


Abstract
Nicotinic acetylcholine receptors (nAChRs) are of great interest because they are implicated in various brain functions. They also are thought to play an important role in nicotine addiction of smokers. Chronic (−)-nicotine, a nAChR agonist, treatment in mice and rats elicits a dose-dependent increase in nAChRs in the brain. Upregulation of nAChRs in postmortem human brains of smokers has also been reported. However, changes in nAChRs after cigarette smoking cessation in humans are poorly understood. The aim of this study was to detect the dynamic changes of nAChRs after smoking and smoking cessation in the brains of living subjects. Methods: We performed 5-123I-iodo-A-85380 (123I-5IA) SPECT on nonsmokers and smokers (n = 16) who had quit smoking for 4 h, 10 d, and 21 d and calculated and compared distribution volumes (Vt) of 123I-5IA. Results: The binding potential of nAChRs (Vt of 123I-5IA) in the brains of smokers decreased by 33.5% ± 10.5% after 4 h of smoking cessation, increased by 25.7% ± 9.2% after 10 d of smoking cessation, and decreased to the level of nonsmokers after 21 d of smoking cessation. Conclusion: Because the upregulation of the nAChRs of the smokers after chronic exposure of the nicotine was downregulated to the nonsmokers' level by around 21 d after smoking cessation, the upregulation is a temporary effect. The decrease in nicotinic receptors to nonsmoker levels may be the breaking point during the nicotine withdrawal period.
Nicotinic acetylcholine receptors (nAChRs) are a family of ligand-gated ion channels that regulate neurotransmission in the central and peripheral nervous systems. These receptors are of great interest because they are implicated in various brain functions, including cognition and memory (1,2) and in nicotine-induced neuroprotective (3) and analgesic effects (4). In addition, these receptors are thought to play an important role in nicotine addiction (5).
Chronic treatment with agonists for most neurotransmitter receptor systems results in a decrease in receptor number. However, it has been demonstrated that chronic treatment of mice (6) and rats (7) with (−)-nicotine, an nAChR agonist, elicits a dose-dependent increase in nAChRs. This upregulation is not permanent, returning to control levels within 7–10 d in mice (6) and 15–20 d in rats (8,9) after cessation of (−)-nicotine treatment. Previous efforts to demonstrate nAChR upregulation in the human brain have also been reported primarily in in vitro binding assays (10,11). Kassiou et al. reported the upregulation of nAChRs with chronic (−)-nicotine treatment in the brain of a live baboon (12). More recently, Staley et al. described the upregulation of nAChRs in human brains after early abstinence of tobacco smoking using 5-123I-iodo-A-85380 (123I-5IA) and SPECT images (13). However, changes in nAChRs in humans after cessation of smoking are poorly understood. Breese et al. studied the levels of 3H-nicotine binding in humans postmortem for changes in nicotinic receptor levels and reported that the nAChR levels in smokers who had stopped smoking at least 2 mo before their death were similar to those in nonsmokers (14); the effects of shorter-term smoking cessation are unknown.
123I-5IA is a nAChR imaging probe that has extremely high selectivity and specificity for the α4β2 subunit of nAChRs in rodent (inhibition constant = 0.37 nM) (15) and primate brain in vivo (16), with relatively low acute toxicity (effective dose equivalent = 30 μSv/MBq) (17,18). Moreover, we have developed the methodology for the quantification of nAChRs in human brain using 123I-5IA and SPECT (19).
The aim of the present study was to detect the dynamic changes of nAChRs in living human brain after smoking and smoking cessation. We performed 123I-5IA SPECT on nonsmokers and smokers who had quit smoking for 4 h, 10 d, and 21 d and compared the distribution volumes (Vt) of 123I-5IA of each group and with nonsmokers. To our knowledge, this is the first in vivo imaging study of nAChR upregulation and recovery in response to short-term smoking cessation in living subjects.
MATERIALS AND METHODS
Volunteers
Six male nonsmokers (23 ± 6 y) and 10 healthy male smokers (28 ± 4 y) were included in this study. Five smokers in the 4-h group were also included in either the 10-d or the 21-d group. In total, 21 123I-5IA SPECT studies were acquired (Table 1). None of the subjects had a history of neurologic or psychiatric illness or the use of psychotropic or sleep-inducing drugs. The nonsmokers had no history of smoking tobacco.
