


Abstract
Human prostate cancers (PC) overexpress gastrin-releasing peptide (GRP) receptors. This observation suggests that GRP receptors may be used as new visualization and treatment modalities for these tumors. Radiolabeled GRP receptor-targeting analogs of GRP and bombesin (BN) have successfully been developed for these purposes. Expression of GRP receptors in human prostate tumors is, however, primarily evaluated in early stages of tumor development and information on expression in the more progressive prostate tumors is uncertain. To evaluate GRP receptor expression in all stages of PC, we investigated GRP receptor expression using a panel of 12 established human PC xenograft models representing the different stages of human PC and the effect of antiandrogen treatment (castration). Methods: Human PC xenografts were grown in male nude mice, and GRP receptor density in the tumors was evaluated using displacement receptor autoradiography with the universal BN receptor analog 125I-[d-Tyr6,β-Ala11,Phe13,Nle14]BN(6–14) and the BN analog 111In-[DTPA-Pro1,Tyr4]BN (DTPA is diethylenetriaminepentaacetic acid) before and after castration. Results: Autoradiography showed high-density GRP receptor expression in the androgen-dependent tumors (3/12 models), whereas only very low receptor expression was found in the androgen-responsive and -independent tumors (9/12 models). Castration resulted in GRP receptor downregulation (11%–36% of initial values) in the 3 androgen-dependent tumors. Conclusion: High GRP receptor density was only observed in androgen–dependent PC xenografts, indicating high GRP receptor expression in the early, androgen-dependent, stages of prostate tumor development and not in later stages. In addition, castration strongly reduced GRP receptor expression in androgen-dependent tumors, indicating that GRP receptor expression in human PC is androgen-regulated.
Prostate cancer (PC) is the third leading cause of cancer-related deaths and the most frequently diagnosed cancer in men in Western countries (1). Early screening for PC has resulted in a sharp increase in the number of locally confined tumors, although the micrometastatic stage of these tumors is often uncertain. Lymph node metastases have a significant impact on the prognosis of PC patients as it dictates the available treatment options: Locally confined tumors can be treated by radical prostatectomy or radiation therapy, whereas tumors that are no longer confined to the prostate require androgen ablation treatment by surgical or chemical castration or treatment with antiandrogens. Most PC patients are initially androgen sensitive and will respond to androgen ablation therapy. However, a major problem in the treatment of metastasized PC is the transition of this androgen-responsive characteristic toward hormone-independent growth. This shift from androgen dependence to androgen-independent tumor growth is not well understood (2,3), and effective treatment of androgen-independent PC is not available (4). Consequently, accurate staging of early-diagnosed PC is a crucial step in the management of PC. Besides the need for accurate staging at presentation, a sensitive diagnostic technique is urgently needed to detect local relapse or metastatic disease of PC patients who have undergone local therapy but are faced with rising prostate-specific antigen (PSA). In case of a local recurrence after radical prostatectomy additional local radiotherapy can be given. This therapy is often initiated on the basis of PSA kinetics, without any direct evidence for a local recurrence because it is very difficult to demonstrate. Clearly, there is a clinical need to find a target present in early stages of PC as well as in metastases that may be used for molecular imaging.
It has been shown that prostate tumors overexpress gastrin-releasing peptide (GRP) receptors while normal and hyperplastic prostate tissue was receptor-negative (5,6). This GRP receptor expression is predominantly evaluated on primary human prostate tumor tissue from radical prostatectomy specimens. In these cases, the state of androgen dependence has not been published. Among the 7 androgen-independent bone metastases tested, 4 cases were GRP receptor-positive with very diverse receptor densities (6). Nevertheless, the GRP receptor expression in later stages of PC is unclear, and, therefore, the value of GRP receptor-targeted therapy in androgen-independent PC remains uncertain.
The GRP receptor is a member of the bombesin (BN) receptor family, which consists of 4 known receptor subtypes. Three of them, the neuromedin B (NMB) receptor (BB1), the GRP receptor (BB2), and the BN receptor subtype 3 (BRS-3 or BB3), are mammalian receptors, whereas the fourth subtype (BB4) is found only in amphibians (7–10). Except for the GRP receptor, these receptor subtypes are not very well characterized with regard to their distribution and function in human tissues (11,12).
