


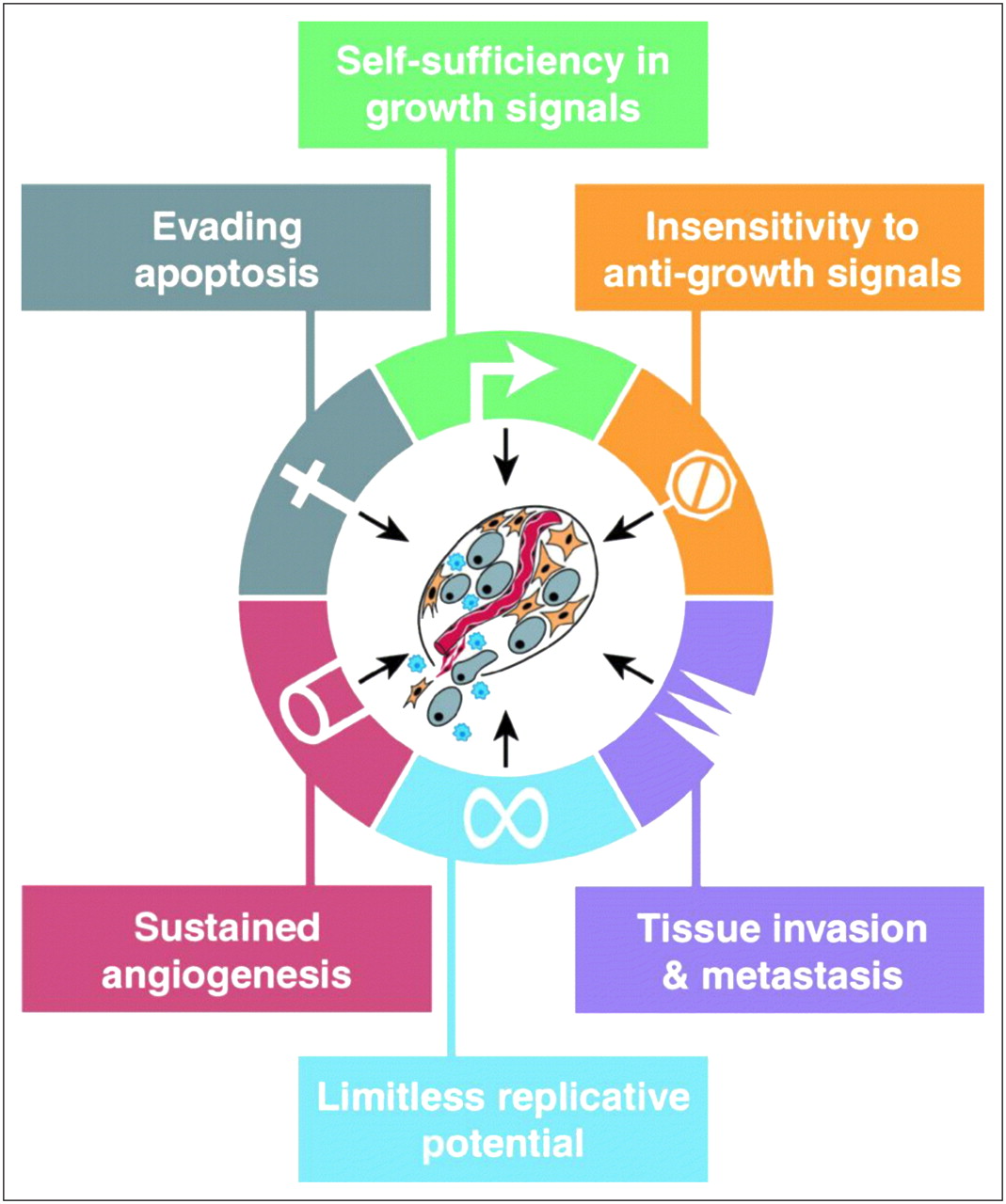
Perhaps the best way to understand cancer is to think of cancer as an organ, just like other organs such as the lungs, kidneys, or liver. Although the cancer organ arises within other organs it does in fact have the complexity of other organs. Instead of simply thinking about cancer cells in isolation, by viewing cancer as an organ we can best dissect this complex entity and truly understand, diagnose, and manage it. We now better grasp that understanding the role of angiogenesis (formation of new blood vessels to feed the tumor), the presence of both cancer cells and cancer stem cells, the interaction of cancer cells and stromal elements, and the interaction of the immune system with all elements of the cancer organ is critical to understanding the complexity of cancer. In its most basic form, cancer is a failure in tissue homeostasis (cell growth vs. cell death). Cancer cells do not respond to signals that usually regulate cell growth and division. These cells grow unchecked, producing more and more cancer cells. Hanahan and Weinberg (1) summarized the biology of tumor cells and categorized the 6 distinct hallmarks of cancer (Fig. 1). Specifically, these are self-sufficiency in growth signals, insensitivity to growth-inhibitory (antigrowth) signals, evasion of programmed cell death (apoptosis), limitless replicative potential, sustained angiogenesis, and tissue invasion and metastasis. Perhaps a seventh hallmark that is not explicitly diagrammed is an ability of cancer cells to evade the immune system. Also, an eighth hallmark not made explicit is aerobic glycolysis of cancer cells. Taken together, these 8 hallmarks help one to understand and tackle the diagnosis and management of cancer in a logical and unified manner.

