


Abstract
Various noninvasive imaging methods have been developed to evaluate atherosclerotic plaques. Among them, 18F-FDG PET and MR imaging with ultrasmall superparamagnetic iron oxide particles (USPIO) have been used to quantify plaque inflammation. Both methods are based on the efficient uptake of FDG and USPIO by macrophages in atherosclerotic lesions. Differently polarized macrophages have been reported to have different characteristics that are involved in the pathologic development of atherosclerosis. M1 polarized macrophages are considered the more proatherogenic phenotype than M2 polarized macrophages. However, little is known regarding the association between macrophage polarization and FDG or USPIO accumulation. In this study, we investigated intracellular FDG and USPIO accumulation in M1 and M2 polarized macrophages. Methods: THP-1 macrophages were differentiated into M1 and M2 polarized macrophages. Under optimal glucose conditions, we investigated the 3H-labeled FDG uptake in M1 and M2 polarized macrophages. We then investigated intracellular USPIO uptake by M1 and M2 macrophages. Results: We found that M1 polarization, compared with M2 polarization, results in increased intracellular accumulation of FDG. To elucidate the mechanism by which FDG was preferentially accumulated in M1 macrophages, we examined messenger RNA expressions of glucose transporters (GLUTs) and hexokinases, which have pivotal roles in glucose uptake, and glucose-6-phosphatase (G6Pase), which catalyzes the reverse reaction of hexokinase. In M1 macrophages, GLUT-1, GLUT-3, hexokinase 1, and hexokinase 2 were upregulated and G6Pase was downregulated. In contrast to FDG, M1 polarization resulted in decreased intracellular accumulation of USPIO. We found that scavenger receptor A and CD11b, which are involved in USPIO binding and uptake, were significantly downregulated by M1 polarization. Conclusion: Compared with M2, proatherogenic M1 macrophages preferentially accumulated FDG but not USPIO, suggesting that FDG PET is a useful method for the detection of proinflammatory M1 macrophages.
The rupture of an atherosclerotic plaque heralds many cardiovascular complications such as myocardial and cerebral infarctions. Macrophage infiltration plays an important role in the development of vulnerable plaques. 18F-FDG PET has been reported to be a promising tool for identifying vulnerable atherosclerotic plaques, because detection of these plaques depends on the extent of macrophage infiltration into the atherosclerotic lesions (1–3). To date, some clinical trials have been conducted to examine the usefulness of 18F-FDG PET for evaluating atherosclerosis (1,4). Furthermore, MR imaging with ultrasmall superparamagnetic iron oxide particles (USPIO)—which are also efficiently taken up by macrophages in the atherosclerotic lesion (5,6)—is used to quantify atherosclerotic plaques (6). Recently, macrophages have been considered as heterogeneous cells, which can be differently polarized in atherosclerotic plaques (7–9). Classically activated M1 macrophages promote the destabilization of atherosclerotic plaques, whereas alternatively activated M2 macrophages stimulate reparative processes, leading to stabilization of atherosclerotic plaques (10). Therefore, effective detection of M1 macrophages may help to predict cardiovascular events with greater accuracy. However, the type of macrophages that can be detected by 18F-FDG PET and USPIO MR imaging and the effect of macrophage polarization on FDG and USPIO accumulation are still unclear.
We report here the relationship between the macrophage polarization and imaging probe uptake to the cells. We first investigated the association between macrophage polarization and FDG and USPIO accumulation using human THP-1 macrophages. Then, to elucidate the molecular mechanism of FDG and USPIO accumulation in polarized macrophages, we analyzed the expression of genes responsible for 18F-FDG uptake and metabolism and USPIO uptake and iron export.
MATERIALS AND METHODS
Cell Culture and Chemicals
Human THP-1 cells were obtained from Dainippon Sumitomo Pharma. Cell culture medium was purchased from Life Technologies. Fetal bovine serum was obtained from Thermo Fisher Scientific. Phorbol 12-myristate 13-acetate (PMA) was obtained from WAKO Pure Chemicals. Lipopolysaccharides (LPS) from Escherichia coli O111:B4 was obtained from Sigma-Aldrich. Recombinant human interferon-γ (IFNγ), human interleukin (IL)-4, and human IL-13 were obtained from R&D Systems. [5,6-3H]-2-fluoro-2-deoxy-d-glucose (3H-FDG) was obtained from American Radiolabeled Chemicals. USPIO coated with alkali-treated dextran (average particle size, 28 nm) was custom-synthesized by Meito Sangyo.
Differentiation of THP-1 Cells into Macrophages and Polarization
THP-1 cells were cultured in RPMI1640 supplemented with 10% heat-inactivated fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 μg/mL). THP-1 cells (seeded at 2 × 105 cells/cm2) were differentiated into macrophages using 320 nM PMA and polarized according to the method of Tjiu et al., with some modifications (11). For M1 polarization, cells were treated with PMA for 6 h and then cultured with PMA plus LPS (10 ng/mL) and IFNγ (20 ng/mL) for another 42 h. For M2 polarization, cells were treated with PMA for 6 h and then cultured with PMA plus IL-4 (20 ng/mL) and IL-13 (20 ng/mL) for another 42 h. We also developed control macrophages, which received no stimuli, that were differentiated from THP-1 cells by incubation with PMA for 48 h. To evaluate the appropriate induction of polarization, we measured an M1 marker gene, inducible nitric oxide synthase (iNOS), and an M2 marker gene, mannose receptor C type 1 (MRC1).