Study Groups
For the smoking withdrawal studies, the smokers were divided in 3 groups: 5 subjects (age, 28 ± 4 y) for 4-h withdrawal, 5 subjects (age, 27 ± 6 y) for 10-d withdrawal, and 5 subjects (age, 28 ± 3 y) for 21 d of smoking withdrawal. The 4 groups were age-matched.
All subjects gave written informed consent to participate in this study in compliance with the regulations of the Joint Committee on Clinical Investigation of the Kyoto University Hospital.
Radiolabeling
Radiolabeling of the 123I-5IA followed the methods we reported previously (19). To a sodium 123I-iodide solution (1,110 MBq) (Nihon Medi-Physics), 100 μg of (S)-5-(tri-n-butylstannyl)-3-([1-t-butoxycarbonyl-2(S)-azetidinyl]methoxy)pyridine, 1.5% acetic acid, 3 mol/L HCl, and 5% H2O2 solution were added, and the mixture was stirred at 75°C for 15 min. Concentrated HCl was then added, and the resulting solution was stirred for another 10 min at 75°C. The mixture was made basic with NaOH and extracted with ethyl acetate, and the organic layer was evaporated. The residue was purified by reverse-phase high-performance liquid chromatography ([HPLC] Cosmosil 5C18-AR-300, 10 × 250 mm; Nacalai Tesque; 10 mmol/L ammonium acetate/methanol/triethylamine = 752:750:2; 1.5 mL/min; retention time for 5IA was 40 min). After evaporation of the HPLC eluent, the residue was dissolved in 0.9% saline and filtered through a 0.2-μm filter into a sterile vial. Radiochemical purity was >98%, and radiochemical yields were ∼42%. The specific activity determined from the ultraviolet absorbance at 254 nm was >169 GBq/μmol (the detection limit for this method).
SPECT
All subjects underwent a set of 5 SPECT dynamic scans (a 120-min scan, followed by 4 sets of 20-min scans). All SPECT dynamic scans were acquired with a triple-head rotating γ-camera system (PRISM 3000; Picker International) equipped with low-energy, high-resolution, fanbeam collimators. Data acquisition and image reconstruction were performed as in our previous study (19). The data acquisition was alternately performed over 120 min after intravenous injection of 123I-5IA, followed by 4 sets of 20-min scans (at 3, 4, 5, and 6 h after the injection). SPECT images were reconstructed using a filtered backprojection algorithm with a ramp filter. Attenuation correction was performed using ellipses outer line approximation and Chang's method (coefficient of 0.06/cm), which assumes that the attenuation process is homogeneous throughout the brain and can be described by an exponential function. Scatter correction was not applied.
A dose of ∼150 MBq of 123I-5IA was administered intravenously over a period of 1 min at a constant rate with an infusion pump, and the SPECT scan was started at the same time as the injection. Arterial blood sampling and metabolite correction were also performed to estimate the arterial input function of the 123I-5IA for each volunteer by the same method as that used in our earlier study (19).
Arterial Input Function
Twenty-five arterial blood samples were obtained at the same time points as described previously (19). From each sample, 100 μL of plasma were removed and the radioactivity was measured in an automatic well-type γ-counter (Cobra 2; Packard Instruments). Sixteen samples were analyzed by thin-layer chromatography (TLC) (10% ammonium acetate and methanol [1:1], LK6DF Silica Gel, 60 Å; Whatman) for metabolite determination (Rf = 0.55 for 123I-5IA) (19). The measured unmetabolized fractions were fitted with a dual exponential curve, and the input function was calculated as all plasma sample counts were corrected for metabolites using the fitted curve.
Data Analysis
Reconstructed SPECT images were automatically coregistered using a coregistration algorism of statistical parametric mapping, SPM99 (Welcome Department of Cognitive Neurology, London, U.K.), to minimize positional error caused by head movement during the scans. Multiple circular regions of interest (ROIs) (21 pixels per circle) were manually drawn in each brain region (basal ganglia, thalamus, brain stem, cerebellum, frontal, parietal, temporal, and occipital cortices) on both sides. ROI data were further decay-corrected. SPECT data were calibrated to the well counter used to measure the injected activity. Time–activity curves were generated from the ROIs and the dynamic image datasets.
