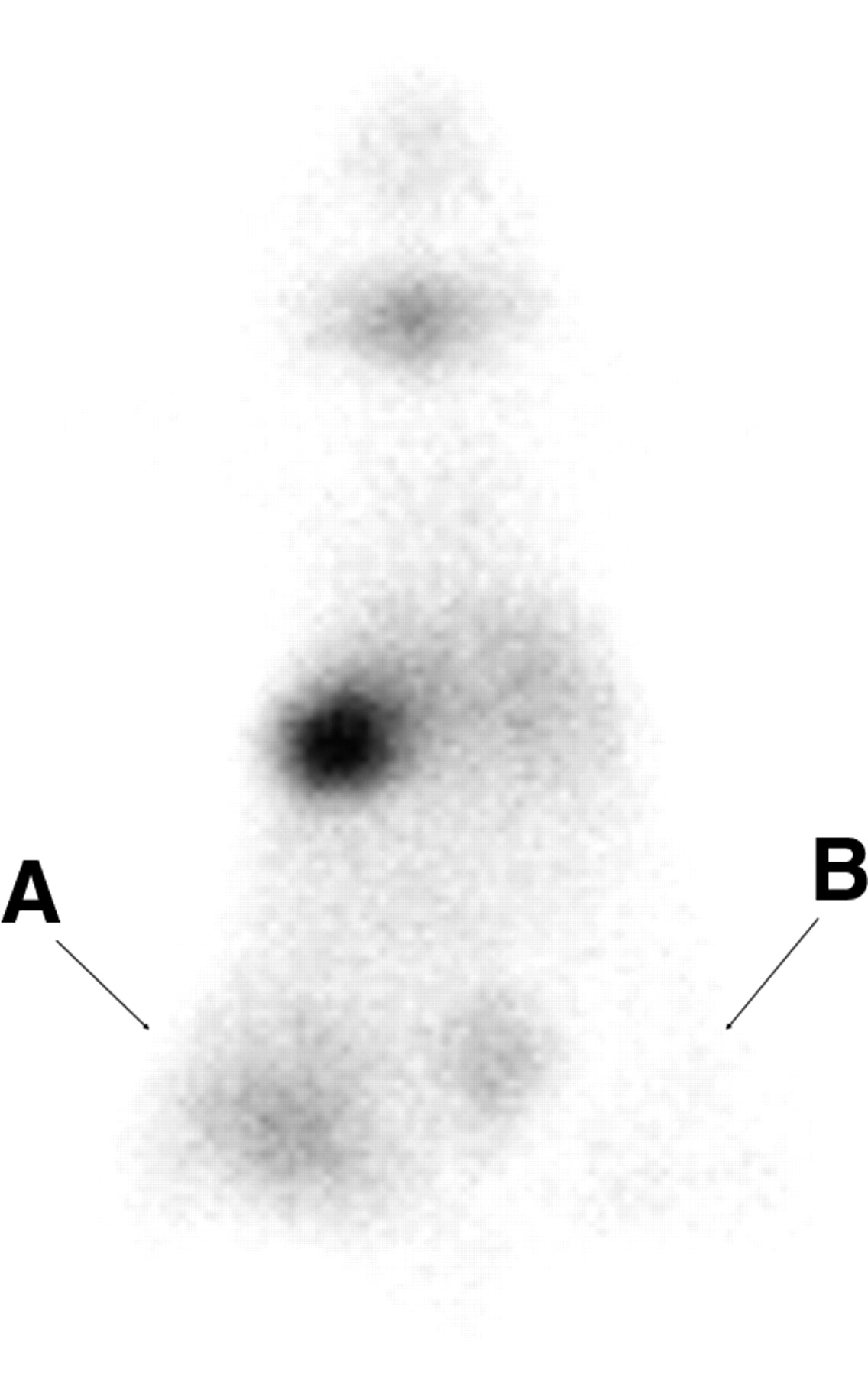
Scintigraphic image of hNIS gene in 62.5 nmol/L TSA-treated F3-NIS III cells implanted into a nude mouse. F3-NIS III cells were treated with 62.5 nmol/L of TSA in vitro for 24 h and then implanted into BALB/c nu/nu mice. Three hours later, animals were injected intraperitoneally with 11.1 MBq (300 μCi) of 99mTc-pertechnetate and images were taken 30 min later. TSA-treated F3-NIS III cells were clearly visible on left (A) compared with nontreated F3-NIS III cells (B).
