


Abstract
In most types of peptide receptor radionuclide therapy, the maximum activity dose that can be administered is limited by high and persistent renal retention of the radiolabeled peptides, which is, at least partly, mediated by the megalin receptor. Several agents that interfere with renal reabsorption of radiolabeled peptides have been identified (e.g., lysine, arginine, succinylated gelatin solution), but none of these inhibit renal reabsorption completely. Albumin, a naturally abundant megalin ligand, might be a safe and potent alternative. In this study, we analyzed the effects of albumin and fragments of albumin (FRALB) on the renal reabsorption of 111In-diethylenetriaminepentaacetic acid (DTPA)-d-Phe1-octreotide (111In-octreotide), [Lys40(aminohexoic acid-DTPA-111In)NH2]-exendin-4 (111In-exendin), and 111In-1,4,7,10-tetraazacyclododecane-N,N′,N″,N′″-tetraacetic acid (DOTA)-Glu1-minigastrin (111In-minigastrin). Methods: The effects of albumin and FRALB on megalin-associated binding of 111In-octreotide, 111In-exendin, and 111In-minigastrin were assessed in vitro using rat yolk sac epithelial (BN16) cells. In vivo, uptake and localization of 111In-albumin and 111In-FRALB in the kidneys of Wistar rats were determined, as well as the effect of lysine, succinylated gelatin solution, albumin, and FRALB on the kidney uptake of 111In-octreotide, 111In-exendin, and 111In-minigastrin. Results: FRALB significantly reduced binding and uptake of 111In-octreotide, 111In-exendin, and 111In-minigastrin by BN16 cells. In rats, renal uptake of 111In-labeled FRALB was significantly higher than that of 111In-labeled intact albumin (P < 0.001). FRALB administration effectively reduced renal uptake of 111In-octreotide, 111In-exendin, and 111In-minigastrin. Administration of 1–2 mg of FRALB reduced renal uptake of 111In-octreotide as efficiently as 80 mg of lysine. Conclusion: Renal uptake of 111In-octreotide and other radiolabeled peptides in rats can be effectively reduced by administration of albumin fragments. Additional studies to identify the albumin fragments responsible for inhibition of renal peptide uptake are warranted.
Elimination of radiolabeled hydrophilic peptides from the body occurs preferably via the kidneys. For scintigraphic imaging, rapid excretion of these compounds is preferable to obtain high-quality images with low background activity (1). In peptide receptor radionuclide therapy, rapid clearance of the radiolabeled peptides from the blood and low retention in the kidneys are important to minimize the radiation dose to normal tissues, especially the kidneys. However, many of these small proteins and peptides are partly retained in the kidneys because of tubular reabsorption. Renal reabsorption of (radiolabeled) peptides and proteins is a multistep process (2,3). It is thought that after glomerular filtration, proteins and peptides in the ultrafiltrate bind to endocytic receptors at the luminal surface of proximal tubular cells and are internalized. Subsequently, the compounds are transferred to the lysosomes and degraded into amino acids (2,3). These amino acids are transported back into the bloodstream. However, radiometal-chelated amino acids are trapped in the tubular cell lysosomes, thereby delivering high radiation doses to the kidneys (1,4).
The exact mechanism of tubular reabsorption of peptides is not completely understood, but several studies have shown a role for megalin (5,6). Megalin is a multiligand receptor belonging to the low-density-lipoprotein receptor family. The receptor binds various structurally different proteins (albumin, vitamin D binding protein, β-2 microglobulin, aprotinin, and many others) (5). It has been shown that some ligands bind to megalin via their cationic sites and that coadministration of basic compounds can inhibit renal reabsorption of these ligands (6,7). Indeed, it has been shown in patients undergoing peptide receptor radionuclide therapy with radiolabeled octreotide, as well as in various animal models, that renal uptake of different radiolabeled peptides and antibody fragments can effectively be reduced by coadministration of basic amino acids such as lysine and arginine (1,4,8,9). However, these compounds can cause side effects at high doses, such as nausea, hyperkalemia, arrhythmias, and even nephrotoxicity (7,9,10).
Because megalin seems to play an important role in tubular reabsorption of radiolabeled peptides, we hypothesized that the megalin substrate albumin could also reduce renal uptake of radiolabeled peptides (11,12). Albumin is a protein with a molecular weight of 67 kDa (7). Only a very small fraction of total albumin passes the glomerular membrane and is reabsorbed by the renal tubular cells (6,13–15).
Because only a small fraction of total intact albumin passes the glomerular membrane, we further hypothesized that fragments of albumin might be more suitable for inhibiting tubular reabsorption of peptides.
In this study, the effect of fragments of albumin (FRALB) on the renal uptake of [111In-diethylenetriaminepentaacetic acid (DTPA)-d-Phe1]-octreotide (111In-octreotide), [Lys40(aminohexoic acid (Ahx)-DTPA-111In)NH2]-exendin-4 (111In-exendin), and [111In-1,4,7,10-tetraazacyclododecane-N,N′,N″,N‴-tetraacetic acid (DOTA)-Glu1]-minigastrin (111In-minigastrin) was analyzed, both in vitro and in rats.