3H-FDG Study
Polarized macrophages were rinsed and then precultured in RPMI1640 containing 1 mM glucose at 37°C for 1 h. After the medium was replaced with fresh medium containing 1 mM glucose and 3H-FDG (3.7 kBq/well), cells were cultured for another 3 h. Cells were washed twice with prechilled saline containing 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 1 mM CaCl2, and 1 mM MgCl2 and dissolved with a 0.1% sodium dodecyl sulfate/0.1 M NaOH aqueous solution. The radioactivities of culture medium and cell lysates were measured using a liquid scintillation counter (Accu FLEX LSC 7400; Hitachi Aloka Medical, Ltd.). The cellular protein concentration was measured using the DC Protein Assay Kit (Bio-Rad). 3H-FDG accumulation was expressed as radioactivities in cell lysate per milligram of cell protein.
USPIO Study
Macrophages were incubated with USPIO (400 μg Fe/mL) for the last 16 h of the 2-d treatment with polarizing stimulus at 37°C in 5% CO2. The cells were washed twice with prechilled saline containing 25 mM HEPES, 1 mM CaCl2, and 1 mM MgCl2 and were dissolved with 0.5% Triton X-100 (MP Biomedicals) aqueous solution. The iron contents of cell lysates were measured using the Iron Assay Kit (BioChain). Cellular protein concentration was measured using the DC Protein Assay Kit. The USPIO accumulation was expressed as cellular iron content per milligram of cell protein.
Quantitative Real-Time Polymerase Chain Reaction
Cellular total RNA was extracted using the RNeasy mini kit (Qiagen) and converted into complementary DNA using the High Capacity cDNA Reverse Transcription Kit (Life Technologies). Gene expression was analyzed using the 7900HT Fast Real-Time PCR System (Life Technologies) with the TaqMan Universal Master Mix II (Life Technologies) and primer-probe sets of TaqMan Gene Expression Assays (Life Technologies) for the following genes: iNOS (Hs00167248_m1), MRC1 (Hs00267207_m1), scavenger receptor A (SR-A; Hs00234012_m1), glucose transporter (GLUT)-1 Hs00892681_m1), GLUT-3 (Hs00359840_m1), hexokinase1 (HK1; Hs00175976_m1), hexokinase2 (HK2; Hs00606086_m1), glucose-6-phosphatase (G6Pase; Hs00292720_m1), CD11b (Hs00355885_m1), CD36 (Hs00354519_m1), ferroportin 1 (Fpn1; Hs00205888_m1), and ferritin light chain (FTL; Hs00830226_gH). β-actin (Hs99999903_m1) was used as an endogenous control gene. Relative messenger RNA (mRNA) expression was calculated using the ΔΔCt method.
Statistics
Data are presented as mean ± SD. Statistical analyses were performed using SAS software (version 8.2; SAS Institute, Inc.). Either a Tukey parametric multiple-comparison test or a Steel–Dwass nonparametric multiple-comparison test (Bartlett test; P < 0.05) was used for comparisons among the 3 different macrophage types (i.e., M1, M2, and control). P values of less than 0.05 were considered statistically significant.
RESULTS
Macrophage Polarization
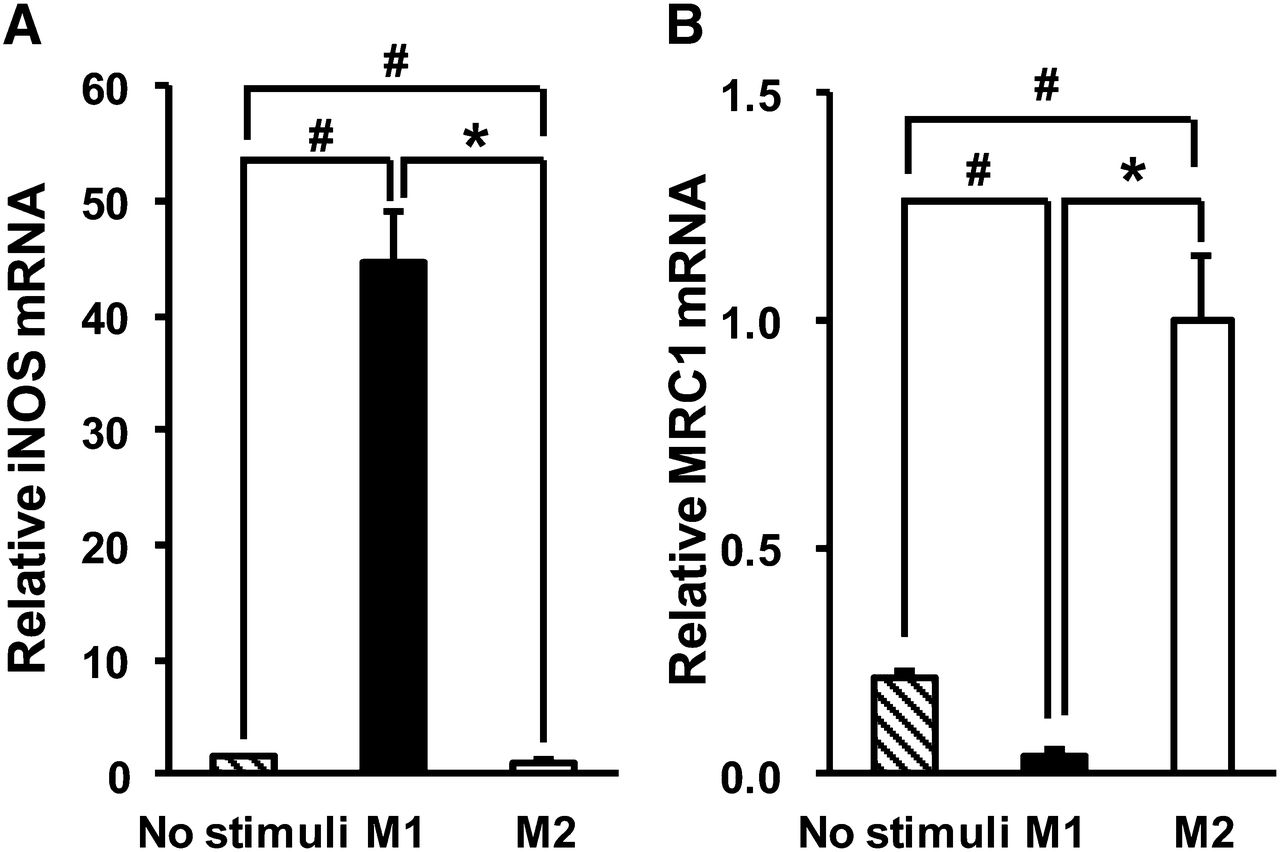
Macrophages treated with IFNγ and LPS showed significant upregulation of iNOS (Fig. 1A) (P < 0.05), whereas those treated with IL-4/IL-13 showed significant upregulation of MRC1 (Fig. 1B) (P < 0.05). The M1 marker genes CCR7 and IL-1β were also significantly upregulated in macrophages treated with IFNγ/LPS, and the M2 marker gene IL-1ra was significantly upregulated in macrophages treated with IL-4/IL-13 (data not shown). Hence, we used these macrophages as for M1 polarization and M2 polarization for the following experiments.
