


Abstract
Shortly after chemotherapy, relatively little is known about the expression of key genes and proteins involved in glycolysis. Doxorubicin (DOX) and 5-fluorouracil (5FU) are two commonly used chemotherapy agents that work through differing pathways. Glucose transporter-1 (Glut-1) and hexokinase II (HKII) proteins are highly expressed in many breast carcinomas, but their status while undergoing DOX or 5FU chemotherapy has not been systematically evaluated. Methods: We evaluated, in vitro, the messenger RNA (mRNA) and protein levels of Glut-1 and HKII in MCF-7, a breast adenocarcinoma cell line, and 3H-FDG uptake, both in untreated conditions and during treatment with either DOX or 5FU for 24 h. Six time points were evaluated: untreated at time 0; treated for 1 h; treated for 24 h; and 1, 2, and 3 d after chemotherapy. We analyzed tumor cell Glut-1 and HKII mRNA expression with real-time polymerase chain reaction and Western blotting, 3H-FDG uptake per cell, and cell viability with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide assay. Results: DOX was more effective than 5FU in killing the cancer cells under our study conditions. In untreated MCF-7 cells, the mRNA levels of HKII were typically higher than those of Glut-1, and 3H-FDG uptake was strongly related to Glut-1 mRNA expression (R2 = 0.85). Once treated with either drug, 3H-FDG uptake declined initially, the mRNA ratio was reversed, and Glut-1 mRNA levels were higher than HKII levels. This was verified in the protein assay. With DOX treatment, the cells showed increased Glut-1 mRNA and decreased HKII mRNA for the duration of active treatment; these levels returned to those seen in the untreated cells once the treatment was stopped for 24 h. However, HKII protein levels remained somewhat low. No correlation was seen between 3H-FDG uptake and HKII mRNA in DOX- and 5FU-treated cells (R2 = 0.14 and 0.0038, respectively). Conclusion: After DOX or 5FU therapy, the relationship between 3H-FDG uptake and viable cell number can become disjointed, with transient declines in 3H-FDG uptake in excess of the decline in cell number despite increased Glut-1 mRNA levels. This transient “stunning” has potential implications for 3H-FDG PET, especially soon after treatment is initiated. However, 3H-FDG remains a generally valid marker of viable cell number after cancer chemotherapy.
In the first hours and days after effective chemotherapy, relatively little is known about the expression of key genes involved in glycolysis. Many of the pathways of cells undergoing active chemotherapy are significantly altered while the cells attempt to maintain homeostasis. Once damage becomes irreversible through various treatment-specific pathways, cell death is triggered, initiating a programmed sequence of events ultimately leading to apoptosis.
Doxorubicin (DOX) is an anthracycline commonly used in the treatment of lymphomas, breast carcinoma, and other solid tumors. DOX has been reported to cause cell death through several pathways, including intercalating nuclear DNA (1); increasing cytosolic levels of H2O2, a reactive oxygen species (2); and causing the breakdown of the outer mitochondrial membrane that leads to the release of cytochrome c and initiates the caspase cascade ultimately leading to apoptosis (3,4).
In contrast, 5-fluorouracil (5FU), which has been in clinical use as an antineoplastic agent for almost 50 years, functions primarily by its inhibition of thymidylate synthase (5). Thymidylate synthase catalyzes the reductive methylation of 2′-deoxyuridine 5-monophosphate to generate thymidylate, an essential precursor for DNA synthesis, and its inhibition results in the cessation of cellular proliferation and growth (6). Indeed, there exist data that suggest a link between overexpression of thymidylate synthase and induction of a neoplastic phenotype (7).
PET is playing an increasingly important role in diagnosis, staging, and follow-up in the oncologic patient (8). Although many tracers have been used with PET, 18F-FDG has consistently demonstrated its value as a general tracer for use with this modality and takes advantage of the hypermetabolic state of neoplastic cells relative to their benign counterparts (9). One aspect of this hypermetabolism is the increased expression of the insulin-independent glucose transporter-1 (Glut-1) (10,11) and the type 2 isoform hexokinase (HKII) (12–14), which are important for the transport of glucose across the cell membrane and for the first step of glycolysis, respectively. In several carcinomas, their expression levels have been demonstrated to have prognostic significance (15).