MATERIALS AND METHODS
Albumin Fragments
Intact albumin was fragmented by trypsin digestion. Trypsinization was performed in 50 mM ammonium carbonate buffer, pH 7.8, by adding 500 mg of trypsin (Sigma) to 4.5 g of bovine serum albumin (Sigma) in 15 mL of buffer. The mixture was incubated at 37°C for 24 h. The trypsinized albumin (FRALB) was analyzed by fast protein liquid chromatography (FPLC) using a Superdex 75 size exclusion column (Pharmacia). The solution of FRALB was purified using 15-mL centrifugal filter units (Centriprep; Millipore), as outlined in Figure 1. After filtration of FRALB with a Centriprep YM-50 (50-kDa cutoff), the residue was discarded. The filtrate, containing albumin fragments with molecular weights of less than 50 kDa (FRALB < 50), was further fractionated using a Centriprep YM-3 filter (3.0-kDa cutoff), producing 2 additional solutions: the filtrate containing fragments with a molecular weight of less than 3 kDa (FRALB < 3) and the residue containing fragments with molecular weights ranging from 3 to 50 kDa (FRALB-3–50). The protein concentration in each fraction was estimated on the basis of light absorbance at 280 nm, as determined spectrophotometrically. The fractions were analyzed by FPLC as described above.
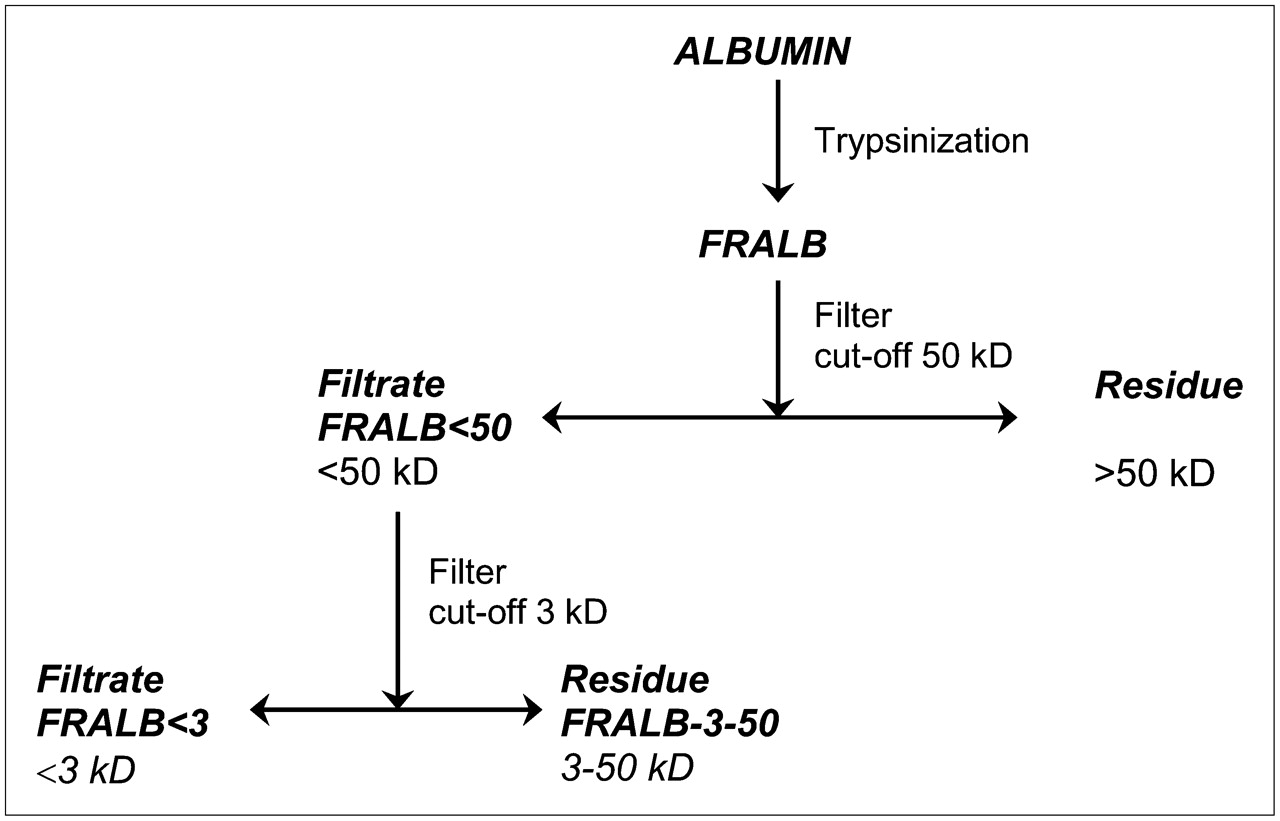
Production scheme of FRALB. Albumin was trypsinized, and resulting FRALB solution was purified via centrifugal filtration tubes to obtain solutions containing fragments with molecular weights of FRALB < 50, FRALB-3–50, and FRALB < 3.
DTPA Conjugation of Albumin and Albumin Fragments
Albumin and FRALB-3–50 were conjugated with DTPA by incubation with a 10-fold molar excess of cDTPA (Sigma) in 0.1 M carbonate buffer, pH 9.0, as described by Hnatowich et al. (16).
Radiolabeling
[DTPA-d-Phe1]-octreotide (Octreoscan; Tyco) was labeled with 111In by adding 37 MBq of 111InCl3 to 1 μg of Octreoscan and incubating for 30 min at room temperature.
[Lys40(Ahx-DTPA)NH2]-exendin-4 (exendin) was synthesized at Peptide Specialty Laboratories GmbH using standard Fmoc chemistry. The peptide was labeled with 111InCl3 in a 0.25 M ammonium acetate buffer, pH 5.5. A mixture of 3.7 MBq of 111In and 1 μg of exendin was incubated for 30 min at room temperature. Subsequently, ethylenediaminetetraacetic acid was added (12 μL, 0.05 M) and incubated for 10 min, after which polysorbate 80 (60 μL, 0.1%; Sigma) was added.