The 6 hallmarks of cancer unifying common phenotypes that cancer cells take on to survive and metastasize. (Reprinted with permission of (1).)
If cancer diagnosis and management are going to be fully advanced through molecular imaging, we must be able to marry fundamental advances in the understanding of cancer as an organ with imaging strategies that can interrogate all aspects of the cancer organ. In this supplement, we review the major areas in which molecular imaging of cancer management has started to make some progress. Although many other areas could also easily be a part of this supplement, the areas reviewed should familiarize the reader with the state of the art of the predominant strategies for using molecular imaging in the management of cancer. A multimodality perspective is used, in which both radionuclide-based methods with SPECT and PET and nonradionuclide-based methods including MRI, MR spectroscopy, optical imaging, and ultrasound imaging are discussed for each major area. Although the radionuclide-based approaches are generally the most advanced in the clinical setting, nonradionuclide-based strategies have significant potential, as has been currently validated predominantly in preclinical models. The necessity of using anatomic imaging with CT or MRI in conjunction with one or more molecular imaging strategies is still important. The review articles that discuss imaging agents map nicely to the 8 hallmarks of cancer. This mapping was done to truly show how molecular imaging is fundamentally being married to the latest advances in cancer cell biology.
We first begin with Dr. Pichler et al. with a review on the instrumentation of molecular imaging (2). Although most molecular imaging strategies to date have been based on single-photon and positron-emission imaging agents, there is still some potential for nonradionuclide strategies such as those that use MRI, optical imaging, and other established or emerging modalities. Although the practice of nuclear medicine currently revolves around PET/CT and SPECT, it will be important for readers to continue to monitor the progress of other modalities that may eventually play a significant role in the molecular imaging of cancer. The emergence of PET/MRI and hyperpolarized MRI as potential strategies for molecular imaging is highlighted. My belief is that multimodality molecular imaging will eventually be critical to interrogating cancer and that even newer modalities that have yet to be clinically translated (e.g., photoacoustic molecular imaging) will eventually play important roles. The chances that any one modality or even a combination of modalities (e.g., PET/CT) will become dominant for each and every clinical cancer application remain low. The Swiss army knife for molecular imaging remains elusive, but by using several knives at our disposal we may yet be able to effectively, noninvasively slice the cancer organ.
The review article by Dr. Gillies et al. (3) tackles the relatively difficult area of understanding tumor glucose metabolism by asking not only how tumors have increased glycolysis but perhaps more importantly why. The authors propose that the phenotype of elevated glucose consumption in cancers is selected through somatic evolution because it leads to increased acid production. They argue that production of acid is a key mechanism by which cancer spreads beyond the sites where it originates. They propose that all commonly observed traits must confer specific adaptive advantages to cancer cells as the cells progress. In other words, the hallmarks of cancer (Fig. 1) can be viewed as the phenotypic adaptations necessary to overcome all the proliferation barriers that evolving tumor populations encounter during their progression through carcinogenesis. Although one might not agree with the why of increased glycolysis, the fact that it is a hallmark allows one to tackle diagnosis and management in ways that molecular imaging has already taken advantage of (e.g., 18F-FDG PET/CT).