18F-FDG PET is clearly of utility for monitoring the response of cancers to therapy (16). Because of the strong chemical and molecular changes induced in cells undergoing chemotherapy, we hypothesized that the typical relationship between 18F-FDG uptake and cell number seen in untreated cancer cells could be disrupted under these conditions. Zhou et al. have already demonstrated that several chemotherapeutic agents have a direct effect on the expression of Glut-1 and HKII (17). Additionally, a study has shown that 18F-FDG uptake is also affected by treatment (18). When combined, these findings, most notably those associated with 18F-FDG uptake, could be of significant importance to physicians in characterizing PET image findings in patients undergoing or having recently undergone chemotherapy.
Given the major roles played by Glut-1 and HKII in cellular metabolism, we sought to quantitatively determine their respective gene expression in a real-time environment in MCF-7 breast cancer cells—both untreated and undergoing chemotherapy with 1 of 2 common therapies, DOX or 5FU. These data were then compared with viability, Glut-1 and HKII protein levels, and 3H-FDG uptake data from cells under the same conditions during and soon after chemotherapy.
MATERIALS AND METHODS
Cell Culture and Chemotherapy
MCF-7 breast adenocarcinoma cells were commercially obtained from the American Type Culture Collection and grown in Dulbecco's modified Eagle medium fortified with 10% fetal bovine serum and 1% penicillin/streptomycin. Cells were incubated at 37°C with 5% carbon dioxide. Immediately before treatment, a sample was taken from each condition and labeled as time 0. Chemotherapy, in the form of either DOX (1 μmol/L) or 5FU (200 μmol/L), was then administered for 24 h in the growth medium. Samples were taken at 1 and 24 h with chemotherapy and labeled as time 1 h and time 24 h, respectively. Fresh, untreated medium was then given to the cells and samples were taken at 1, 2, and 3 d after chemotherapy (48, 72, and 96 h, respectively). In every case, untreated cells were grown, samples were taken at each corresponding time point, and fresh medium was given instead of DOX or 5FU.
Gene Expression
MCF-7 cells were grown in T75 flasks, one for each time point and condition, and brought to 35%–40% confluence before treatment in the manner stated previously.
cDNA Preparation.
For each sample, the cells were washed with phosphate-buffered saline (PBS) and given 1 mL of Buffer RLT (Qiagen, Inc.) with β-mercaptoethanol added, as prescribed. The cells were scraped, collected, and frozen at −80°C until extracted for total RNA using an RNeasy Mini Kit (Qiagen). RNA was quantitated using a spectrophotometer, load-equalized to 5 μg per reaction, and converted to cDNA using a SuperScript first-strand synthesis kit (Invitrogen).
Real-Time Analysis.
We used a real-time sequence detection system (ABI Prism 7900HT; Applied Biosystems) that applies a 2-cycle polymerase chain reaction program in which denaturing is performed at 94°C and both annealing and extension are performed at 60°C; probes and primers (TaqMan; Applied Biosystems) are designed with this in mind. Glut-1 and HKII probes contained a 6FAM label and TAMRA quencher. The β-actin probe used VIC and TAMRA. Reactions for each condition were performed separately, in triplicate, in a 384-well plate for 40 cycles. All treated/chemo data were analyzed using the Applied Biosystems SDS 2.1 software normalizing to β-actin levels and given as a percentage of its corresponding untreated cells. Sequences for the TaqMan primers and probes used are outlined in Table 1.
Primers and Probes Used in Real-Time Polymerase Chain Reactions
Protein Analysis
Protein Harvesting and Quantitation.
Cells were grown and treated or left untreated as already described. At the appropriate time point, the cells were washed with PBS, trypsinized, pelleted, and lysed with radioimmunoprecipitation assay buffer (Upstate Cell Signaling Solutions). Total protein amounts were measured with a bicinchoninic acid protein assay kit (Pierce Biotechnology, Inc.).
Western Blotting.
