


Abstract
α-Particle–emitting radionuclides are of increasing interest in radionuclide therapy. The decay scheme of α-emitting radionuclides typically includes a chain of unstable progeny. It is generally assumed that α-particle emission by the parent radionuclide will break the chemical bond with its carrier molecule and that the resulting daughter atom will no longer be associated with the carrier molecule. If the daughter is very short lived, it will not have enough time to be carried any significant distance from the site of parent decay and a cellular, absorbed dose estimate must consider the energy deposited by the daughter as well as the parent. Depending on the site of parent decay and the expected removal rate of daughter atoms from this site, the contribution of emissions from longer-lived daughters may also be warranted. In this study, dose conversion factors (DCFs) for cellular dimensions that incorporate the fate of daughter radionuclides were derived for 225Ac, 213Bi, 211At, and 223Ra, the α-particle–emitting radionuclides of interest in radionuclide therapy. Methods: The dose contribution of daughter radionuclides at the site of parent decay was made dependent on a cutoff time parameter, which was used to estimate the fraction of daughter decays expected at the site of parent decay. Previously tabulated S values (cell-surface to nucleus and cell-surface to cell) for each daughter in the decay scheme were scaled by this fraction and a sum over all daughters was performed to yield a cutoff time–dependent set of corresponding DCF values for each radionuclide. Results: DCF values for the absorbed dose to the nuclear or cellular volume from cell-surface decays are presented as a function of the cutoff time for 4 different cellular and nuclear dimensions. Conclusion: In contrast to the cellular S values that account only for parent decay, the DCF values provided in this study make it possible to easily include the contribution of daughter decays in cellular α-particle emitter dose calculations.
Radionuclides that emit α-particles are of increasing interest in radionuclide therapy (1,2). Because the decay chain of several important α-particle emitters includes daughters that decay, which releases more α-particles, the cellular S value for the parent alone may not properly reflect the true α-particle energy deposition to a particular target per decay of the parent. The actual energy deposited will depend on the degree to which daughter decays contribute dose to the site of parent decay. This, in turn, will be influenced by the half-life of the daughter products and the rate at which they are removed from the site of parent decay. The latter will be related to the milieu within which the parent has decayed. If the half-life is very short compared with the removal or diffusion rate of the radionuclide, then the dose contribution from daughter decays should be included in cellular dose calculations.
In this study, we present a set of dose conversion factors (DCFs) that are analogous to cellular S values but extend this concept by including the cutoff time–dependent dose contribution of daughter decays at the site of parent decay. The cutoff time is thus introduced as a parameter that represents assumptions regarding the degree to which daughters should be included.
MATERIALS AND Methods
Cellular S values for 4 α-particle emitters (225Ac, 211At, 213Bi, and 223Ra) and their progeny were obtained from the MIRD tabulation of cellular S values (3). S values for 213Po, one of the elements in the decay chain of 213Bi, are not tabulated and therefore needed to be calculated by spline interpolation of the monoenergetic α-particle S value tables. Similarly, S values for 207Bi, one of the elements in the 211At decay chain, which decays with electron capture or—albeit with very small probability—through positron decay, are not tabulated. These were calculated by spline interpolation as well. The contribution of daughter radionuclides to the absorbed dose at the site of parent decay was obtained by calculating the fraction of daughter decays, f, expected within a cutoff time parameter, τ0.
The total number of daughter decays for a given initial radioactivity, A0, is given by:
 Eq. 1
where λ = [ln(2)/T1/2] and T1/2 = the half-life of the radionuclide.
Eq. 1
where λ = [ln(2)/T1/2] and T1/2 = the half-life of the radionuclide.
The number of decays occurring within τo is:
 Eq. 2
Eq. 2
The ratio of Equations 1 and 2 gives the fraction of total decays, g, occurring within a cutoff time, τo:
 Eq. 3
where τ corresponds to the average lifetime of the radionuclide (=1/λ).
Eq. 3
where τ corresponds to the average lifetime of the radionuclide (=1/λ).
Because the contributing fraction of daughter decays is dependent on the amount of daughter decays and the branching ratio (the percentage that decays into a specific daughter) of the radionuclide immediately preceding it, the actual fraction is the product of the calculated fraction, gi, for daughter i, of the preceding radionuclide’s calculated fraction, g(i−1), and of the branching ratio for production of daughter i, ri:
 Eq. 4
Eq. 4
These fractions, fi, are then multiplied by the cellular S values, Si, for each daughter i and summed to yield the final DCF,
 Eq. 5
Eq. 5
Equation 5 was used to generate graphs of DCFs as a function of τ0 for 225Ac, 213Bi, 211At, and 223Ra for 4 different cellular dimensions.