BN-like peptides, including BN, GRP, and NMB, are involved in the regulation of a large number of biologic processes in the gut and central nervous system (13) and mediate their action by binding to BN receptors. It has been proposed that radiolabeled analogs of these BN-like peptides may be used to image and treat GRP receptor–expressing tumors, such as prostate tumors (14,15). This peptide-based receptor-targeted imaging and treatment of tumors have been demonstrated successfully for somatostatin receptor–expressing tumors using radiolabeled somatostatin analogs (16–18).
A number of researchers are working on the development of radiolabeled BN analogs that specifically target GRP receptor–expressing tumors in vitro and in vivo (15,19–22). Most of these research groups use human GRP receptor–expressing prostate tumor models such as PC-3 and DU-145. These cell lines grow independent of androgen and do not express androgen receptor or secrete PSA, which in itself is an uncharacteristic feature of most PC patients (23). Therefore, it is questionable whether these cell lines can be considered representative models for clinical PC. To determine the value of newly developed BN analogs in the various stages of prostate tumor development, more realistic prostate tumor models would add significant value.
The availability of experimental human prostate tumor models is rather limited. A unique panel of human prostate tumor xenograft models has been established and reviewed by Van Weerden et al. (24,25). This panel comprises 12 xenograft models from 9 different patients. The xenografts represent the various aspects of human PC and, therefore, are excellent tools to study PC progression from androgen-dependent to androgen-independent stages. Three models (PC-295, PC-310, PC-82) are strictly dependent on androgens for their development and growth: These tumors do not develop in castrated male mice, and castration of tumor-bearing animals results in regression without tumor relapse (26,27). Two models, PC-346 (including the sublines PC-346 P, PC-346 I, PC-346 B, and PC-346 iB) and PC-374, are androgen-responsive but not androgen-dependent: Tumors do develop and continue to grow in castrated male mice, but at a lower growth rate. Finally 4 xenografts (PC-133, PC-135, PC-324, and PC-339) are representatives of androgen-independent PC. These tumors develop identically in intact or castrated male mice, and their growth rate does not change on testosterone depletion or supplementation.
In this study we evaluated the expression of GRP receptors and its androgen regulation throughout the various stages of prostate tumor development using the panel of 12 human prostate tumor xenograft models. In vitro BN receptor– binding studies were performed on frozen xenograft sections using radiolabeled BN analogs.
MATERIALS AND METHODS
Prostate Cancer Xenografts
The panel of human PC xenografts is listed in Table 1. Xenografts were routinely propagated in intact male nude NMRI mice (Taconic M&B) (24,25). Mice were kept in 14 × 13 × 33.2 cm3 individually ventilated cages (Techniplast) with 3 or 4 mice per cage, on sawdust (Woody-Clean, type BK8/15; Technilab-BMI) on a 12-h light–dark cycle, at 50% relative humidity, in a temperature-controlled (22°C) room. Mice received irradiated chow and acidified drinking water ad libitum. The experiment was approved by the Animal Experimental Committee (DEC) of the Erasmus University and performed in agreement with The Netherlands Experiments on Animals Act (1977) and the European Convention for Protection of Vertebrate Animals used for Experimental Purposes (Strasbourg; March 18, 1986).
PC Xenograft Panel
Mice were implanted subcutaneously with small fragments of human prostate tumor. In the case of androgen-dependent tumor transplantation, mice were supplemented with testosterone to obtain optimal tumor take (80%–85%) and tumor growth. Hormonal substitution was achieved by implanting Silastic tubings filled with crystalline steroid (6 mg/tubing; AppliChem). These testosterone implants lead to testosterone levels exceeding the low physiologic levels in intact male nude mice for at least 75 d (28).
Androgen withdrawal was performed by surgical castration under ketamine (Alfasan) and Rompun (Bayer AG) anesthesia (1:1 mixture), which was followed by removal of the Silastic testosterone implants. Mice were sacrificed 8–10 d after castration. Tumors were removed and snap frozen in liquid nitrogen for in vitro autoradiography.
Radiolabeled Peptides
BN analog [d-Tyr6,β-Ala11,Phe13,Nle14]BN(6–14), referred to as universal ligand, was provided by Biosynthema Inc., and radiolabeled with 125I as described previously (29).
Analog [DTPA-Pro1,Tyr4]BN (DTPA is diethylenetriaminepentaacetic acid) was provided by Mallinckrodt Inc. and radiolabeled with 111In (111InCl3 [DRN 4901], Tyco Healthcare; 370 MBq/mL in HCl, pH 1.5–1.9) as described earlier (30).