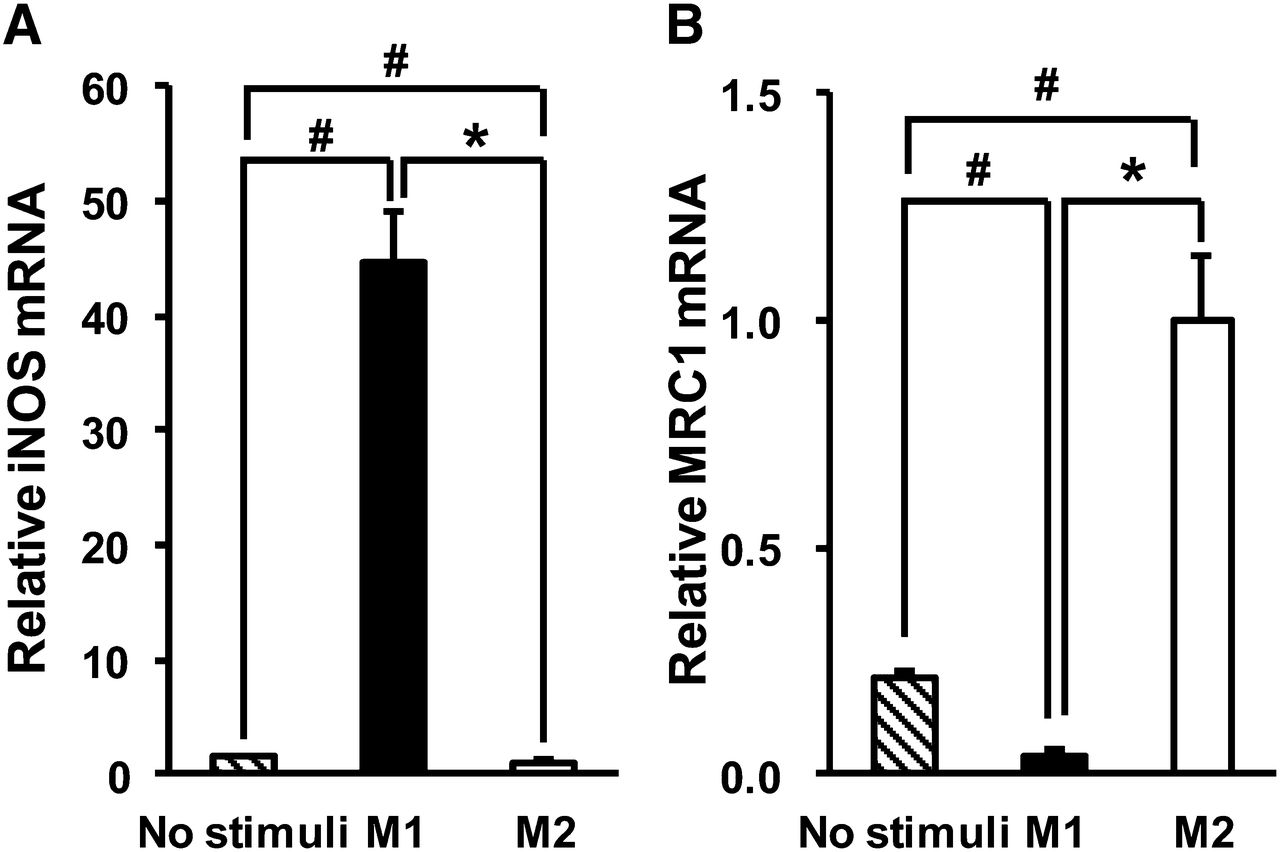
Gene expression of M1 and M2 markers in human THP-1 macrophages treated with each polarizing factor: iNOS as M1 marker (A) and MRC1 as M2 marker (B). Data are presented as mean ± SD (n = 6). *P < 0.05, comparison between M1 and M2. #P < 0.05, compared with no stimuli by Steel–Dwass test.
3H-FDG Accumulation in M1 and M2 Macrophages
To study accumulation of FDG in macrophages, we first optimized the glucose concentration in the medium, because FDG PET signals decrease if blood glucose levels exceed 200 mg/dL. In a preliminary study using a medium containing a high glucose concentration of 200 mg/dL, FDG accumulation in macrophages was markedly reduced (data not shown). In a low-glucose medium of 1.8 mg/dL (∼0.1 mM), we observed that cells were damaged (data not shown). Therefore, we added glucose at 1 mM to the medium—an amount sufficient to detect FDG accumulation without damaging the cells. M1 macrophages, compared with M2 macrophages, showed a 2.6-fold increased uptake of 3H-FDG (Fig. 2) (P < 0.01).

Intracellular 3H-FDG accumulation in human THP-1 macrophages. Data are presented as mean ± SD (n = 6). **P < 0.01, comparison between M1 and M2. ##P < 0.01, compared with no stimuli by Tukey test.