[DOTA-Glu1]-minigastrin (minigastrin) was synthesized at Peptide Specialty Laboratories GmbH using standard Fmoc chemistry and labeled with 111InCl3 in a 0.25 M ammonium acetate buffer, pH 5.5, containing 0.6 g of gentisic acid per liter. 5.5 MBq of 111In were added to 0.5 μg (5 μL) of minigastrin. The mixture was incubated at 95°C for 25 min.
Albumin-DTPA and FRALB-3–50-DTPA were labeled with 111InCl3 in a 0.25 M ammonium acetate buffer, pH 5.5. In a total volume of 500 μL, 10 MBq of 111InCl3 were added to 200 μg of albumin-DTPA or FRALB-3–50-DTPA and incubated for 30 min at room temperature.
Radiochemical purity and labeling efficiency of all compounds were determined by silica gel instant thin-layer chromatography and reverse-phase high-performance liquid chromatography. Radiochemical purity of all compounds was equal to or higher than 95%.
Compounds Used to Reduce Renal Reabsorption
Gelofusine (Braun) is a succinylated gelatin solution (40 g/L) (17,18). Lysine solution (lysine) was prepared at a concentration of 160 g/L in phosphate-buffered saline (PBS). Albumin and fragmented albumin (FRALB < 50, FRALB-3–50, and FRALB < 3) were prepared as described above. The molar mass of Gelofusine averages 3 × 104 g/mol (manufacturer's data); the molar mass of FRALB < 50 was estimated to average 1 × 104 g/mol.
In Vitro Studies
Rat yolk sac epithelial (BN16) cells expressing megalin (19) were kindly provided by Pierre J. Verroust. Cells were cultured at 37°C in BN16 medium consisting of Dulbecco's modified Eagle's medium plus 10 mM glucose (Invitrogen) with 10% fetal calf serum, 100 units of penicillin per milliliter, and 100 μg of streptomycin (Invitrogen) per milliliter.
The potency of FRALB to inhibit the interaction of the radiolabeled peptides with BN16 cells was compared with the potency of Gelofusine and lysine. The cells were seeded into 6-well plates at 3 × 105 cells per well and cultured until confluent. The cells were washed with PBS and incubated for 1 h with serum-free BN16 medium, after which they were washed twice with PBS, and 2 mL of Ringer's solution (pH 7.4) was added. Then the inhibitor was added, either lysine, Gelofusine, or FRALB < 50 at a 100,000-fold molar excess over the radiolabeled peptide. The concentrations of the inhibitors in the medium were 7 × 104 g of lysine per liter, 1 × 101 g of Gelofusine per liter, and 3 × 102 g of FRALB < 50 per liter. Directly after addition of the inhibitor, 1 kBq of 111In-octreotide, 111In-exendin, or 111In-minigastrin was added. After 1 h of incubation at 37°C, cells were rinsed once with Ringer's solution and harvested using cotton swabs. Radioactivity was measured in a shielded well-type γ-counter (Wizard; Wallac).
Animal Studies
Groups of 4 male Wistar rats (200–220 g) were used in all experiments. To investigate whether FRALB reached the proximal tubule of the kidney more efficiently than albumin, 111In-albumin and 111In-FRALB were administered intravenously to rats.
To compare the potency of FRALB to inhibit renal reabsorption of radiolabeled peptides with the potency of known inhibitors, animals were injected intravenously with 0.5 mL of the inhibitor (either PBS, lysine [80 mg], Gelofusine [20 mg], or FRALB < 50 [5 mg]) via the tail vein. After 2–5 min, 111In-octreotide, 111In-exendin, or 111In-minigastrin was injected intravenously (2 MBq/rat). For 111In-octreotide, a dose-response study of FRALB < 50 was performed, and the potencies of the filtered FRALB preparations (FRALB < 50, FRALB-3–50, and FRALB < 3) were compared.
Animals were killed 20 h after injection, a blood sample was drawn, and organs were dissected. Tissues were weighed and counted in a shielded well-type γ-counter (Wizard) together with the injection standards. Radioactivity concentration was expressed as percentage of the injected dose per gram of tissue (%ID/g). For autoradiography, a central 5-μm section was cut from each kidney and mounted on poly-l-lysine–coated slides. Kidney sections were exposed to a phosphor screen overnight, which was subsequently scanned in an imaging system (Molecular Imager GS363; BioRad Laboratories) at a pixel size of 100 × 100 μm.
Statistical Analysis
Data are presented as mean values ± SD. Analysis of cell binding and kidney uptake was performed using the Student t test. Biodistribution data in other organs were analyzed by 1-way ANOVA, with a correction for multiple datasets using Bonferroni's multiple comparison test. The significance level was set at 0.05.
RESULTS
Analysis of Albumin Fractions
The FPLC elution profiles of albumin and of the 3 FRALB preparations are shown in Figure 2. Approximately 50% of intact albumin was still present in the unpurified FRALB preparation (Fig. 2B). After ultrafiltration, FPLC analysis demonstrated that FRALB < 50 contained less than 30% of intact albumin (Fig. 2C). In FRALB < 3, no detectable intact albumin was present (Fig. 2D).

FPLC elution profile of albumin and FRALB. Analysis of 100 μg of albumin (A), unpurified FRALB (B), FRALB < 50 (C), and FRALB < 3 (D). AU = arbitrary units.