In Vitro Autoradiography
In vitro autoradiography with the radiolabeled BN receptor–binding 125I-universal ligand was performed on PC xenografts sections to detect the expression of all members of the BN receptor family. Displacement with GRP (only GRP receptor binding), NMB (only NMB receptor binding), or BN (mainly GRP receptor and some NMB receptor binding) was used to discriminate between binding to the different receptor subtypes. This is an established method to detect BN receptor subtype expression on several human tissues, as described by Reubi et al. (5).
Frozen PC xenograft sections (10 μm) were incubated for 1 h at room temperature with 0.1 nmol/L 125I-universal ligand (125I-[d-Tyr6,β-Ala11,Phe13,Nle14]BN(6–14)) (74 TBq/mmol) in 167 mmol/L Tris (pH 7.6), supplemented with 5 mmol/L MgCl2, 1% bovine serum albumin (BSA), and 40 μg/mL bacitracin. To differentiate between binding of the universal ligand to the different members of the BN receptor family, the sections were incubated in the presence of excess amounts (1 μmol/L) of either universal ligand, GRP, NMB, or BN (Sigma-Aldrich). After incubation, the sections were washed 2 times for 5 min each in 167 mmol/L Tris (pH 7.6), 5 mmol/L MgCl2, and 0.25% BSA (4°C), 5 min in 167 mmol/L Tris (pH 7.6) and 5 mmol/L MgCl2 (4°C), and finally rinsed in MilliQ water (Millipore) (4°C). The sections were then dried and exposed to phosphor imaging screens (Perkin Elmer) for 72 h. The imaging screens were read using a Cyclone Storage Phosphor System (Packard), and the autoradiograms were quantified using Optiquant Software (Packard).
In vitro receptor binding of 111In-[DTPA-Pro1,Tyr4]BN (200 MBq/nmol) was determined on PC xenograft sections as described. Sections were incubated for 1 h with 1 nmol/L of the 111In-labeled peptide with or without 1 μmol/L [Tyr4]BN (Sigma-Aldrich). The imaging screens were read after an overnight exposure of the xenograft sections.
RESULTS
BN receptor expression was determined on the 12 human prostate tumor xenografts using 125I-universal ligand and 111In-[DTPA-Pro1,Tyr4]BN. The results of the in vitro autoradiography studies are shown in Figures 1 and 2, respectively.
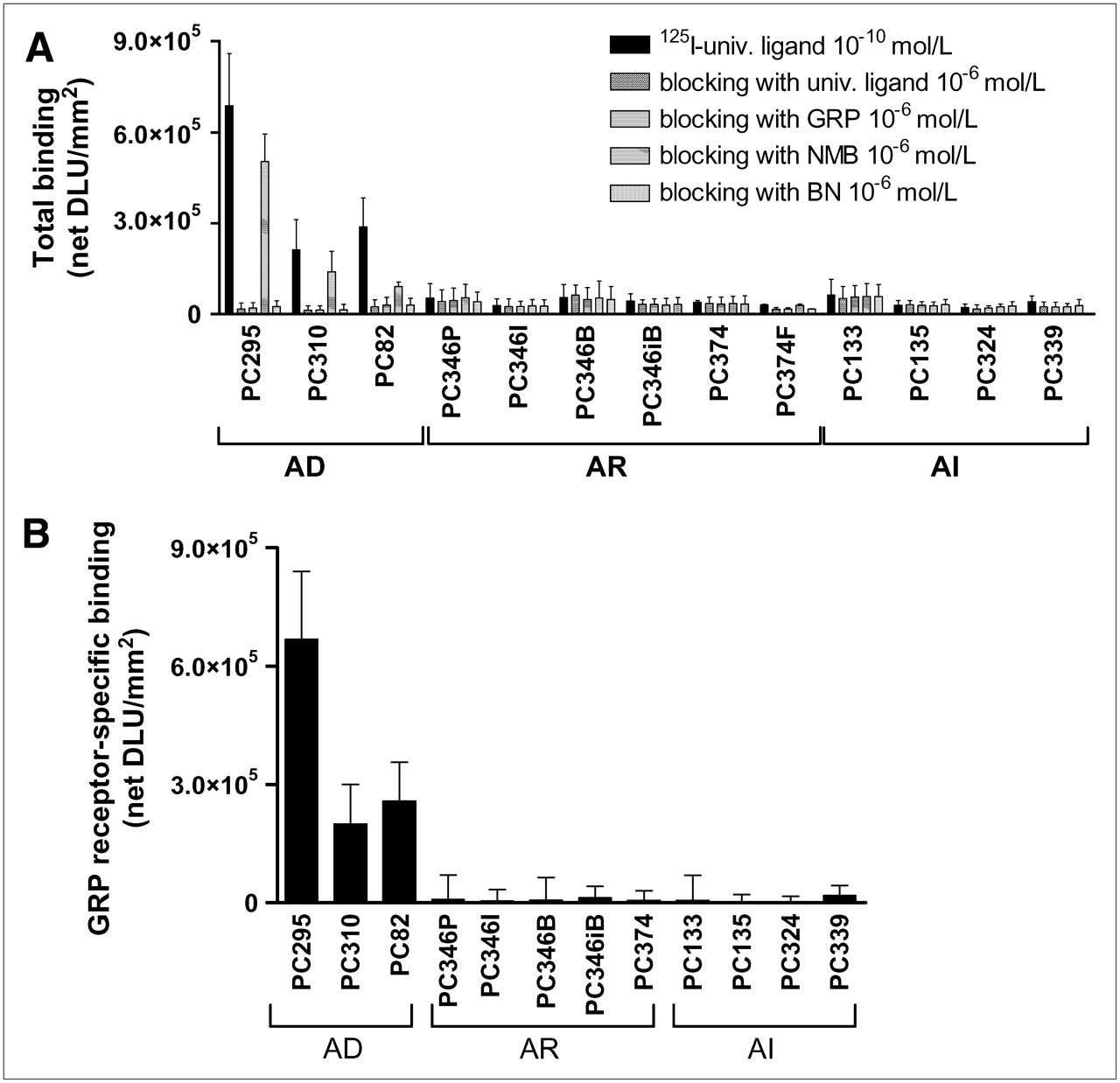