Results
Table 1 lists cellular S values for 213Po, and Table 2 lists cellular S values for 207Bi for various source–target combinations and increasing radii for cell (RC) and nucleus (RN). The possible source locations are cell, cell surface, cytoplasm, and nucleus. The targets are either cell (C) or nucleus (N). For example, S(N ← Cy) refers to the S value when the nucleus is the target and the activity is uniformly distributed in the cytoplasm (3). Figure 1 depicts the decay schemes of the 4 radionuclides studied (4). Figures 2 and 3 show the DCF plots for cell-surface to cell and cell-surface to nucleus for those radionuclides. With the exception of 213Bi, DCF values at the shortest cutoff time (0.001 min) are comparable with the original cellular S value for a parent radionuclide. At the longest cut-off times (1,000 min), the DCF values reflect the sum of parent and all daughter S values.
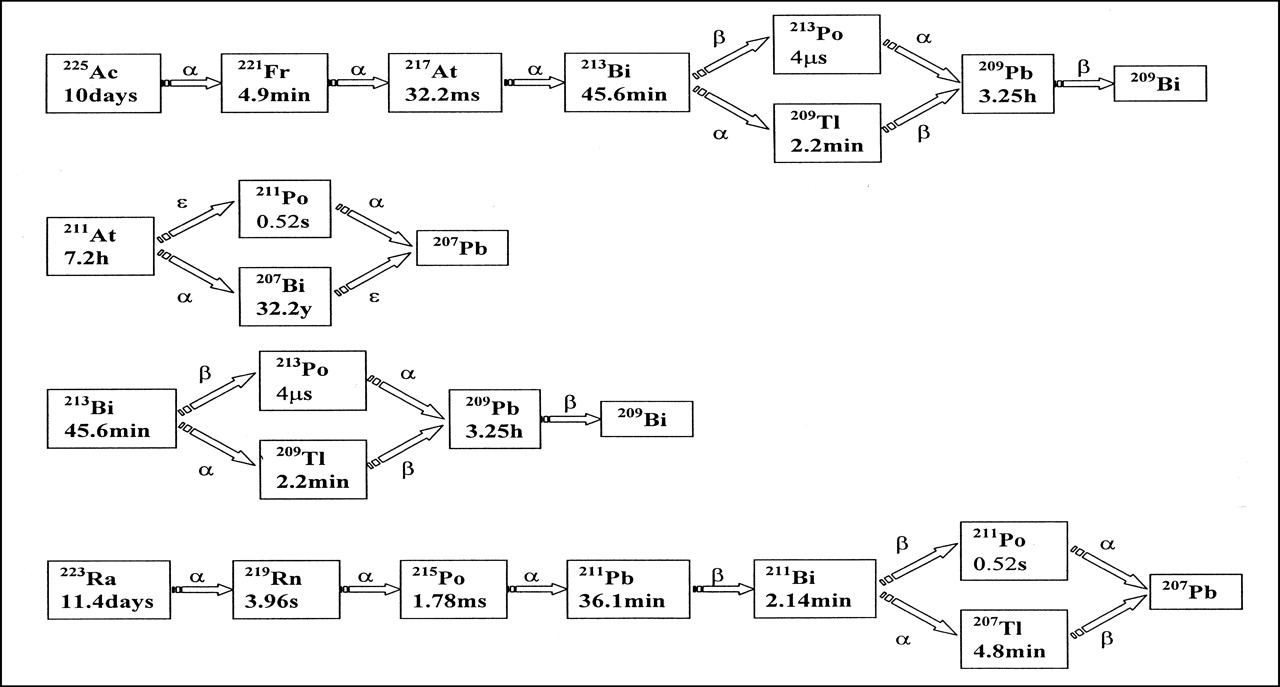
Decay schemes for 225Ac, 211At, 213Bi, and 223Ra.

DCF values vs. τ0 for (C ← CS) where CS represents cell surface. Plots for several different cellular dimensions are shown and denoted by (RC, RN), representing radius of cell and of nucleus, respectively.

DCF values vs. τ0 for (N ← CS) where CS represents cell surface. Plots for several different cellular dimensions are shown and denoted by (RC, RN), representing radius of cell and of nucleus, respectively.