In Vitro Studies
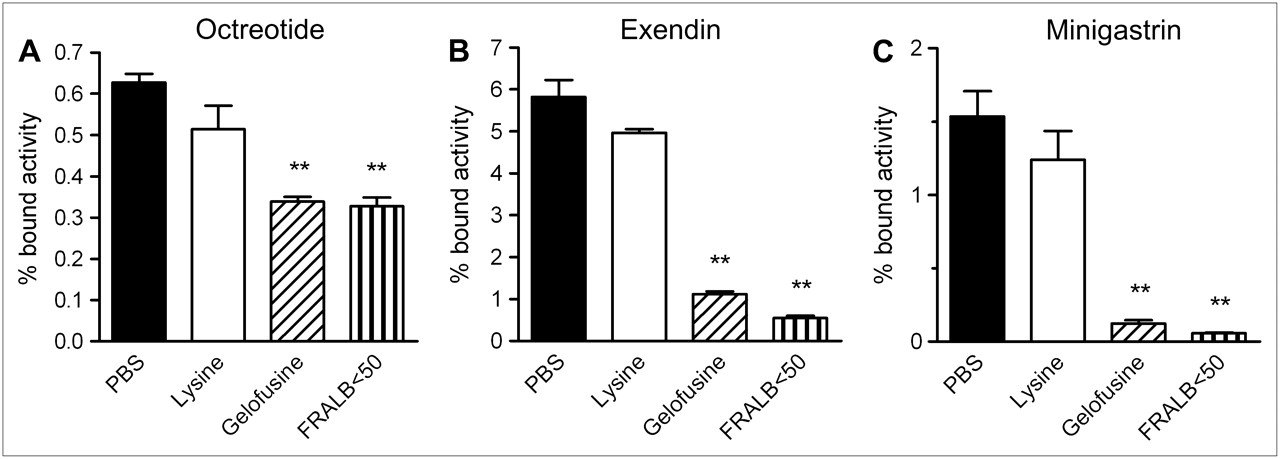
The BN16 assay showed that FRALB < 50 decreased 111In-octreotide binding by 48% ± 5.7% (P < 0.001), as illustrated in Figure 3A. Gelofusine decreased uptake by 46% ± 3.0% (P < 0.001). Lysine did not significantly reduce the binding of octreotide in this assay (P = 0.14).

Inhibition of binding or uptake of 111In-octreotide (A), 111In-exendin (B), and 111In-minigastrin (C) in BN16 cells by lysine, Gelofusine, or FRALB < 50. Results are presented as mean percentage of bound activity; error bars indicate SEM. *P < 0.05; **P < 0.005.
The binding of 111In-exendin to BN16 cells was decreased by Gelofusine and FRALB < 50 by 81% ± 1.9% (P < 0.001) and 91% ± 1.3% (P < 0.001), respectively (Fig. 3B). Lysine did not significantly reduce 111In-exendin binding (P = 0.10).
The binding of 111In-minigastrin was reduced by 96% ± 0.5% by FRALB (P < 0.005) and by 92% ± 2.7% by Gelofusine (P < 0.005) and was not decreased significantly by lysine (Fig. 3C). Higher concentrations of lysine, up to 2 mg (106-fold molar excess), did not reduce binding significantly either (data not shown).
Biodistribution of Albumin and FRALB
Measurement of tissue radioactivity concentration in rats 20 h after injection of 111In-labeled albumin or FRALB-3–50 demonstrated that kidney retention of 111In-albumin was significantly lower than retention of 111In-FRALB (2.47 ± 0.28 %ID/g vs. 15.2 ± 1.2 %ID/g, respectively; P < 0.0001, data not shown). Furthermore, the radioactivity concentration of 111In-albumin in the blood was significantly higher (0.069 ± 0.004 %ID/g vs. 0.011 ± 0.002 %ID/g; P < 0.0001, data not shown), indicating faster blood clearance of the low-molecular-weight FRALB. Phosphor imaging autoradiography of kidney sections demonstrated that, after intravenous injection of 111In-albumin and 111In-FRALB, radioactivity was present mainly in the renal cortex, as shown in Figure 4. Autoradiography also confirmed that renal radioactivity concentration of 111In-FRALB was much higher than that of 111In-albumin.