Drs. Plathow and Weber (4) review the molecular imaging of tumor metabolism. Because alterations in metabolism are a key hallmark, the molecular imaging of these alterations is key to the management of cancer. The authors summarize recent data on the molecular mechanisms underlying the increased metabolic activity of cancer cells and discuss imaging techniques for studies of tumor glucose, lipids, and amino acid metabolism. In contrast to other molecular imaging probes (e.g., targeting of a cell surface receptor by an imaging agent), metabolic imaging probes do not target the expression of one molecule but provide information about the integrated function of multiple transporters and enzymes involved in a metabolic process. This provides a unique look at a cancer phenotype that is not available through any other mechanism. Due in large part to the tremendous success of 18F-FDG PET/CT, we must remain vigilant about looking for other imaging probes to measure metabolic characteristics of cancer, especially in areas where 18F-FDG PET has some difficulties. This search may eventually lead to a “Super-FDG” and other imaging agents with applications in a given subset of cancer patients.
Drs. Bading and Shields (5) provide a review on imaging cell proliferation. Cell proliferation with synthesis of new DNA is a key hallmark of cancer. Enormous effort has been expended in this area, with many PET agents developed, which are reviewed in this article. These include 11C-thymidine, 3′-deoxy-3′-fluorothymidine (FLT), and 1-[2′-deoxy-2′-fluoro-1-β-darabinofuranosyl]-thymine. The early clinical data with FLT demonstrate that its uptake correlates well with in vitro measures of proliferation. Although FLT can be used to detect tumors, its tumor–to–normal-tissue contrast is generally lower than that of 18F-FDG in most cancers outside the brain. Its greatest potential will be in monitoring response to therapy, and clinical trials are clearly needed to further clarify the limitations of these approaches.
Dr. Blankenberg (6) provides an update on imaging of apoptosis (programmed cell death). The dysregulation of apoptosis can lead to the growth of tumors (too little apoptosis). For cancer, the imaging of apoptosis remains an important potential method for monitoring the efficacy of various anticancer therapies, including chemotherapy and radiation therapy, as well as of new targeted drugs. One key challenge that remains for the imaging of apoptosis is the lag time between exposure to the apoptotic trigger and the time of observable morphologic signs of apoptosis. This time is highly variable and depends on cell type, type of trigger, intensity and exposure, duration of trigger, and the local environmental conditions of the cells. This variability continues to challenge the implementation of clinical trials to validate various apoptosis-imaging strategies. The article also reviews the potential for use of multimodality imaging strategies for imaging apoptosis with MR spectroscopy, which likely has limited sensitivity, and ultrasound, which will require much greater validation.
Dr. Winnard et al. (7) provide an update on imaging metastatic potential and imaging metastases themselves. Metastatic potential, in particular, as something that would be difficult to assess with conventional anatomic imaging strategies, is ideally suited for molecular imaging. Metastatic potential is also one of the most critical challenges, because it is the spread of cancer cells to sites distant from their site of origin—the usual reason for death by cancer. The article reviews matrix metalloproteinases, which are proteolytic enzymes that catalyze the breakdown of the extracellular matrix and are involved in tumor growth and invasion. The use of imaging agents targeting matrix metalloproteinases is a possible new method for imaging metastatic potential but has its own challenges, including the need for signal amplification. The article also reviews the ability to use other cancer imaging strategies (e.g., angiogenesis and hypoxia) that may assess metastatic potential. As a community, we must develop better methods to determine metastatic potential, and this article lays the foundations for helping us to do so.
Drs. Cai and Chen (8) review the rapid progress to date in imaging tumor angiogenesis. Because the growth of tumors is critically dependent on the growth of new blood vessels, a unique opportunity exists to image angiogenesis. The authors review several different strategies for imaging tumor angiogenesis with receptors such as αvβ3 integrins and the vascular endothelial growth factor receptor. The use of small molecules, antibodies, and nanoparticles is discussed. Although few of the strategies have been clinically translated to date, the likelihood is strong that a multimodality approach to the imaging of tumor angiogenesis will occur over the next 5 years. This will be critical, especially because many anticancer strategies targeting angiogenesis have already been developed and more will likely continue to be developed.