DISCUSSION
Because reporter transgenes, but not MRI probes, can be transmitted to progeny cells, optical or nuclear imaging looks better than MRI for stem cell tracking (10–14). However, though transmissible to daughter cells, the expression of the transgenes is not perpetual; either the transgenes might be excluded or the expression of the transgenes is silenced. Stability of transgene expression in stem cells is not easily achieved, and this characteristic is one of the obstacles for stem cell tracking using optical (15) or nuclear imaging. After culture passage, the expression of green fluorescent protein (GFP) was silenced in neural precursor cell lines (15), and we now report that the expression of hNIS was also silenced after culture passage in HB1.F3 neural stem cell lines.
In this study, we produced stable HB1.F3 human neural stem cells transfected with hNIS and, at first, the 3 colonies stably expressed the hNIS gene. After several passages of in vitro culture of these clones, 1 colony gradually lost expression almost down to the pretransfection level. Initially, the expression level of hNIS in each colony differed, probably because of the positional effects caused by the influence of the site of chromosomal integration or further epigenetic modulation, such as DNA methylation and histone deacetylation.
To our knowledge, no way has been reported so far to prevent the silencing of transgenes in neural stem cells. If we are going to use our transfected cells for stem cell tracking, we have either to maintain the expression of the transgenes or, if silenced, to reactivate the transgenes. The mechanisms of transgene silencing were studied in various situations, such as allele-specific imprinting in embryonic tissues (16), imprinting of differentiation genes early in the mouse development (17), and tumor suppressor gene silencing in cancer (18). DNA methylation and histone deacetylation were the 2 major well-known mechanisms. In cancer tissues, drug-induced inhibition of DNA methylation and histone deacetylation had anticancer effects through the epigenetic modulation of tumor suppressor genes, and these 2 mechanisms showed synergism (18).
Upon separation of a methyl group from the methylated CpG sites of promoter, gene expression began (6,17,19,20). As the silencing of an endogenous gene is not completely irreversible by methylation of the promoter CpG sites, the reactivation of the gene can be induced by the demethylation of the promoter and by subsequent inhibition of histone deacetylase (18,21). Although there are the other mechanisms for gene silencing (22), DNA methylation and histone deacetylation are mainly involved in transgene silencing (15,23).
In this study, we demonstrated that these 2 well-known mechanisms of epigenetic modulation were involved in silencing the reporter hNIS transgene in the F3-NIS III cells. The 125I uptake, which means the functional integrity of the hNIS transgene protein in F3-NIS III cells, decreased according to the cell passage. Although exogenous hNIS still existed in F3-NIS III cells even after the eighth passage, the amount of mRNA decreased. When we treated F3-NIS III cells with 5-azacytidine or TSA, the expression of hNIS in F3-NIS III cells recovered to more than the initial level. This evidence indicated that the reduced expression of hNIS in F3-NIS III cells was due to epigenetic modulation. In terms of the quantity in F3-NIS III cells, inhibition of histone deacetylation contributed more to reactivate hNIS transgenes than DNA demethylation. Interestingly, the expression of endogenous hNIS of F3 cells was also increased, which implied that endogenous hNIS was also regulated by the same mechanism of epigenetic modulation. Whether this modulation is confined to these neural stem cells or may apply to the other cells in general warrants further study.
We also observed the synergistic effects of combined treatment with high-dose 5-azacytidine (10 or 20 μmol/L) and TSA (12.5 or 25 nmol/L) for reactivating transgenes just like in U937 transduced cells (23). We did not investigate the reversing capability of the demethylating agent (5-azacytidine) in the other passage lines, either earlier or later than the eighth passage.