Twenty-five micrograms of protein were electrophoresed through either 8% or 10% Tris-glycine gels (Invitrogen) and electrophoretically transferred onto Immobilon-P (Millipore Corp.) membranes. After transfer, the membranes were subjected to standard blocking (KPL, Inc.) and incubation procedures and were incubated to the various proteins as follows: Glut-1 polyclonal, 1:1,000; HKII polyclonal, 1:8,000; and β-actin monoclonal, 1:1,000 (Chemicon International). The membranes were washed 4 times for 10 min in PBS with polysorbate and then further incubated for 1 h at room temperature with horseradish peroxidase–labeled secondary antibody (donkey antirabbit or goat antimouse IgG) diluted 1:4,000 in blocking buffer. After three 10-min washes in PBS with Tween, the membranes were treated with Immobilon Western Chemiluminescent HRP Substrate (Millipore Corp.) for 3 min and then were exposed on x-ray film (Denville Scientific, Inc.) for the required times (15–30 s) and developed. Densitometry was also performed on the digitized images.
3-(4,5-Dimethylthiazol-2-yl)-2, 5-Diphenyltetrazoliumbromide (MTT) Assay
Concurrently with the uptake study, approximately 350,000 MCF-7 cells were seeded into 6-well plates, 3 for each time point, and allowed to attach overnight. After treatment with either DOX or 5FU as already described, cell viability was assessed with an MTT cell proliferation kit (Roche Diagnostics) and treated according to protocol. Readings were made daily on a plate-reading spectrophotometer at 580 nm (cells) and 750 nm (background), subtracted, and plotted against a standard curve of known numbers.