Total binding of 10−10 mol/L 125I-universal (univ.) ligand with or without a blocking concentration (10−6 mol/L) of unlabeled universal ligand, GRP, NMB, or BN (A) and GRP receptor–specific binding (= total binding [125I-universal ligand 10−10 mol/L] minus unspecific binding [blocking with GRP 10−6 mol/L]) (B) to androgen-dependent (AD), androgen-independent but responsive (AR), and androgen-independent (AI) PC xenograft sections. Results are shown as average net density light units per square millimeter (net DLU/mm2) ± SD. Results are average of 3 independent experiments (n = 2–4 sections/experiment).

Total binding of 10−9 mol/L 111In-[DTPA-Pro1,Tyr4]BN with or without a blocking concentration (10−6 mol/L) of unlabeled [Tyr4]BN, to androgen-dependent (AD), androgen-independent but responsive (AR), and androgen-independent (AI) PC xenograft sections. Results are shown as average net density light units per square millimeter (net DLU/mm2). Results are average of 1 experiment (n = 2 sections/experiment).
The androgen-dependent PC xenografts PC-295, PC-310, and PC-82 showed high binding of 125I-universal ligand, which could be almost completely blocked by 10−6 mol/L of universal ligand, GRP, and BN and to a much lesser extent by 10−6 mol/L NMB (Fig. 1A). This blocking pattern indicates high receptor-mediated binding of 125I-universal ligand to predominantly GRP receptors (Fig. 1B). The androgen-independent, but responsive and the completely androgen-independent, PC xenografts showed very low receptor-mediated binding of 125I-universal ligand (Figs. 1A and 1B). These results suggest that these xenografts express GRP receptors only at very low density. The other members of the BN receptor family, NMB and BB3, are not, or to a much lesser degree, expressed in this panel of PC xenografts.
Similar results were found in the 111In-[DTPA-Pro1,Tyr4]BN in vitro autoradiography: high receptor-mediated binding in androgen-dependent PC xenografts and only very low binding in androgen-responsive and androgen-independent PC-xenografts (Fig. 2).
To determine hormonal regulation of the BN receptor family members, PC xenograft–bearing mice were androgen ablated by castration and implant removal. The effect of this treatment on the expression of BN receptors was determined 8–10 d after castration using 125I-universal ligand and 111In-[DTPA-Pro1,Tyr4]BN in an in vitro autoradiographic study. The GRP receptor–mediated binding in the androgen-dependent PC xenografts was drastically reduced after castration (Figs. 3A and 3B). The androgen-independent but responsive and the androgen-independent PC xenografts showed no alteration in their binding profile of the radiolabeled peptides after castration of the animals (data not shown).