Expression of Glucose Metabolism–Related Genes in Macrophages
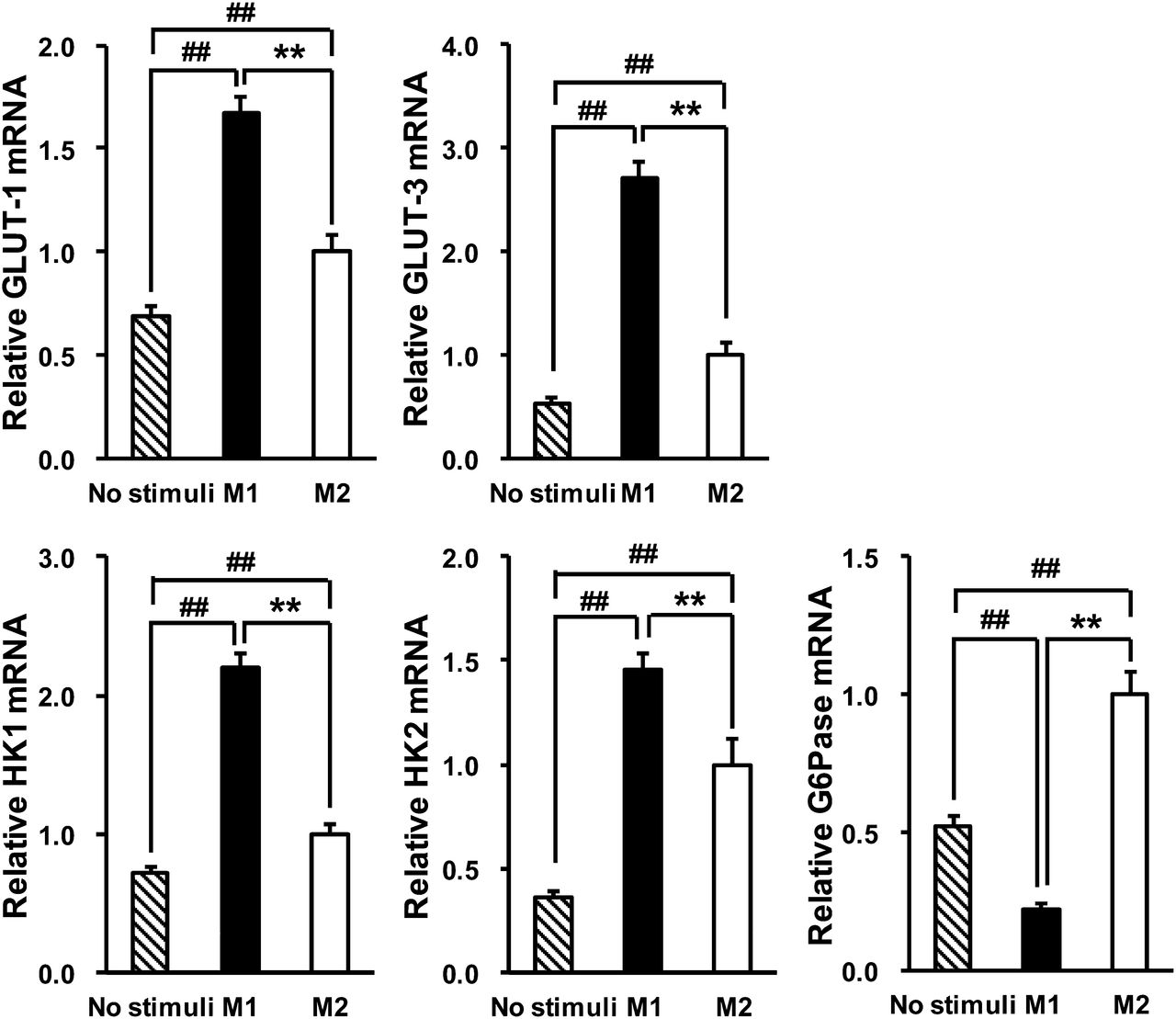
We measured the mRNA expression of glucose metabolism–related genes to elucidate the mechanism by which FDG was preferentially taken up by M1 macrophages. We found that GLUT-1 and GLUT-3, which are major isoforms of a glucose transporter in macrophages (12,13), were significantly upregulated in M1 macrophages, compared with M2 macrophages (Fig. 3) (P < 0.01 and < 0.01, respectively). HK1 and HK2, which catalyze the intracellular phosphorylation of glucose into glucose-6-phosphate, were also significantly upregulated in M1 macrophages, compared with M2 macrophages (Fig. 3) (P < 0.01 and < 0.01, respectively). However, the expression of G6Pase, which is an enzyme that catalyzes the reverse reaction of hexokinases, was significantly downregulated in M1 macrophages, compared with M2 macrophages (Fig. 3) (P < 0.01). These results suggest that the increase of FDG accumulation in M1 macrophages was caused by the upregulation of GLUT and hexokinase genes as well as the downregulation of G6Pase gene.

Expression of glucose metabolism–related genes in human THP-1 macrophages: GLUT-1, GLUT-3, HK1, HK2, and G6Pase. Data are presented as mean ± SD (n = 6). **P < 0.01, comparison between M1 and M2. ##P < 0.01, compared with no stimuli by Tukey test.
USPIO Accumulation in M1 and M2 Macrophages
Previous USPIO accumulation studies have used macrophages treated with USPIO in concentrations ranging from 50 to 500 μg Fe/mL and for time periods lasting from 1 to 72 h (14–17). On the basis of our preliminary study findings, macrophages were incubated with USPIO (400 μg Fe/mL) for the last 16 h of the 2-d treatment with polarizing stimulus at 37°C in 5% CO2. USPIO accumulation in the cells increased linearly with both USPIO concentration and incubation time (data not shown). In contrast to FDG, USPIO accumulation in M2 macrophages was 1.4-fold higher than that in M1 macrophages (Fig. 4) (P < 0.01).