3H-FDG Uptake
Approximately 350,000 MCF-7 cells were seeded into 6-well plates, 3 wells for each time point and condition, and allowed to attach overnight. At time 0, the cells were given either fresh medium or chemotherapy as already described. One hour before the appropriate testing point, the cells were washed with PBS and given low-glucose (1.0 g/L) Dulbecco's modified Eagle medium. They were then given 7.4 MBq (0.2 mCi) of tritiated FDG (3H-FDG) in a volume of 3 mL of low-glucose medium for another hour. During each step, the cells were kept incubated at 37°C with 5% carbon dioxide. After this uptake phase, the medium containing the 3H-FDG was removed and the cells were washed twice with ice-cold PBS. The cells were trypsinized with 400 μL of 0.25% trypsin and, once the cells lifted, were lysed with 500 μL of NaOH (0.33 mol/L) containing 1% sodium dodecylsulfate. The contents were placed into scintillation vials, given 10 mL of scintillation fluid (Formula 989; Perkin-Elmer, Inc.), and counted in a β-counter.
RESULTS
MTT Assay
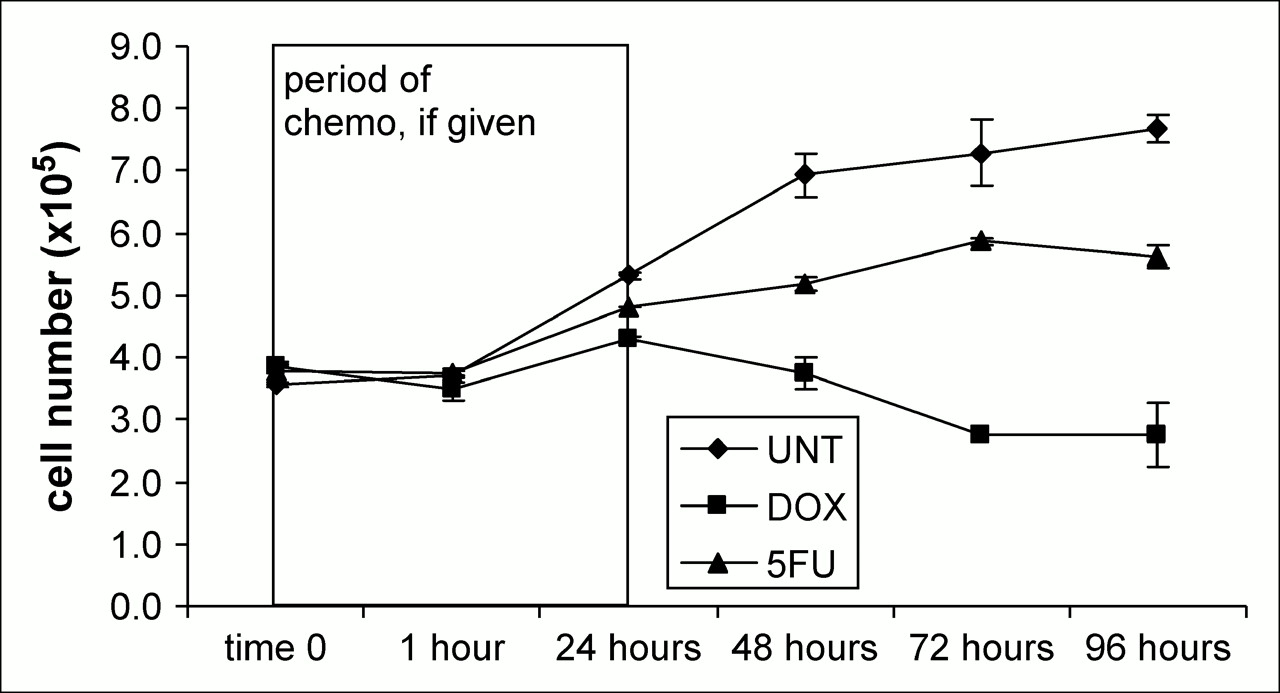
Cell number was determined from the MTT assay for each time point and condition and plotted in Figure 1. When applied using the method and doses described previously, DOX is the more effective chemotherapy agent for treating breast carcinoma cells. A growth divergence is seen at 24 h, and untreated cells are 49% more numerous than at time 0, whereas DOX- and 5FU-treated cells are more numerous by 11% and 