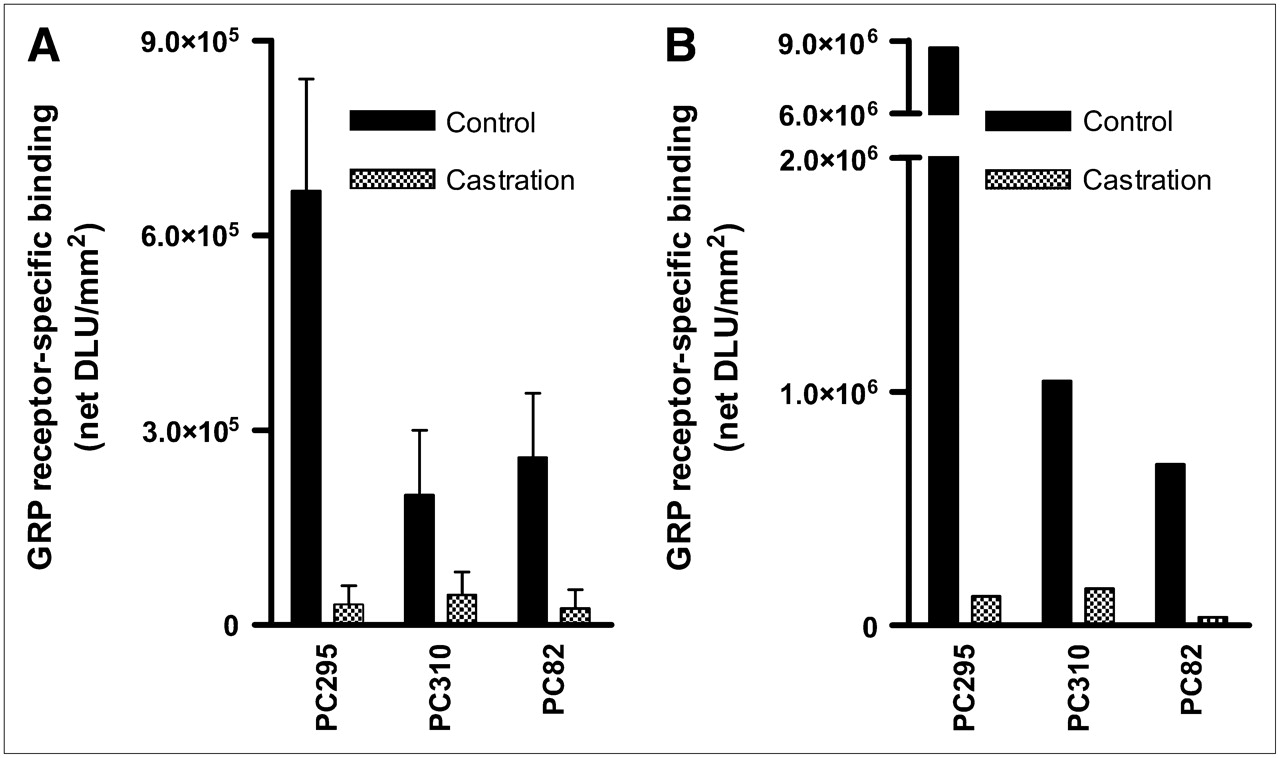
Receptor-specific binding of 125I-universal ligand (= total binding [125I-universal ligand 10−10 mol/L] minus unspecific binding [blocking with GRP 10−6 mol/L]) (A) and 111In-[DTPA-Pro1,Tyr4]BN (= [111In-[DTPA-Pro1,Tyr4]BN 10−9 mol/L] minus [blocking with [Tyr4]BN 10−6 mol/L]) (B) to androgen-dependent (AD), androgen-independent but responsive (AR), and androgen-independent (AI) PC xenograft sections. Xenografts were obtained from control mice or castrated mice (castration 8–10 d before tumor collection). Results are shown as average net density light units square millimeter (net DLU/mm2). Results are average of 3 independent experiments with 125I-universal ligand (1 experiment with 111In-[DTPA-Pro1,Tyr4]BN) (n = 2 sections/experiment).