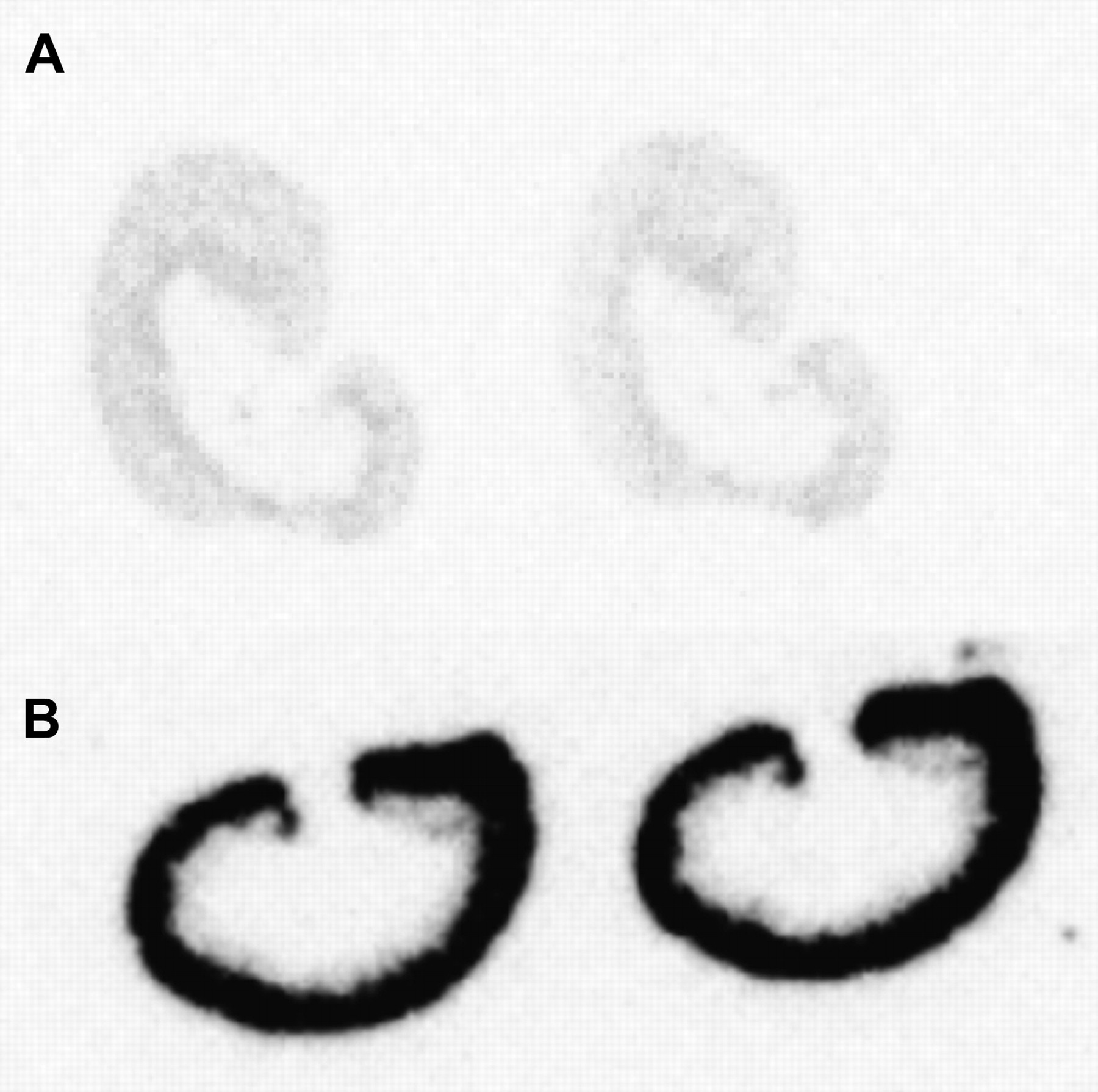
Autoradiography of renal sections of 111In-albumin (A) or 111In-FRALB-3–50 (B) dissected from rats, 20 h after injection. Radiolabeled compounds were present mainly in renal cortex.
Renal Uptake of Radiolabeled Peptides
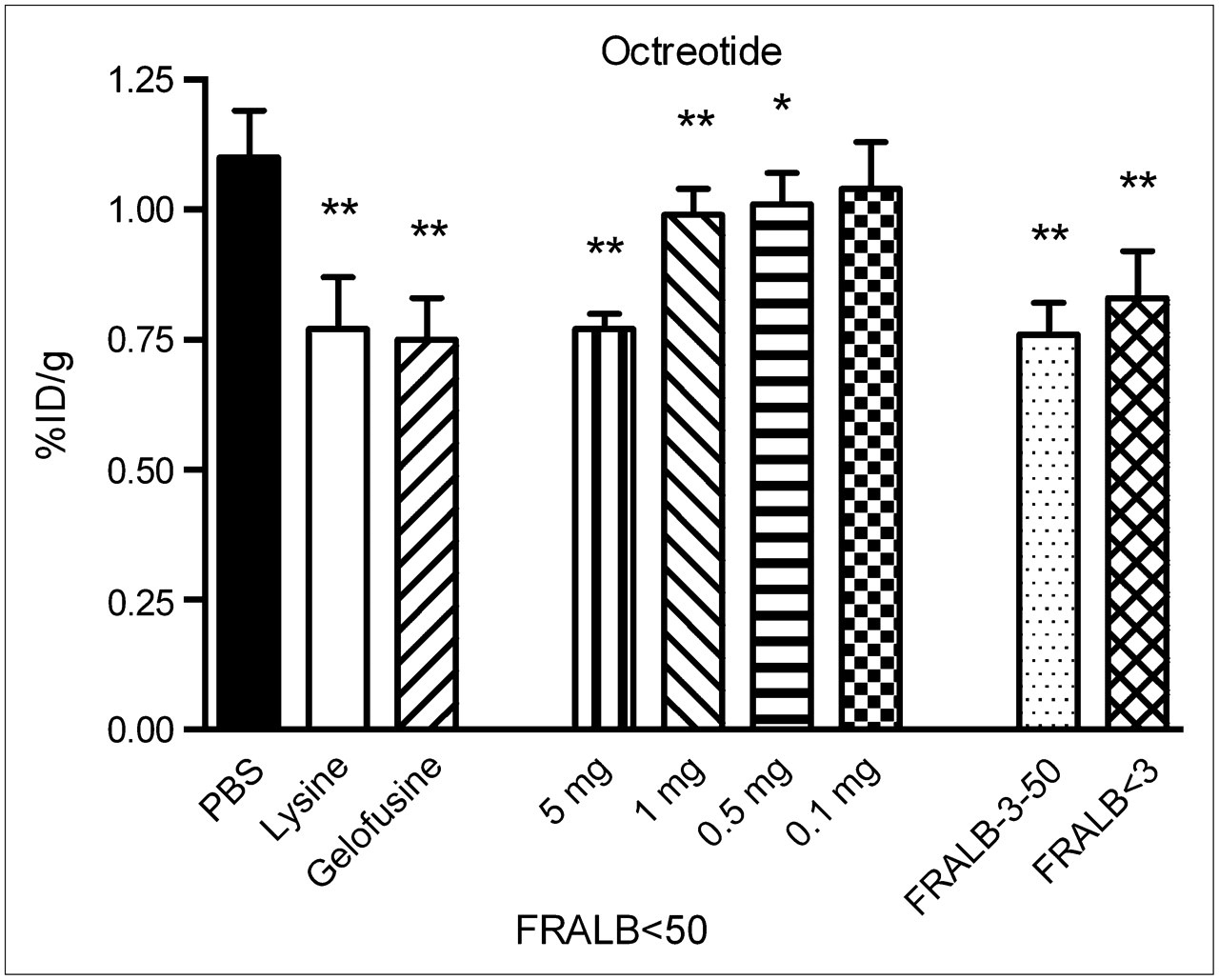
The effect of FRALB on renal uptake of 111In-octreotide is shown in Figure 5. A dose of 5 mg of FRALB < 50 reduced renal uptake of 111In-octreotide by 30% ± 2.7% (P < 0.0001), which is similar to the effect of 20 mg of Gelofusine (32% ± 7.0%, P < 0.0001) and 80 mg of lysine (30% ± 9.1%, P < 0.0001). Doses of 1 and 0.5 mg of FRALB < 50 reduced uptake by, respectively, 10% ± 4.6% (P < 0.005) and 8.1% ± 5.5% (P < 0.05). The 2 other purified FRALB preparations (FRALB-3–50, 1 mg; FRALB < 3, 4 mg) both significantly reduced renal uptake of 111In-octreotide (P < 0.0001) and were as potent as 80 mg of lysine.