28%, respectively. At the time corresponding to 1 day after chemotherapy, untreated cells were 94% greater than at time 0. By contrast, changes of −3% and 37% were seen in DOX- and 5FU-treated cells, respectively, at these same times. At the last time point, 3 d after chemotherapy, overall changes in viable cell number by MTT for the untreated, DOX-treated, and 5FU-treated cells were 115%, −28%, and 48%, respectively.

Actual cell number as calculated from standard curve.
3H-FDG Uptake
3H-FDG uptake values per cell and per well were obtained and are reported in Figure 2 and given as a percentage of the values at time 0. As expected, the untreated cells showed the highest uptake per cell and per tissue-culture well. Results from the real-time analysis of Glut-1 and HKII messenger RNA (mRNA), along with those from MTT and 3H-FDG uptake assays in untreated and treated MCF-7 cells, are shown in Figures 3–5⇓. In the untreated state, 3H-FDG per cell and MTT per cell increased gradually with proliferation (Fig. 3). In the DOX-treated cells, a decrease in 3H-FDG uptake was seen at 24 h of treatment (Figs. 2 and 4), and with 5FU this decrease was seen earlier, after 1 h of treatment (Figs. 2 and 5). With both treatments, 3H-FDG uptake and viable cell number, by MTT, generally moved in parallel.
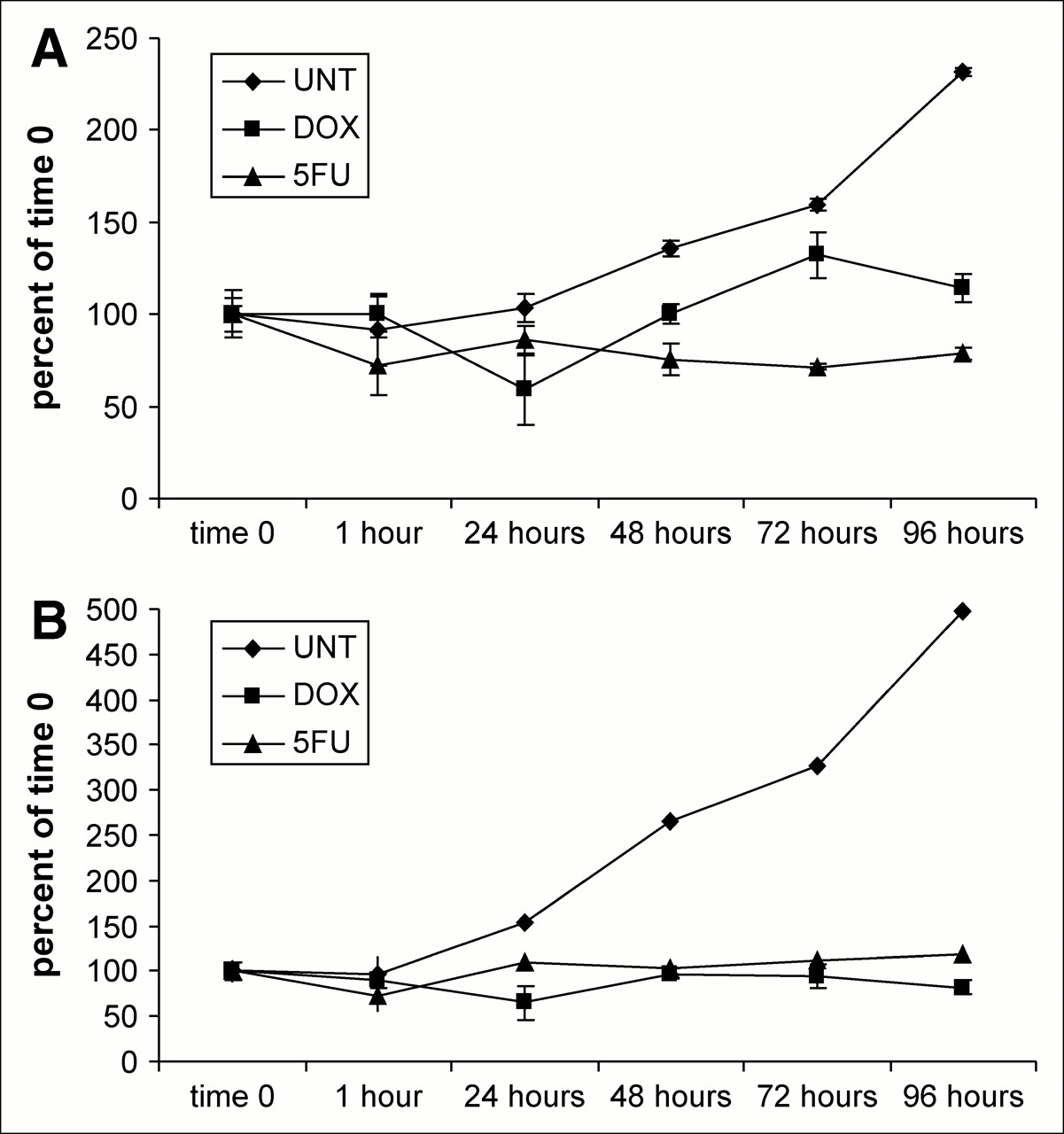
3H-FDG uptake per cell (A) and per well (B) in untreated and treated MCF-7 cells.
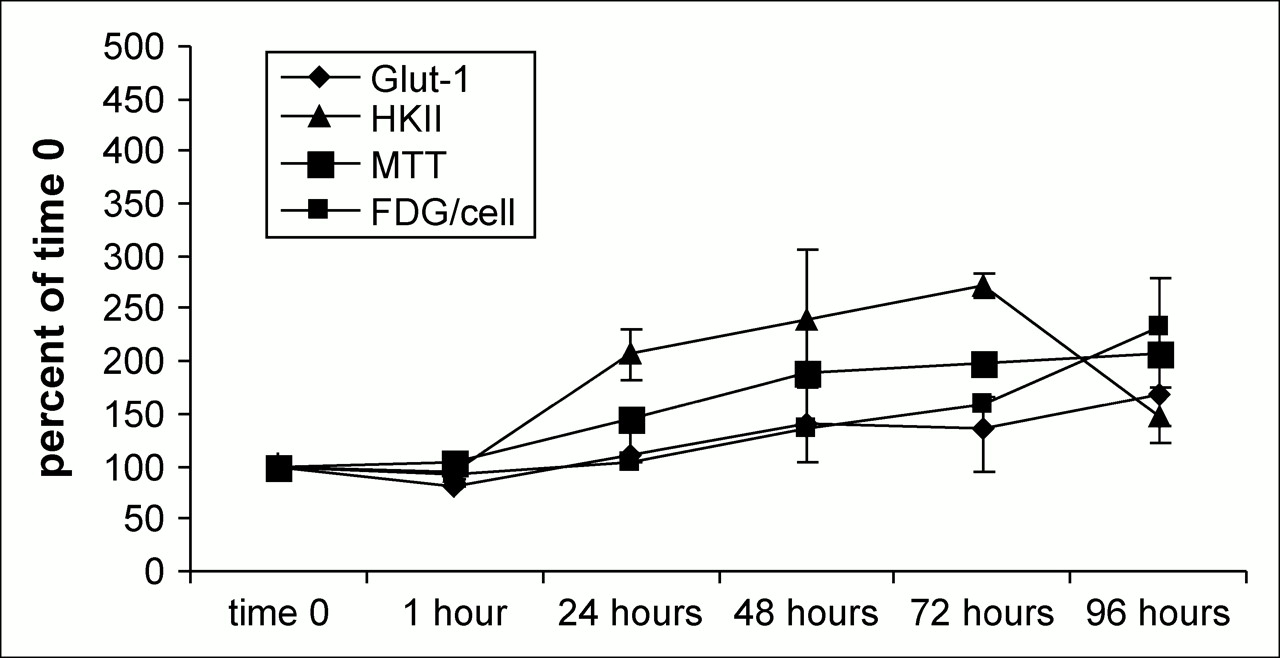
Comparison of glut-1 mRNA, HKII mRNA, 3H-FDG per cell, and MTT results in untreated MCF-7 cells.