Dr. Krohn et al. (9) review the molecular imaging of hypoxia, which continues to be a rapidly expanding area of investigation for the molecular imaging community. Oxygen is an essential nutrient for mammalian cells because of its role as the terminal electron acceptor in oxidative phosphorylation. Yet as cancer cells grow, they tend to outgrow their vascular supply, leading to hypoxia (oxygen levels insufficient to support metabolism). Several PET tracers for the imaging of hypoxia are reviewed (e.g., fluoromisonidazole and copper-diacetyl-bis(N4-methylthiosemicarbazone)), along with nonradionuclide-based strategies. In addition, the relative advantages and disadvantages of each available strategy are presented in order to assess future studies that are still needed to make hypoxia imaging routine and informative. The growing body of literature shows that hypoxia identified by imaging is predictive of poor survival in numerous cancer settings. Studies are still needed to evaluate the predictive value of hypoxia imaging in tumors so as to provide convincing data on the extent to which it is an independent predictive marker.
Dr. Mankoff et al. (10) review tumor receptor imaging. Perhaps one of the most challenging areas for molecular imaging is receptor imaging because of the relatively low levels of target available in many cases and the necessity to not saturate all receptors with the imaging agent so as to not perturb the underlying cells being imaged. The authors first review aspects of receptor biochemistry and biology that underlie tumor receptor imaging, using the estrogen receptor system as a prototype system. Examples of progress in radionuclide receptor imaging in 3 receptor systems—steroid receptors, somatostatin receptors, and growth factor receptors—are highlighted, along with a review of recent investigations of receptor imaging using other molecular imaging modalities. Applications of predicting response to receptor-targeted therapies by first assessing receptor levels using molecular imaging are also highlighted.
Drs. Kang and Chung (11) review the important emerging area of the imaging of reporter gene expression. These strategies require an introduction of one of several available reporter genes into cells of interest (e.g., immune cells) followed by imaging with a reporter probe that is trapped primarily in cells that express the reporter gene. These strategies are being used in many preclinical models and have also been translated into pilot clinical applications of imaging human gene therapy with PET. One of the most important applications of these reporter strategies may be related to the ability to monitor the interaction of immune cells with tumors. Adoptive immunotherapy, where we introduce into cancer patients genetically engineered immune cells (e.g., T cells), may be successful only if these same cells can be marked with reporter genes so that they can be imaged when they are trafficking and expanding in patients. Although these strategies are still in their infancy, Drs. Atkins and Dubey et al. (12) review the potential for making significant advances if molecular imaging is properly married to adoptive immunotherapy. Many scientists, including myself, believe that eventually cancer will have to be dealt with as a chronic disease and that modulation of our own immune system will be needed to keep cancer cells in check. As with all therapies, individualization of the therapy through imaging will be the likely route to true success.
A key challenge for the molecular imaging community lies not only in validating all the above strategies for imaging cancer as an organ but also in selecting a given strategy for a specific patient management problem. This is especially true in that no imaging methods currently allow high multiplexing of molecular imaging strategies in patients. For example, in a patient diagnosed with non–small cell lung cancer, it would not be easy to image tumor angiogenesis (e.g., vascular endothelial growth factor receptor, αvβ3), levels of hypoxia, glucose use, levels of epidermal growth factor receptors, and apoptosis after treatment with a new-generation receptor-targeted drug. Although molecular imaging strategies continue to evolve, it is unlikely that in the clinical setting we can easily interrogate (simultaneously or near-simultaneously) all the hallmarks of cancer. It is unlikely that we can cost-effectively image all the hallmarks of cancer for every cancer patient for initial characterization, as well as for predicting and monitoring response to treatment. We are forced to choose the molecular imaging applications carefully, as well as clinically validate imaging approaches in a sensible fashion. As a community, we should not shy away from these difficult challenges and instead must embrace them and provide logical and cost-effective solutions so that molecular imaging may be properly used.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication April 22, 2008.
- Accepted for publication April 23, 2008.





{kind=link}