Kinetic analysis of the 123I-5IA was performed using a 2-compartment model including K1 and k2 rate constants and a curve-fitting method following our previous study (19). Vt values of the 123I-5IA were calculated and used as a quantitative index correlated with the regional binding potential of the nAChRs. The Vt values were further evaluated in terms of interval change after the smoking withdrawal.
Statistical Analysis
All data are expressed as the mean ± SD. The Vt values obtained from the different regions in the brain were analyzed by 1-way ANOVA with the Bonferroni protected least significant difference test. The interval changes of the 123I-5IA Vt were analyzed between the 3 phases after the smoking withdrawal using the Tukey−Kramer multiple comparison test. All tests were 2-sided, and probability values of P < 0.05 were considered significant.
RESULTS
As in our previous study (19), the characteristics of the arterial input functions for all volunteers (nonsmokers and smokers) were similar. The peak plasma activity occurred between 70 and 80 s after injection in all subjects and decreased rapidly to 6.5%−9.0% of the peak level in 10 min. Analysis of the unmetabolized compound by TLC demonstrated a high parent fraction of 123I-5IA in the plasma (87.7% ± 6.3%) in the first minute. 123I-5IA was rapidly metabolized, and the unchanged fraction represented 50.9% ± 8.8% and 32.4% ± 12.6% of total plasma activity at 20 and 60 min, respectively.
Figure 1 shows the representative standardized time–activity curve of 123I-5IA in the frontal cortex. The concentrations of radioactivity were slightly higher in the nonsmokers and in the 4-h smoking-cessation group followed by the 10-d and 21-d smoking-cessation groups. The peaks of radioactivity occurred ∼50 min after injection of 123I-5IA for nonsmokers and for the 4-h and 21-d smoking-cessation groups, whereas it was at ∼70 min for the 10-d smoking-cessation group. A differential dissociation of 123I-5IA from the binding sites was noted in the brain. The 4-h smoking-cessation group showed a faster dissociation compared with that of the nonsmokers. However, the 10-d and 21-d smoking-cessation groups showed a slower dissociation than that of the nonsmokers (more pronounced in the 21-d group). These findings reflected a temporal change of the nAChRs in the human brain.

Representative standardized time–activity curves of 123I-5IA in frontal cortex from a nonsmoker and smokers.
Packs per day and pack years of cigarette smoking before cessation were similar for the different groups of smokers (Table 2). Only 2 subjects (subjects 7 and 11) had detectable amounts of nicotine in their plasma after 4-h smoking cessation (Table 2).
Characteristics of Volunteers and Plasma Concentration of Nicotine and Cotinine
To validate the Vt values of the nonsmokers as a baseline group, we compared (t test) our current data (nonsmokers) with our published data (19). No significant difference was observed between these groups. Similar findings were also seen for K1 and k2. Therefore, we used the Vt values from nonsmokers as a reference for further comparisons with groups of smokers at several smoking-cessation intervals.
Table 3 describes the Vt values of different groups of volunteers (nonsmokers and smokers). There was a significant difference among those groups (ANOVA; P < 0.001). Individualized comparisons between 2 groups of volunteers were also performed. After 4 h of smoking cessation, the Vt values in all brain regions decreased significantly compared with those of the nonsmoker group (P < 0.05, except for frontal, parietal, and occipital cortices). On the other hand, after 10 d of smoking cessation, the Vt values were significantly higher than those of nonsmokers (P < 0.05, except for basal ganglia and thalamus). Then, after 21 d of smoking cessation, the Vt values decreased significantly compared with those of the 10-d group (P < 0.01, except for thalamus), returning to the level in nonsmokers (Vt values did not show any significant difference compared with those in the nonsmokers). Figure 2 shows the percentage of reduction and increment in each group of smokers in comparison with the nonsmoker group. In the Tukey−Kramer multiple comparison test, the interval changes of the 123I-5IA Vt between the 3 phases after the smoking cessation were significantly different (P < 0.001).

Percentage of reduction and increment of Vt of 123I-5IA in smokers after smoking cessation compared with nonsmokers. Eight brain regions are frontal, parietal, temporal, occipital, basal ganglia, thalamus, brain stem, and cerebellum from the left, respectively.