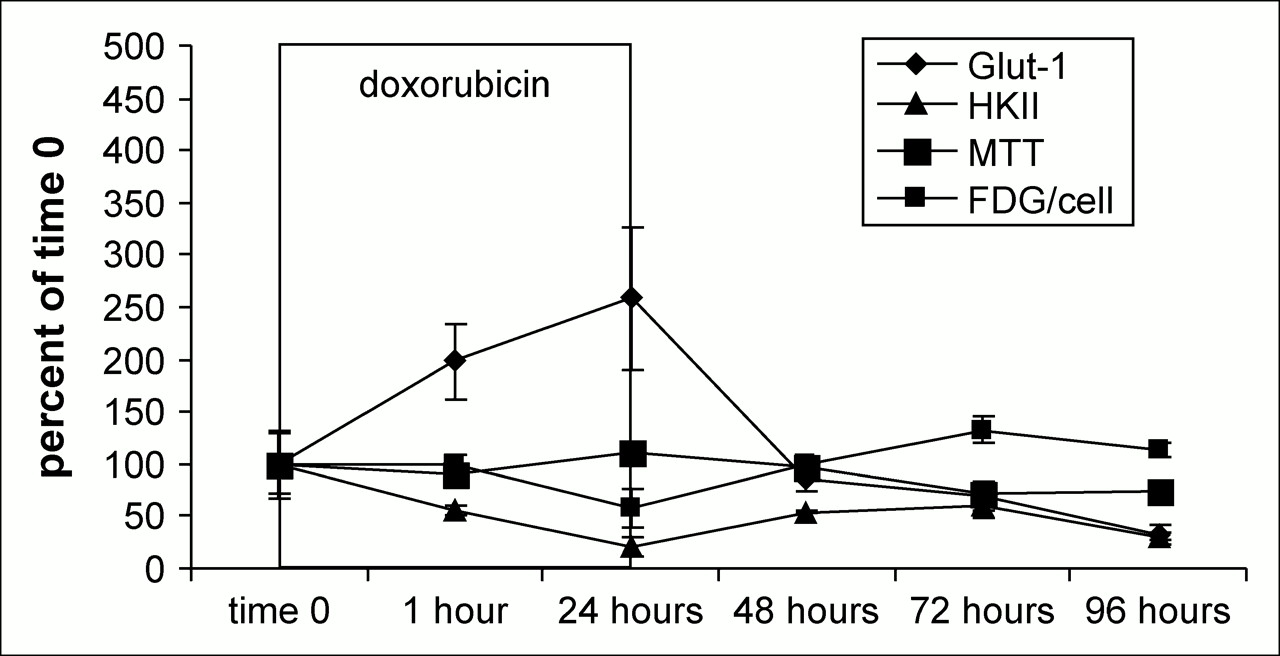
Comparison of glut-1 mRNA, HKII mRNA, 3H-FDG per cell, and MTT results in DOX-treated MCF-7 cells.