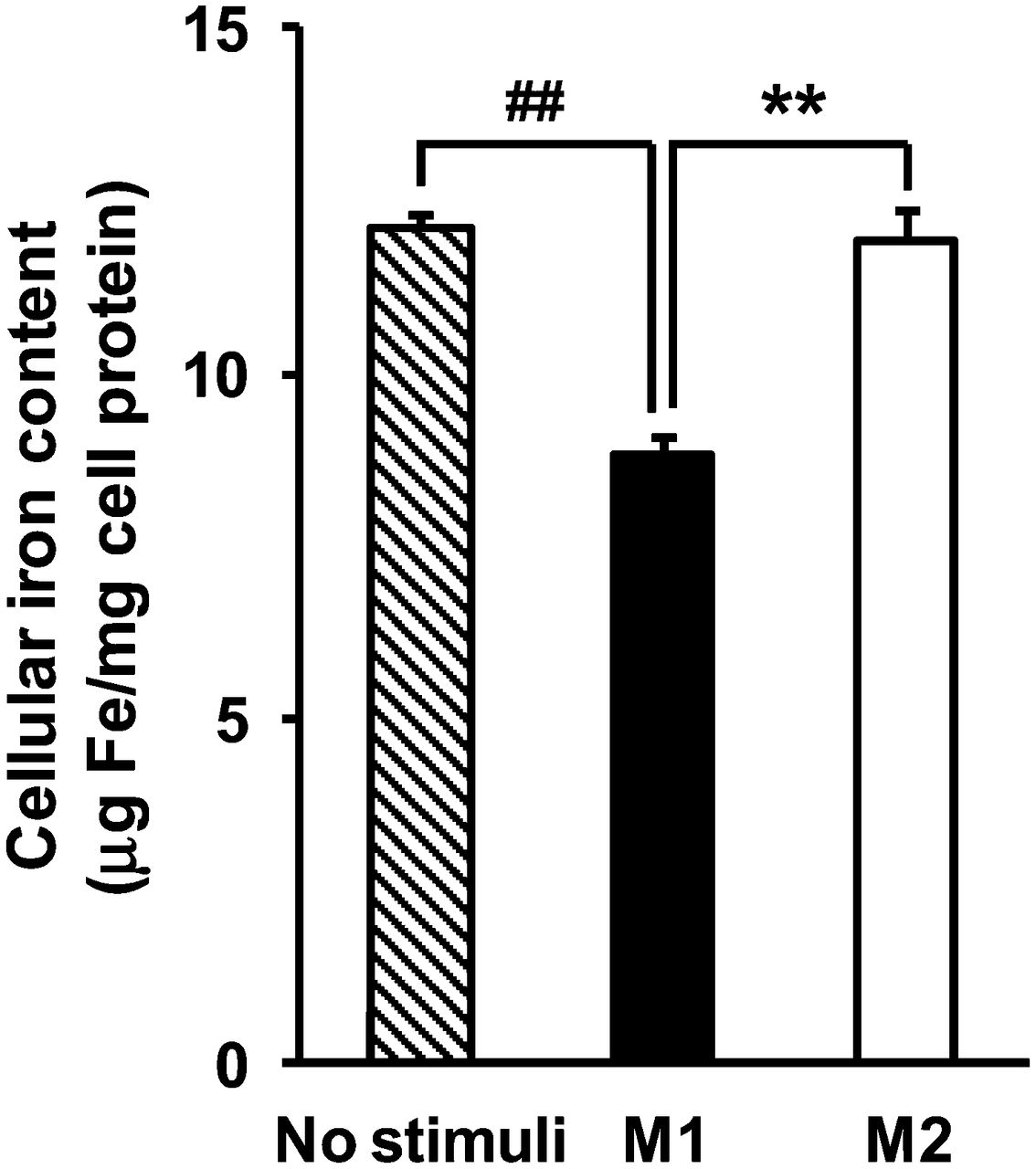
Intracellular USPIO accumulation in human THP-1 macrophages. Data are presented as mean ± SD (n = 6). **P < 0.01, comparison between M1 and M2. ##P < 0.01, compared with no stimuli by Tukey test.