123I-5IA Vt Estimates for 2-Compartment, 2-Parameter Model for Various Brain Regions from Nonsmokers and Smokers After Withdrawal of Cigarette Smoking
The rate constant K1 had some fluctuations among the different groups of volunteers (nonsmokers and smokers); however, these differences were not statistically significant (ANOVA; not significant) (Fig. 3A). On the other hand, the values of the rate constant k2 were significantly different among the groups of volunteers (ANOVA; P < 0.01) (Fig. 3B). This difference was due basically to the increase of k2 in the group with 4 h of smoking cessation.
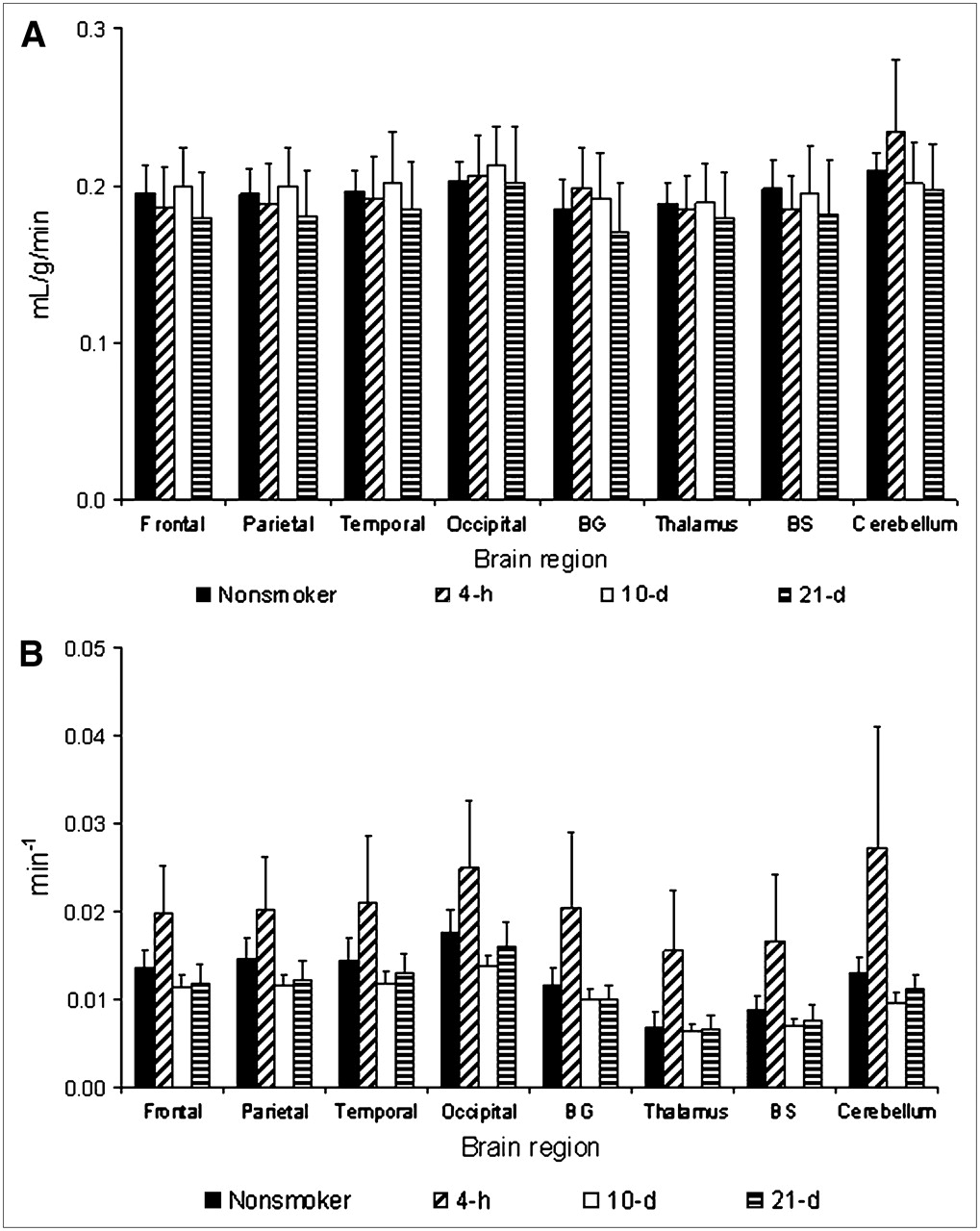
Temporal changes in rate constants K1 and k2 in nonsmokers and smokers after smoking cessation. (A) Rate constant K1 (mL/g/min). (B) Rate constant k2 (min−1). BG = basal ganglia; BS = brain stem.