The transcription level of hNIS mRNA increased in a 5-azacytidine–dependent manner, but the function of hNIS was increased, not in its full capacity. We speculate that the other contributing genes are still not active for the function of the symporter activity that is not fully recovered by DNA demethylation. On the other hand, the transcription level of hNIS mRNA increased in a small amount after TSA treatment, but the hNIS function was much increased. The efficiency of posttranscriptional translation or posttranslational modulation might have also been affected by HDAC inhibition. The synergism of 5-azacytidine and TSA implies that this posttranscriptional or posttranslational effect was not restricted to TSA.
DNA demethylation and inhibition of histone deacetylation are involved in cell proliferation and differentiation (24,25). These 2 mechanisms can induce a variety of effects that are dependent on the genes expressed, their balances, and the cell lineage. After modulation, they might differentiate or proliferate and escape from the resting stage to the proliferative stage in the cell cycle. Neuroprogenitor cells matured to neurons after demethylation (26), but the cortical progenitor cells differentiated to oligodendrocytes by histone deacetylation (27). Promoter demethylation of the GFAP (glial fibrillary acidic protein) gene in normal brain tissue and in neoplastic glial cells lets the progenitor cells in brain differentiate to neurons (28). Using these characterstics, we might be able to investigate simultaneously the expression of reporter transgenes and the cellular differentiation in neural stem cells.
Rosenqvist et al. (15) reported similar modulation of GFP transgenes in neural stem cells. The increase of HDAC by TSA inhibition was 3-fold. In quantitative terms, the increases of hNIS activity (125I uptake) were more prominent and favored imaging with a much better signal-to-noise ratio. When we implanted a billion cells in the subcutaneous tissue of nude mice, we could clearly demonstrate that F3-NIS III cells took up the radioactive probe—that is, 99mTc-pertechnetate. TSA-rescued F3-NIS III cells took up 99mTc-pertechnetate but hNIS-silenced F3-NIS III cells did not. These positive data demonstrated that the TSA treatment effect persists in vivo after implantation. However, it is unknown whether pharmacologic modulation of hNIS expression in already transplanted neural stem cells is possible. Further studies are necessary to determine how long the reactivation would last.
The distribution and homing of transplanted neural stem cells are not yet fully disclosed. hNIS transfected human neural stem cells were found to be effective in inducing the functional recovery of a neurologic deficit in small animals (9,29). However, because we do not know the exact fate of transplanted stem cells, we suggest that hNIS transfected neural stem cells should be monitored for their fate after transplantation. Molecular imaging of transplanted stem cells in small animal models of neurologic diseases will elucidate still-unknown mechanisms of the efficacy of stem cell-based therapy of human diseases. The detection sensitivity of NIS activity as a reporter probe in vivo and the feasibility of using NIS transfected human neural stem cells should be examined carefully before considering clinical imaging application.
CONCLUSION
For tracking neural stem cells transfected with hNIS reporter gene, reversing the silencing and thus perpetuation of transgene expression was tried in neural stem cells. In this study, we demonstrated that 2 well-known mechanisms of epigenetic modulation are involved in the silencing of the reporter hNIS transgene in a human neural stem cell line. This reversing of hNIS transgene silencing was found to be effective in vivo as well as in vitro by the DNA demethylation agent (5-azacytidine) and the histone deacetylase inhibitor (TSA).
Acknowledgments
This study was supported by the BK21 Human Life Science Program.
Footnotes
Received June 1, 2004; revision accepted Aug. 12, 2004.
For correspondence or reprints contact: Dong Soo Lee, MD, Program in Neuroscience, Seoul National University College of Natural Science, Department of Nuclear Medicine, Seoul National University College of Medicine, 28 Yeongeon-Dong Jongno-gu, Seoul, 110-744, Korea.
E-mail: dsl{at}plaza.snu.ac.kr
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Visualization of Hypoxia-Inducible Factor-1 Transcriptional Activation in C6 Glioma Using Luciferase and Sodium Iodide Symporter Genes
- Imaging of Mesenchymal Stem Cell Transplant by Bioluminescence and PET
- Imaging Stem Cells Implanted in Infarcted Myocardium
- Stem Cell Tracking: Reversing the Silence of the Lambs...