Expression of USPIO Uptake- and Iron Export–Related Genes in Macrophages
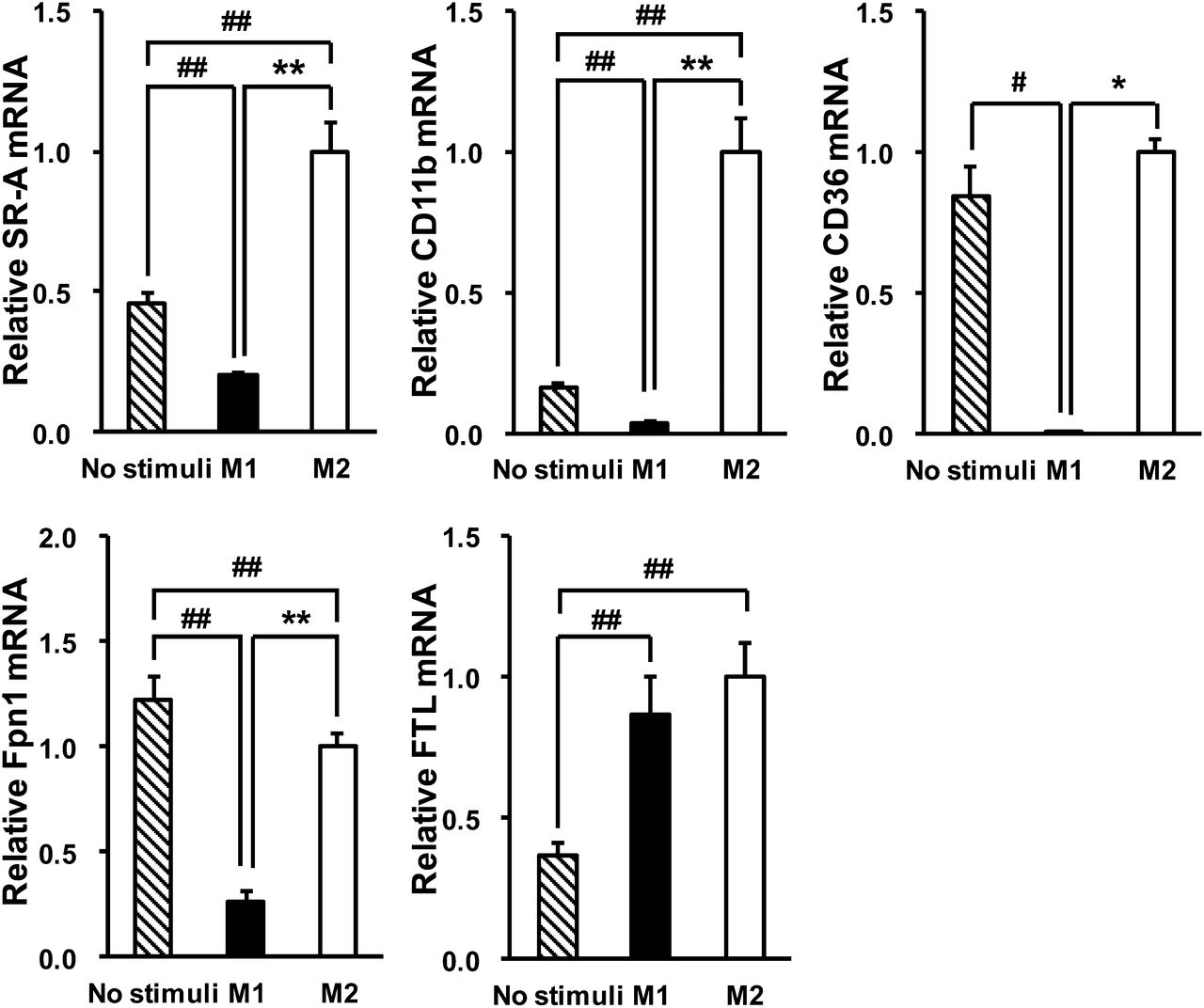
To elucidate the mechanism by which USPIO was preferentially taken up by M2 macrophages, compared with M1 macrophages, we measured the mRNA expression of SR-A and CD11b (14,18). Both SR-A and CD11b were significantly upregulated in M2 macrophages, compared with M1 macrophages (Fig. 5) (P < 0.01 and < 0.01, respectively). Considering that other scavenger receptors may contribute to the uptake of USPIO, we measured the expression of CD36, another major scavenger receptor. As shown in Figure 5, CD36 was also upregulated in M2 macrophages. The expression of Fpn1, the major iron exporter in mammalian cells (19), was also significantly upregulated in M2 macrophages, compared with M1 macrophages (Fig. 5) (P < 0.01). There was no significant difference in the expression of FTL, which is responsible for intracellular iron storage (20), between M1 and M2 macrophages (Fig. 5).
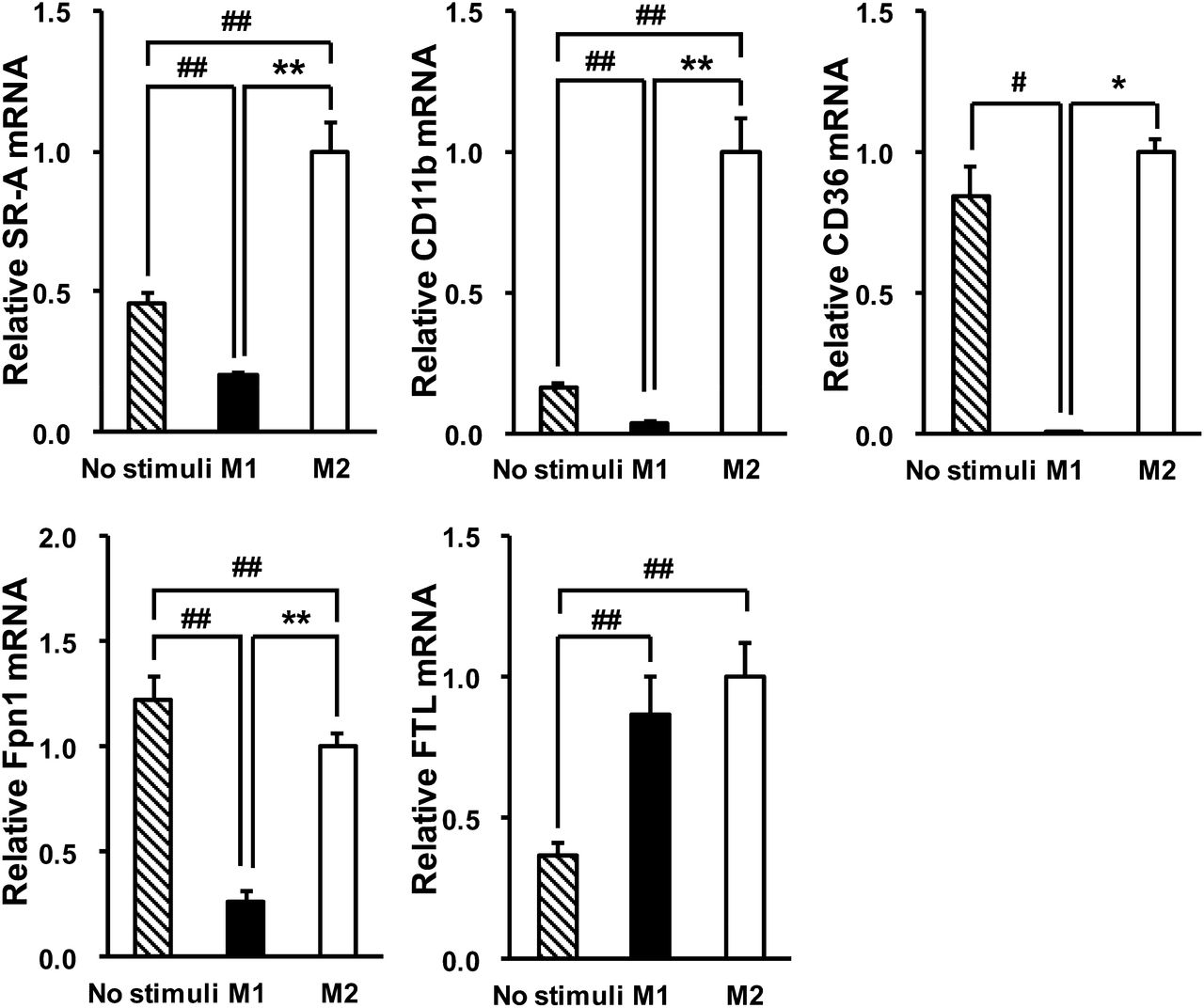
Expression of USPIO uptake- and iron export–related genes in human THP-1 macrophages: SR-A, CD11b, CD36, Fpn1, and FTL. Data are presented as mean ± SD (n = 6). *P < 0.05 and **P < 0.01, comparison between M1 and M2. #P < 0.05 and ##P < 0.01, compared with no stimuli by Tukey test or Steel–Dwass test.