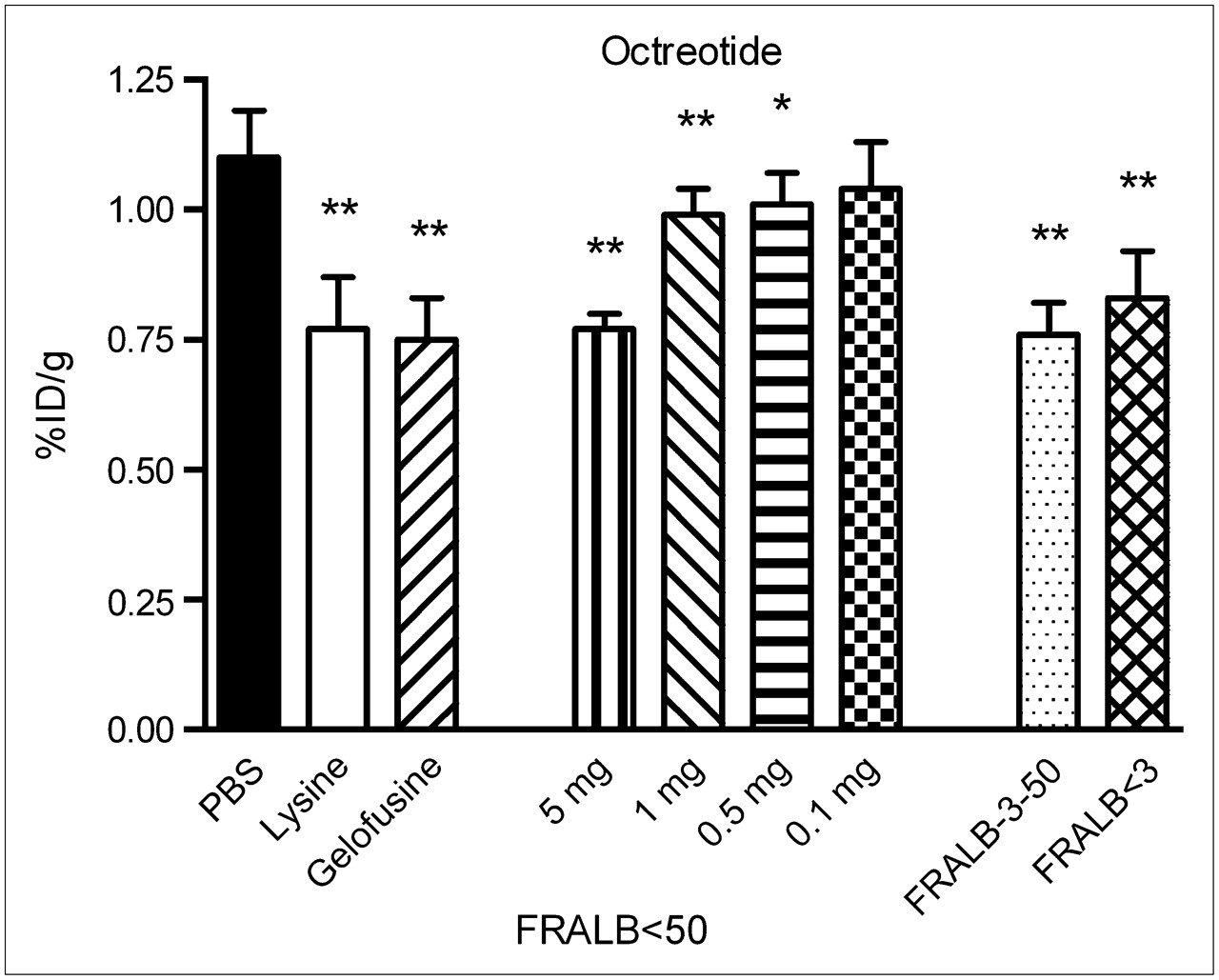
Kidney radioactivity concentrations 20 h after intravenous injection of 111In-octreotide in rats. Two to 4 min before injection of 111In-octreotide, groups of 4 rats received 0.5 mL of either PBS, lysine, Gelofusine, decreasing concentrations of FRALB < 50, or different filtered preparations of FRALB. Results are presented as mean %ID/g; error bars indicate SEM. *P < 0.05; **P < 0.005.