Cellular S Values for 213Po
Cellular S Values for 207Bi
Points of transition to higher DCF values reflect the inclusion of additional daughters as contributing to the absorbed dose at the site of parent decay. In the 223Ra graph (Fig. 2), a plateau is present from 0 min to approximately 0.01 min because only the parent contributes. From τ0 = 0.01 min through τ0 = 0.3 min, an increase in the DCF can be seen that is caused by the progressive retention of 219Rn (τ = 0.1 min) and 215Po (τ = 4 × 10−5 min) at the site of parent decay. Because 215Po has an average lifetime shorter than that of 219Rn, the fraction of 215Po that is removed from the 219Rn decay site is smaller than the fraction of 219Rn that is removed from the 223Ra decay site. Thus, both will always contribute together to the DCF. The plateau present from τ0 = 0.3 min through τ0 = 10 min represents cutoff times at which the emissions from subsequent daughters may be ignored. The next increase, from τ0 = 10 min through τ0 = 100 min, represents the inclusion of 211Pb (τ = 52 min) and all subsequent daughter radionuclides at the site of parent decay. The last plateau, from 100 min to 1,000 min, corresponds to the maximum DCF value in which all daughters are included. In the plot for 213Bi (Fig. 2), the S value contribution of the β-emitter, 209Pb (τ = 281 min), although small relative to α-particle S values, may be seen as a slight increase between 100 min and 1,000 min.
Discussion
The decay scheme of α-particle–emitting radionuclides typically includes a chain of unstable progeny. It is generally assumed that α-particle emission by the parent radionuclide will break the chemical bond with its carrier molecule and that the resulting daughter atom will no longer be associated with the carrier molecule. If the daughter is very short lived (e.g., 213Po with a 4.2-μs half-life), then it will not have enough time to move any significant distance from the site of parent decay and an absorbed dose estimate must consider energy deposited by the daughter as well as by the parent.
Drawing on recently published cellular S values and incorporating the considerations described above, we have derived DCFs for α-particle–emitting radionuclides that either have already been used clinically, such as 213Bi and 211At, (5,6) or are considered to be promising candidates for clinical use, such as 225Ac and 223Ra (7,8).
Using Figures 2 and 3, biologic and kinetic considerations regarding the site of parent decay and possible fate of daughters may easily be incorporated into cellular absorbed dose calculations. Because blood convection will lead to a more rapid loss of daughter radionuclides from the site of parent decay than would tumor interstitium diffusion, the likelihood that daughters generated within the circulation will contribute significantly to the dose at the site of parent decay is considerably less than if the daughters were generated within the interstitium. By selecting appropriate cutoff times in Figures 2 and 3, different levels of daughter contribution may be chosen to match the milieu in which the parent radionuclide decays. In general, DCFs for parent decays in circulation are best approximated by the leftmost portions of the plots, where only the parent is included. In the tumor interstitium, longer cutoff times may be appropriate and the middle portions of the plots would apply. The rightmost portions of the plots should be used to obtain DCF values in situations in which all daughters are expected to remain at the site of parent decay.
Conclusion
The extent to which daughter products remain at the site of parent decay will greatly influence the potential efficacy, toxicity, and clinical utility of α-particle–emitting radionuclides with decay schemes that include α-particle–emitting daughters. The figures presented in this article facilitate the use of different assumptions regarding daughter radionuclide retention at the site of parent decay.
Acknowledgments
The authors thank Joseph O’Donoghue for helpful discussions. This study was supported by National Institutes of Health grants R01 CA62444, R01 CA72683, and P01 CA-33049 and the Scallon Foundation.
Footnotes
Received Aug. 2, 2000; revision accepted Nov. 7, 2000.
For correspondence or reprints contact: George Sgouros, PhD, Department of Medical Physics, Memorial Sloan-Kettering Cancer Center, 1275 York Ave., New York, NY 10021.
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Targeting Aberrant DNA Double-Strand Break Repair in Triple-Negative Breast Cancer with Alpha-Particle Emitter Radiolabeled Anti-EGFR Antibody
- MIRD Pamphlet No. 22 (Abridged): Radiobiology and Dosimetry of {alpha}-Particle Emitters for Targeted Radionuclide Therapy
- Radioimmunotherapy of Breast Cancer Metastases with {alpha}-Particle Emitter 225Ac: Comparing Efficacy with 213Bi and 90Y
- Microdosimetric Analysis of {alpha}-Particle-Emitting Targeted Radiotherapeutics Using Histological Images
- {alpha}-Particle Emitters in Radioimmunotherapy: New and Welcome Challenges to Medical Internal Dosimetry