DISCUSSION
18F-FDG PET and USPIO MR imaging have been clinically used to quantify atherosclerotic plaques, because both FDG and USPIO are efficiently taken up by macrophages in atherosclerotic lesions. Recently, macrophages in varying polarized states have been reported to have different characteristics that are involved in the pathologic development of atherosclerosis (7,10). In this study, we compared the ability of differently polarized macrophages to take up FDG and USPIO using human THP-1 macrophages. We showed that FDG was taken up preferentially by M1 macrophages, whereas USPIO was taken up by M2 macrophages.
The preferential uptake of FDG by M1 macrophages led to the hypothesis that M1 and M2 differed in the expression of the glucose uptake–related molecules because FDG is taken up by cells in a manner analogous to glucose. We observed that the mRNA expression of GLUT-1 and GLUT-3 was upregulated in M1 macrophages. Kim et al. also reported that the mRNA expression of GLUT-1 was upregulated and intracellular FDG accumulation increased in mouse RAW264.7 macrophages stimulated by LPS (21). In addition, we showed that the mRNA expression of HK1 and HK2, which are major hexokinase isoforms, was upregulated, whereas the mRNA expression of G6Pase was downregulated in M1 macrophages. FDG is taken up into cells via glucose transporters and then phosphorylated by hexokinases to FDG-6-phosphate (FDG6P) in a mechanism similar to glucose. FDG6P is retained inside the cell, such that it can neither follow the process of glycolysis nor easily pass through the plasma membrane. If G6Pase, the catalyst of the reverse reaction of hexokinase, converts FDG6P into FDG, FDG can theoretically diffuse out of cells via glucose transporters. The present study suggests that the regulation of gene expression of key molecules involved in glucose uptake contributed to the increase in intracellular FDG accumulation in M1 macrophages. In particular, the increased intracellular FDG uptake via glucose transporters, the phosphorylation of FDG by hexokinases, and the decrease in dephosphorylation of FDG6P by G6Pase contributed to the increase of FDG uptake in M1 macrophages. On the other hand, inconsistent with our observation, Folco et al. reported that inflammatory stimulation did not increase the glucose uptake in human primary monocytes (22). They used IFNγ or a combination of IFNγ, TNFα, and IL1β as inflammatory stimulation, whereas we adopted a classic M1 stimulation for characterizing typical phenotypes of M1 macrophages. We need to consider a more relevant stimulation to atherosclerosis in future experiments.
Small iron oxide particles (SPIO) and USPIO have been used clinically as superparamagnetic MR imaging contrast agents. These iron oxide particles are efficiently taken up by macrophages. A range of molecules is considered to be involved in SPIO and USPIO uptake and iron export. Lunov et al. identified the clathrin-mediated SR-A–dependent endocytosis as a major pathway of USPIO uptake in peripheral blood monocyte–derived macrophages (14). von Zur Muhlen et al. showed that USPIO uptake was inhibited by treatment with anti-CD11b antibody in PMA-stimulated human peripheral blood monocytes (18). In the present study, we determined that both SR-A and CD11b were significantly upregulated in M2 macrophages, which might contribute to the increase in USPIO uptake by M2 macrophages (Fig. 5). Gu et al. reported that internalized USPIO in mouse macrophages was degraded in lysosomes, and free iron was released into intracellular iron pools (23). Fpn1 is known to have an important role in the export of iron from cells (19). We observed that Fpn1 was also upregulated in M2 macrophages, compared with M1 macrophages (Fig. 5), consistent with the previous report (24). In the present study, we observed a statistically significant increase of USPIO uptake in M2 macrophages, compared with M1 macrophages, but the difference was relatively small, despite the marked induction of genes related to the uptake of USPIO. The upregulation of Fpn1 could cause the reduction of the cellular iron levels, which might attenuate the accumulation of USPIO in M2 macrophages. Taken together, the accumulation of cellular USPIO was higher in M2 macrophages than in M1 macrophages, most likely as a result of the balance of transcriptional regulation of genes related to uptake and export of USPIO. In the present study, there were no difference in USPIO accumulation between M2 macrophages and control ones (Fig. 4), suggesting the difficulty of visualizing only M2 macrophages in distinction from nonpolarized macrophages by USPIO. Recently, it has been reported that the status of macrophage polarization in atherosclerotic plaques was dramatically changed in the progression and regression process of atherosclerosis using an experimental murine model (8,25). In addition, a substantial number of macrophages were polarized in those lesions. Therefore, USPIO should be able to light up the mainly M2 lesion, although we need to pay attention to the false-positive results. In contrast to our findings, Rogers et al. reported that not only Th2 cytokine IL-4 but also Th1 cytokine IFNγ increased SPIO uptake in mouse J774A.1 macrophages (26). However, differing experimental conditions including cell type and polarization methods might explain these discrepancies. Rogers et al. used IFNγ and IL-4 alone as Th1 and Th2 cytokines, respectively, whereas we used IFNγ in addition to LPS and IL-4 plus IL-13 as M1-polarizing and M2-polarizing stimuli, respectively. In this study, we investigated the USPIO uptake with a 16-h incubation to obtain the reasonably sufficient amount of intracellular USPIO for quantitative analysis, whereas the incubation time was 3 h for FDG. The exposure time might affect the uptake of FDG and USPIO in M1 and M2 polarized macrophages. Cell viability also should be another important factor to evaluate the cellar uptake. In this study, at least, the cell damage was not observed morphologically. Further investigation about exposure time and cell viability is needed for precise analysis.