DISCUSSION
The overexpression of peptide receptors in human tumors is of considerable clinical interest (31). For example, the overexpression of somatostatin receptors on human neuroendocrine tumors has enabled successful receptor-targeted visualization and treatment of these tumors. The long-term octreotide treatment of patients with somatostatin receptor–expressing tumors has been successful in relieving the symptoms related to excessive hormone production by these tumors (32). Also the use of radiolabeled somatostatin analogs has permitted visualization of neuroendocrine tumors and their metastases in patients (16). Additionally, 177Lu- and 90Y-labeled somatostatin analogs have successfully been used in radionuclide therapy in this group of patients (33–35).
BN is a neuropeptide with a high affinity for the GRP receptor. This opens attractive clinical applications using the GRP receptor as a target, such as treatment with BN receptor antagonists, targeted chemotherapy with cytotoxic BN analogs, and peptide-receptor scintigraphy and radionuclide therapy using radiolabeled BN analogs. It has been reported that the GRP receptor is expressed in high densities on PC cells (5,6,36,37). Markwalder and Reubi described high GRP receptor expression in primary PC and prostatic intraepithelial neoplasias, whereas normal prostate tissue and, in most cases, prostatic hyperplasia were GRP receptor-negative. They also found GRP receptor expression in 4 of 7 bone metastases derived from hormone-refractory patients, albeit with very diverse receptor densities (6).
The panel of PC xenografts used in this study represents the different stages in tumor progression from androgen-dependent to androgen-independent (24,25). This panel is well established and has been used in a variety of PC-related studies showing their relevance as perfect tools in basic and translational research (26,38–40).
In vitro autoradiography on sections of all tumors of the panel using the radiolabeled BN receptor–binding 125I-universal ligand and the 111In-labeled BN analog [DTPA-Pro1,Tyr4]BN has shown that GRP receptors are predominantly expressed in the androgen-dependent xenografts. This GRP receptor binding largely disappeared in the androgen-independent but responsive and the completely androgen-independent tumors. Also, castration (androgen ablation) of mice bearing the androgen-dependent PC xenografts resulted in strongly reduced GRP receptor expression. GRP receptor expression of the androgen-responsive and androgen-independent xenografts was not altered. These results suggest that GRP receptor expression is regulated by androgens.
Further preclinical and clinical studies are required to determine whether androgen regulation of GRP receptor expression found in these PC xenograft models can also be established in PC patients. Androgen-regulated GRP receptor expression in PC patients would imply that PC patients having received standard hormonal therapy may have very low levels of GRP receptor. As a result, GRP receptor–based technologies may not be relevant in patients who have been pretreated by hormonal therapy. On the other hand, it should not be omitted that the androgen-independent PC-3 and DU-145 cell lines, though not clinically representative, express GRP receptors and that some bone metastases derived from hormone-refractory patients were found to be GRP receptor-positive (6).
The results of this study provide strong support for the use of GRP receptors for a sensitive new image modality for PC patients at the earliest time possible. Early detection and accurate staging at the time of diagnosis is essential and determines the treatment options available, as the presence of lymph node metastasis will make local treatment needless.
Also, a significant number of patients who have been treated locally by radical prostatectomy or radiation therapy will sooner or later show increasing levels of PSA of unknown origin and require additional therapy. These patients who have not yet been treated with additional hormonal therapy, but who are faced with generally low-volume metastases, may benefit from new systemic treatments using GRP receptor–based, BN-radiolabeled therapeutic analogs.
CONCLUSION
The results of this study suggest that in this model the overexpression of GRP receptors on prostate tumors is limited only to the androgen-dependent stages of PC. This overexpression of receptors is drastically reduced after androgen ablation in the animal model, indicating androgen regulation of the GRP receptor. Further studies are underway to determine GRP receptor density in tumor tissue derived from both hormonally treated and untreated patients as well as from patients at different stages of tumor progression. We are currently investigating the reversibility of castration-induced GRP receptor downregulation using the PC xenograft models.
Footnotes
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication August 11, 2006.
- Accepted for publication October 19, 2006.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Pilot Comparison of 68Ga-RM2 PET and 68Ga-PSMA-11 PET in Patients with Biochemically Recurrent Prostate Cancer
- Dosimetry and First Clinical Evaluation of the New 18F-Radiolabeled Bombesin Analogue BAY 864367 in Patients with Prostate Cancer
- International Union of Pharmacology. LXVIII. Mammalian Bombesin Receptors: Nomenclature, Distribution, Pharmacology, Signaling, and Functions in Normal and Disease States