DISCUSSION
The present study described the effect of nicotine intake in tobacco smokers and smoking cessation on the high-affinity nicotinic receptors in humans using 123I-5IA SPECT. To our knowledge, this is the first in vivo imaging of nAChR upregulation and recovery in response to short-term smoking cessation in living smokers.
Previous animal studies have shown that chronic nicotine treatment induces an increase in high-affinity nicotinic receptor binding (6–9), and human postmortem studies have found a similar increase in 3H-nicotine binding to high-affinity receptors in the postmortem cortex, cerebellum, and hippocampus of smokers compared with that in nonsmokers (10–12).
The mechanism by which the chronic exposure of nicotine evokes an increase in the density of the binding sites is not fully understood. Marks et al. reported that the increase in nicotinic receptor numbers in rodents is not caused by an increase in messenger RNA levels (20). The lack of an effect on nicotinic receptor transcription in mice suggests that nicotine-induced increases in nicotinic receptor levels are most likely related to a decrease in receptor turnover (21). The increase in nicotinic receptor number and the decreased rate of receptor turnover may be related to nicotinic receptor channel desensitization, which appears to reflect the conformational state of the receptor channel (21,22). Once the nicotinic receptor channels are desensitized and rendered inactive, additional receptors would be recruited to maintain the nicotinic response of the neuron, which results in an overall increase in nicotine binding, possibly due to a conversion of low-affinity receptors to a conformation with a high affinity for agonists (23).
In this study, the 123I-5IA Vt measured at 4 h after smoking cessation was significantly lower than that in nonsmokers. The mean value of the calculated Vt of the smokers was ∼33.5% ± 10.5% lower than that of nonsmokers and was more pronounced in the thalamus (51%) and brain stem (45%). In this group of volunteers, the plasma nicotine level 4 h after smoking cessation was detectable in only 2 subjects and was below the detection limit in the other subjects. Nicotine is highly lipophilic and demonstrated high levels of nonspecific uptake in brain (24,25). Rowell and Li have reported that levels of nicotine in the brain were ∼3-fold higher than those in the plasma (7), which explains the lack of plasma nicotine measurements in 3 subjects in this group of volunteers. Because of high levels of nonspecific uptake of nicotine in the brain, nicotine or its metabolites may accumulate in nonspecific compartments in the brain (i.e., white matter) and then diffuse slowly into areas with higher levels of nAChRs, maintaining high levels of occupancy of the nAChRs. In addition, Brody et al. have shown saturation of the nicotinic receptors in human brain for up to 4 h in smokers (26). Thus, we would expect some level of nicotine or metabolites in the brain that would compete with 123I-5IA and impair imaging of the upregulation of nAChRs. Also, we believe that some level of nicotine in the brain resulted in a high level of occupancy of the receptors, which reduced specific tracer uptake (27). Because of competitive binding between the radioligand and nicotine in the brain, imaging of upregulation of nAChRs in vivo requires sufficient time for nicotine clearance (>4-h smoking cessation).
After 10 d of smoking cessation, the Vt of 123I-5IA was significantly higher than that in nonsmokers, with a 25.7% ± 9.2% increase among brain regions. The result of the increased Vt of the 123I-5IA was in agreement with the upregulation of the nAChRs in the brains of smokers reported in postmortem human studies (10–12) and in animal studies (6–9). Staley et al. have described similar findings in human brain (13). The authors noted that after 6.8 ± 1.9 d of tobacco abstinence, the uptake of 123I-5IA increased significantly throughout the cerebral cortex (26%∼36%) in smokers (13). After 10 d of smoking cessation, nicotine and cotinine were not detected in the plasma. Thus, blood nicotine levels were negligible in the 123I-5IA SPECT scans of smokers as well as nonsmokers.
After 21 d of smoking cessation, the Vt of 123I-5IA was significantly lower compared with that after 10 d of smoking cessation and was not significantly different from that in the nonsmokers. Breese et al. showed that smokers who had quit at least 2 mo before death had nicotinic receptor binding levels similar to those in nonsmokers (14). In the present study, the interval of 21 d was thought to be the recovery time during which upregulated nAChRs return to the level of the nonsmoker. This suggests that nicotine-induced upregulation of receptor number is a temporary effect, similar to that found in rodents (28,29).