The ultimate purpose of atherosclerosis imaging, and the detection of vulnerable rupture-prone plaques in particular, is the accurate prediction of cardiovascular events. Recently, studies have suggested that the quality rather than quantity of macrophages is critical in the evolution of vulnerable plaques (27–29). M1 macrophages play a key role in the development of atherosclerosis, which is mediated by production of proinflammatory cytokines or reactive nitrogen oxides, whereas M2 macrophages have antiatherogenic properties mediated by the production of antiinflammatory cytokines and the suppression of proinflammatory signaling (30,31). However, the contribution of these differently polarized macrophages to vulnerable plaque formation, potentially resulting in plaque rupture, has not yet been fully understood. For example, Mauriello et al. recently reported that M2 but not M1 macrophages were present in the fibrous cap near the rupture site in the human carotid artery, suggesting that M2 macrophages might also modulate the process of plaque rupture (32). Although further investigations regarding the roles of M1 and M2 macrophages in atherosclerotic lesions are needed, it is clear that macrophage polarization affects plaque diagnosis and prognosis. In our preliminary in vivo study, stronger signals were observed in abdominal plaques than in plaques in the aortic arch in en face autoradiography of the whole aorta of 14C-FDG–injected apolipoprotein E knockout mice (Supplemental Fig. 1; supplemental materials are available online only at http://jnm.snmjournals.org). The M1 markers iNOS, IL-1β, and CCR7 were upregulated in the abdominal plaques, whereas the M2 marker MRC1 was upregulated in the aortic arch plaques (Supplemental Fig. 2). These results suggest that 18F-FDG PET should dominantly visualize the M1 macrophage–infiltrated area. Some reports have suggested that 18F-FDG PET may be useful to monitor the therapeutic effects of drugs (33–35), and some of the drugs are reported to affect the polarization of macrophages (8,36). Because therapies that reduce M1 macrophages are garnering more attention, it becomes increasingly important to select appropriate imaging methods that accurately detect plaque development.
CONCLUSION
The present study provides in vitro evidence that 18F-FDG PET preferentially detects proinflammatory M1 macrophages, whereas USPIO MR imaging preferentially detects antiinflammatory M2 macrophages. Although further in vivo investigations are needed, 18F-FDG PET imaging should be useful in the detection of proinflammatory M1 macrophages.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. No potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Drs. Masaaki Mori, Masakuni Noda, Shota Ikeda, Richard Urquhart, Ryogo Hayashi, Hiroki Nagase, Aya Tanaka, Takahisa Matsuda, and Yukio Yamada in our laboratories for their invaluable suggestions and feedback.
Footnotes
Published online May 13, 2013.
- © 2013 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication June 28, 2012.
- Accepted for publication December 17, 2012.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Diagnostic Applications of Ultrasmall Superparamagnetic Particles of Iron Oxide for Imaging Myocardial and Vascular Inflammation
- Multimodality Imaging of Inflammation and Ventricular Remodeling in Pressure-Overload Heart Failure
- Molecular Imaging of Myocardial Inflammation With Positron Emission Tomography Post-Ischemia: A Determinant of Subsequent Remodeling or Recovery
- Molecular Imaging of Vulnerable Coronary Plaque: A Pathophysiologic Perspective
- GM-CSF Enhances Macrophage Glycolytic Activity In Vitro and Improves Detection of Inflammation In Vivo
- Imaging Atherosclerosis
- Systemic Atherosclerotic Inflammation Following Acute Myocardial Infarction: Myocardial Infarction Begets Myocardial Infarction
- Can the Inflammatory Response Be Evaluated Using 18F-FDG Within Zones of Microvascular Obstruction After Myocardial Infarction?
- Splenic Metabolic Activity Predicts Risk of Future Cardiovascular Events: Demonstration of a Cardiosplenic Axis in Humans
- 18F-Fluorodeoxyglucose Positron Emission Tomography/Computed Tomography Enables the Detection of Recurrent Same-Site Deep Vein Thrombosis by Illuminating Recently Formed, Neutrophil-Rich Thrombus
- Pharmacological Activation of the Melanocortin System Limits Plaque Inflammation and Ameliorates Vascular Dysfunction in Atherosclerotic Mice
- Clinical and research applications of simultaneous positron emission tomography and MRI
- Measurement of Arterial Activity on Routine FDG PET/CT Images Improves Prediction of Risk of Future CV Events
- Magnetic Resonance Imaging Investigation of Macrophages in Acute Cardiac Allograft Rejection After Heart Transplantation