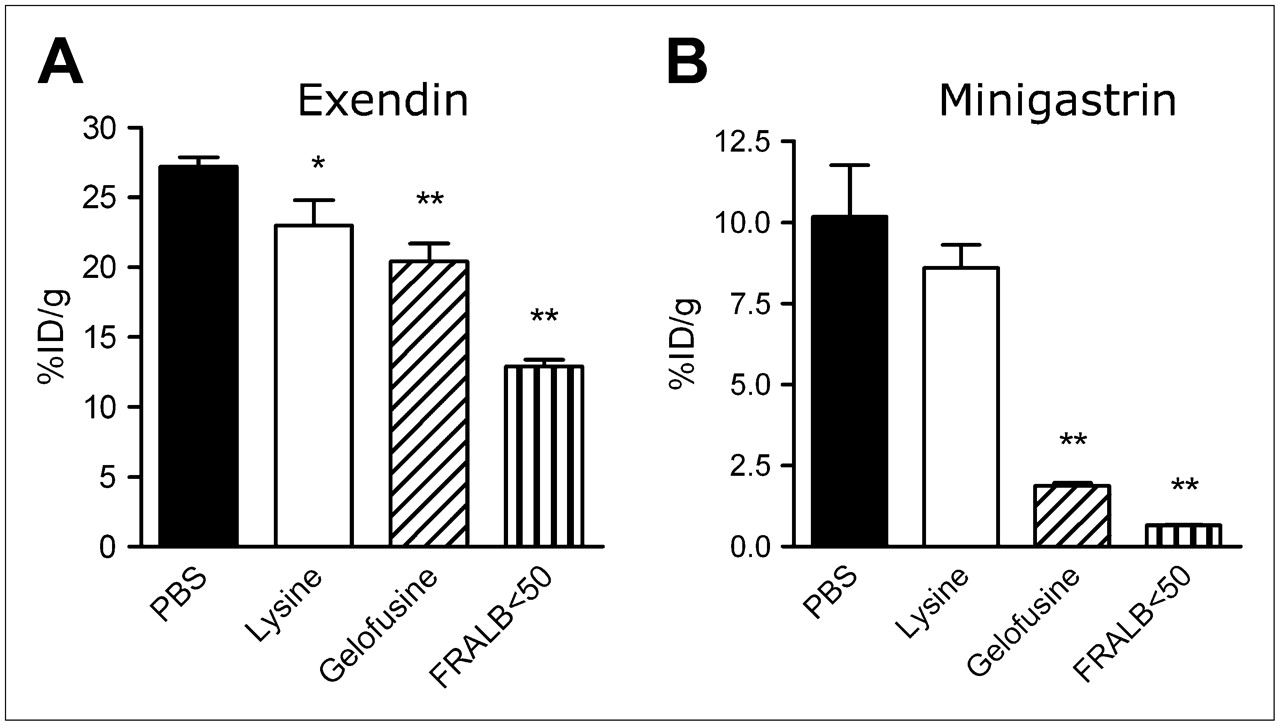
Renal uptake of 111In-exendin (Fig. 6A) was reduced by FRALB < 50 by 52% ± 4.7% (P < 0.0001), more potently than by either Gelofusine (25% ± 13%, P < 0.0005) or lysine (15% ± 18%, P < 0.05). Figure 6B shows that renal uptake of 111In-minigastrin was almost completely blocked by FRALB < 50 (93% ± 0.62% reduction, P < 0.0001). Reduction in uptake by Gelofusine was 82% ± 2.5% (P = 0.0001). Lysine did not reduce uptake of minigastrin significantly.

Kidney radioactivity concentrations 20 h after intravenous injection of 111In-exendin (A) or 111In-minigastrin (B) in rats. Two to 4 min before injection of radiolabeled peptide, groups of 4 rats received 0.5 mL of either PBS, lysine, Gelofusine, or FRALB < 50. Results are presented as mean %ID/g; error bars indicate SEM. *P < 0.05; **P < 0.005.
In all experiments the FRALB did not significantly affect uptake of 111In-octreotide in tissues and organs other than the kidneys (blood, spleen, adrenals, and pancreas), as determined by ANOVA with subsequent Bonferroni testing (data not shown).
DISCUSSION
In this study, we tested the hypothesis that FRALB, a natural ligand of megalin, might be a potent inhibitor of renal reabsorption of radiolabeled peptides. We hypothesized that FRALB may pass the glomerular membrane efficiently (18,20) and may subsequently competitively inhibit the megalin-mediated reabsorption of these peptides.
The in vitro experiments with 111In-octreotide, 111In-exendin, and 111In-minigastrin showed specific binding of exendin and minigastrin to megalin-expressing cells, supporting the assumption that megalin is involved in the binding and uptake of these peptides. However, a contribution of other mechanisms of uptake cannot be excluded. Relatively small amounts of FRALB inhibited binding of 111In-octreotide, 111In-exendin, and 111In-minigastrin to the BN16 cells at least as efficiently as Gelofusine. Remarkably, in vitro lysine did not reduce binding significantly.