The upregulation of the nAChRs was similar in almost all brain regions, except the thalamus and basal ganglia, which showed a slightly different pattern. In thalamus and basal ganglia, after 10 d of smoking cessation the Vt was higher than that of the nonsmokers, as in the other regions, but was not significantly different. Staley et al. have shown similar findings (13). In addition, it has been reported in a study of mice that nicotine-induced increases in nicotinic receptor numbers do not increase to the same degree in all brain regions (30). Moreover, the nicotinic receptor is more abundant in the thalamus, with greater receptor heterogeneity, than in other brain regions (19). However, the characteristics of the acute response of neuronal nAChRs to nicotine depend on their subunit composition (31,32). Nicotinic receptor subtypes are affected differentially by chronic exposure to nicotine, both in cell models (33–35) and in vivo (35). Multiple factors are thought to be responsible for these differences.
This study should be interpreted in the context of several limitations. (a) The number of subjects evaluated was small, which reduced the statistical accuracy. (b) The study design used did not allow us to deal with a within-subject analysis of the whole group, as the variables were analyzed independently. (c) The smokers varied in their rate and depth of inhalation of smoke, and these interindividual differences could have affected our measurements. (d) The detection limit of the plasma nicotine measurements was not enough to evaluate all blood samples. Thus, we were not able to correlate the plasma nicotine/cotinine levels with Vt. (e) We have not coregistered SPECT images with MRI, which would have been the most appropriate method for placement of ROIs. (f) We have not evaluated the smoker's behavior during the smoking-cessation period. We believe that the nicotine binding and desensitization of the nAChRs in the brain alleviate the cigarette craving and that craving will be the worst during the first 10 d of cessation due to the upregulation of nAChRs. The craving process should minimize after 21 d, as we observe similar levels of occupancy as nonsmokers at that time. However, we cannot exclude the possibility of other nAChR subtypes being involved in the process of tobacco dependence.
CONCLUSION
We have described the in vivo imaging of nAChR upregulation and recovery in response to short-term smoking cessation in smokers using 123I-5IA SPECT. Our results clearly suggest that tobacco smoking is associated with an upregulation of nicotine binding sites in the brain. The upregulation of the nAChRs of the smokers after chronic exposure to nicotine was downregulated to the level in nonsmokers after ∼21 d of smoking cessation. Thus, the upregulation of receptor numbers is a temporary effect. Nicotine dependence and difficulty in smoking cessation are also interesting with regard to the findings of the 123I-5IA SPECT study. The decrease in nicotinic receptors to nonsmoker levels may be the breaking point during the nicotine withdrawal period.
Acknowledgments
The authors thank Dr. Masanori Ichise (Department of Radiology, Columbia University, College of Physicians and Surgeons) for his tremendous editorial support and suggestions on this project. The authors also thank Nihon Medi-Physics Co. Ltd., Japan, for providing sodium 123I-iodide. This work was supported in part by a grant from the Research for the Future Program of the Japan Society for the Promotion of Science (JSPS-RFTF97K00201); Grants-in-Aid for Scientific Research from the Ministry of Education, Science and Technology of Japan; a research grant from Longevity Sciences from the Ministry of Health and Welfare; and a grant from the Smoking Research Foundation.
Footnotes
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication May 9, 2007.
- Accepted for publication July 30, 2007.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Overexpression of Cancer- and Neurotransmitter-Associated Genes in the Nucleus Accumbens of Smokers
- {beta}4-Nicotinic Receptors Are Critically Involved in Reward-Related Behaviors and Self-Regulation of Nicotine Reinforcement
- Chronic Nicotine Exposure Alters the Neurophysiology of Habenulo-Interpeduncular Circuitry
- Change in mental health after smoking cessation: systematic review and meta-analysis
- Nicotine exploits a COPI-mediated process for chaperone-mediated up-regulation of its receptors
- COPI polices nicotine-mediated up-regulation of nicotinic receptors
- Greater Nicotinic Acetylcholine Receptor Density in Smokers Than in Nonsmokers: A PET Study with 2-18F-FA-85380