The biodistribution studies of 111In-DTPA-albumin and 111In-DTPA-FRALB in rats support the hypothesis that FRALB passes the glomerular membrane more efficiently than does intact albumin. The renal activity concentration of FRALB was more than 5-fold higher than the concentration of intact albumin. Both albumin and FRALB were retained mainly in the renal cortex, the same anatomic location in which previous studies showed localization of 111In-octreotide, 111In-exendin, 111In-minigastrin, and megalin (21–24).
In our experiments in rats, coadministration of FRALB effectively reduced renal uptake of 111In-octreotide, presumably because of interaction of the albumin fragments with the megalin receptor in the kidneys. Barone et al. demonstrated that uptake of 111In-octreotide by opossum kidney cells was, at least partly, mediated by endocytosis via the megalin/cubilin receptor complex (25). The most convincing evidence was reported by de Jong et al., who showed that renal uptake of 111In-octreotide was very low in kidney-specific megalin knock-out mice (21) and that 111In-octreotide colocalized with megalin staining on murine kidney sections (21,22).
FRALB significantly reduced renal radioactivity concentrations of 111In-octreotide in a concentration-dependent manner. Fragments with a molecular weight between 3 and 50 kDa appeared to be most potent. Injection of as little as 1 mg of FRALB-3–50 per rat resulted in the same reduction in renal 111In-octreotide concentration as the administration of 20 mg of Gelofusine or 80 mg of lysine.
Gelofusine and FRALB efficiently reduced renal uptake of all studied peptides, whereas a high dose of 80 mg of lysine did not decrease uptake of 111In-minigastrin significantly. This confirms our previous hypothesis that inhibition of tubular reabsorption of radiolabeled peptides such as octreotide by basic amino acids may be the exception rather than the rule for radiopeptides (23).
The megalin receptor possesses 4 distinct negatively charged binding domains (26). This may explain why renal reuptake of some substances is blocked by lysine, whereas reuptake of other substances is not. Lysine and octreotide may interact with the same binding domain, whereas exendin and minigastrin may bind to 1 or more other domains. One or more components of FRALB and Gelofusine, consisting of various different smaller and larger molecules of different structure and charge, may interact with 2 or more of the binding domains of megalin, thus efficiently blocking renal reabsorption of various substances.
An alternative explanation could be that renal uptake of 111In-exendin and 111In-minigastrin is not inhibited by lysine, because uptake of these radiolabeled peptides may be mediated by mechanisms other than the megalin receptor. Whereas renal uptake of 111In-octreotide has been shown to be, at least partially, mediated by megalin (21,22), the role of megalin in uptake of exendin and minigastrin is not yet established. Our group (23) and Melis et al. (24) have previously suggested the possible involvement of various different uptake mechanisms in the tubular reabsorption of radiolabeled peptides, such as organic anion transporters, cubilin, and fluid-phase endocytosis. We are currently conducting further studies into the mechanisms of renal reuptake of these radiolabeled peptides.
The effect of FRALB on the reduction of renal reabsorption of 111In-octreotide, 111In-exendin, and 111In-minigastrin in this study was equal to or more pronounced than the effects of either Gelofusine or lysine. On a mass basis, the administered dose of FRALB that induced this effect was 5–20 times lower than the Gelofusine dose and 16–80 times lower than the lysine dose. Using FRALB, more potent inhibition of uptake may be achievable with less chance of side effects. However, although intact albumin is not toxic under physiologic conditions, albumin-derived peptides could be toxic as they have physical and chemical characteristics different from those of albumin. Unlike albumin, these peptides can reach relatively high concentrations in the glomerular filtrate on intravenous injection. Obviously, toxicity studies of FRALB must be performed before it can safely be used in patients.
The FRALB used in these studies, obtained by trypsinization of albumin, contains a poorly defined mixture of many different albumin fragments. To identify the most potent inhibitor of renal reuptake of radiolabeled peptides and to minimize the chance of unwanted side effects, further studies are warranted to purify FRALB and to further clarify its mechanism of action.
CONCLUSION
Renal uptake of 111In-octreotide, 111In-minigastrin, and 111In-exendin is effectively reduced by administration of relatively low amounts of FRALB. The administration of 1 mg of FRALB with a molecular weight between 3 and 50 kDa was as effective in reducing the renal uptake of 111In-octreotide as an administration of 80 mg of lysine. Further studies will be performed to identify which specific albumin fragments are responsible for this effect.
Acknowledgments
We thank Gerry Grutters, Maarten Brom, and Bianca Lemmers (University of Nijmegen, Central Animal Laboratory) for their expert technical assistance in the animal experiments.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication April 8, 2008.
- Accepted for publication May 28, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Reduced Renal Uptake of Various Radiopharmaceuticals with Sodium Paraaminohippurate Coadministration in a Rat Model
- Safety and Efficacy of Para-Aminohippurate Coinfusion for Renal Protection During Peptide Receptor Radiotherapy in Patients with Neuroendocrine Tumors
- Succinylated Gelatin Improves the Theranostic Potential of Radiolabeled Exendin-4 in Insulinoma Patients
- Dual-Target Binding Ligands with Modulated Pharmacokinetics for Endoradiotherapy of Prostate Cancer
- Megalin Contributes to Kidney Accumulation and Nephrotoxicity of Colistin
- Imaging of Human Epidermal Growth Factor Receptor Type 2 Expression with 18F-Labeled Affibody Molecule ZHER2:2395 in a Mouse Model for Ovarian Cancer
- 64Cu-Labeled Peptide for PET of Breast Carcinomas Expressing the Thomsen-Friedenreich Carbohydrate Antigen
- Renal Toxicity of Radiolabeled Peptides and Antibody Fragments: Mechanisms, Impact on Radionuclide Therapy, and Strategies for Prevention