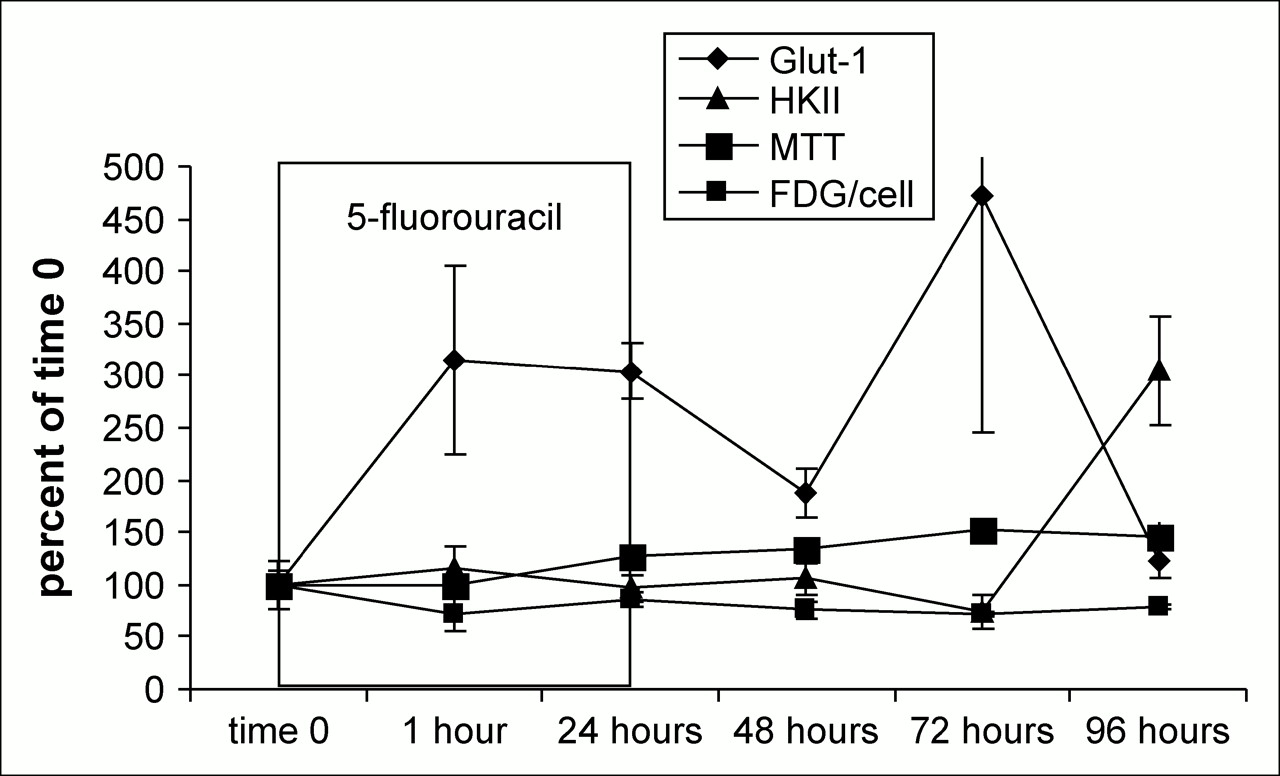
Comparison of Glut-1 mRNA, HKII mRNA, 3H-FDG per cell, and MTT results in 5FU-treated MCF7 cells.
Glut-1 and HKII mRNA Expression
Untreated Cells.
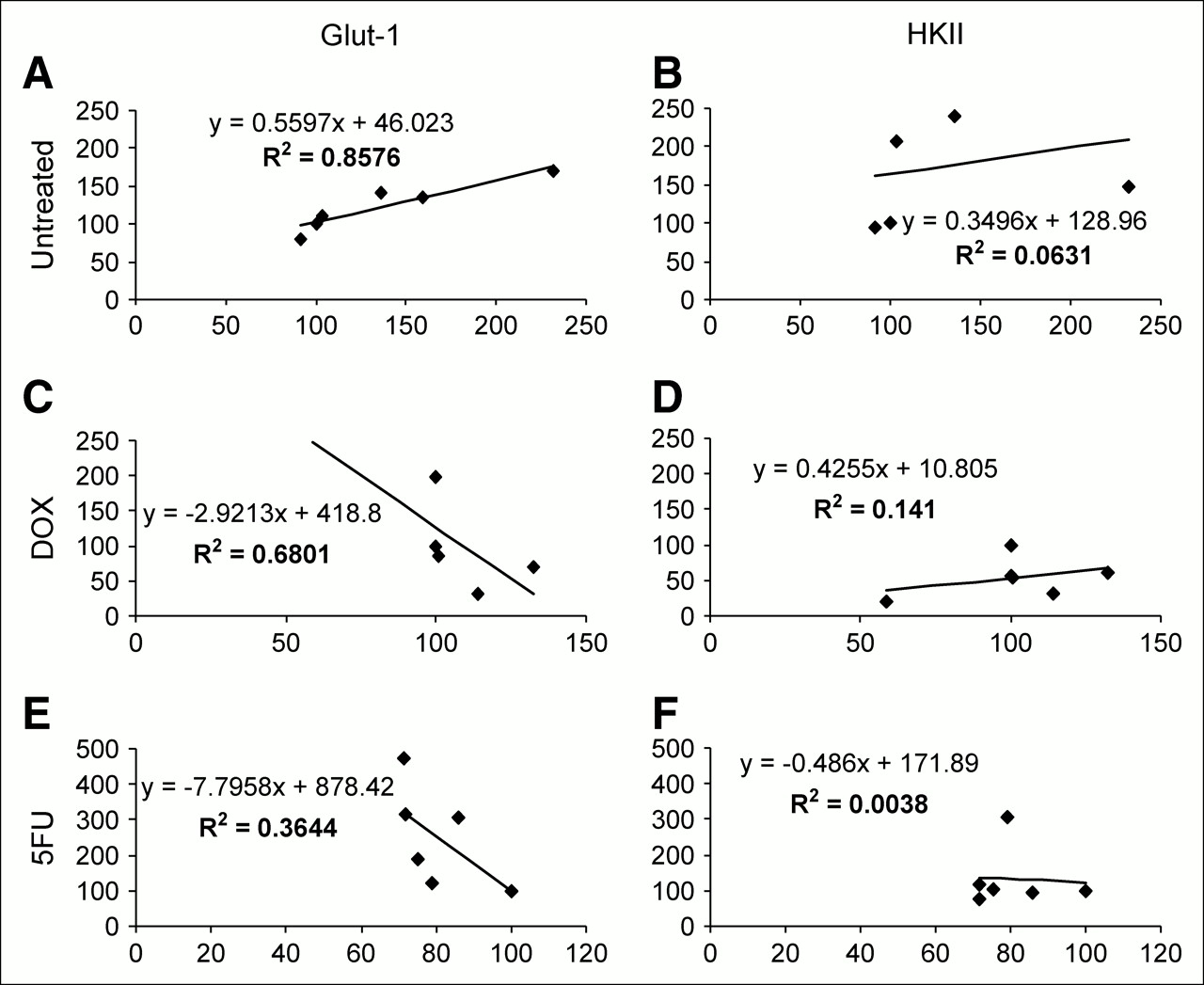
With the exception of the final, 96-h, time point, the ratio of HKII mRNA levels to Glut-1, normalized to time 0, were higher (Fig. 3). At some time points, the HKII mRNA was nearly 2 times higher than its time 0 values. This was observed in the real-time polymerase chain reaction for both DOX and 5FU when untreated cells were used as both normalization and comparative factors for determining the functional up- or downregulation of the test genes. 3H-FDG uptake was strongly related (R2 = 0.85) to the levels of Glut-1 message, whereas the relationship of 3H-FDG uptake to HKII message was weak (R2 = 0.063) in the untreated cells.
Treated Cells.
The expression of mRNA for Glut-1 and HKII in DOX- and 5FU-treated MCF-7 cells, as well as MTT and 3H-FDG uptake data, are shown in Figures 4 and 5. With both DOX and 5FU treatments, the relative levels of Glut-1 mRNA to HKII mRNA were high during the 24 h in which the cells received treatment. After treatments, Glut-1 message levels become inversely proportional to 3H-FDG uptake with DOX and 5FU treatment, and the R2 values for these correlations were 0.68 and 0.36, respectively.
Statistical Analysis
Mean values for the expression of untreated and DOX- and 5FU-treated Glut-1 and HKII, and mean values obtained from the 3H-FDG uptake study, given as uptake per cell, were subjected to regression analysis for the 6 time points tested. Correlation coefficients were determined, and these results are shown in Figure 6.

Results of regression analysis using real-time mRNA data and 3H-FDG uptake per cell: untreated vs. glut-1 (A), untreated vs. HKII (B), DOX vs. glut-1 (C), DOX vs. HKII (D), 5FU vs. glut-1 (E), and 5FU vs. HKII (F). y-Axis represents percentage of baseline expression of mRNA; x-axis represents percentage of baseline 3H-FDG uptake.
Western Blotting and Protein Evaluation
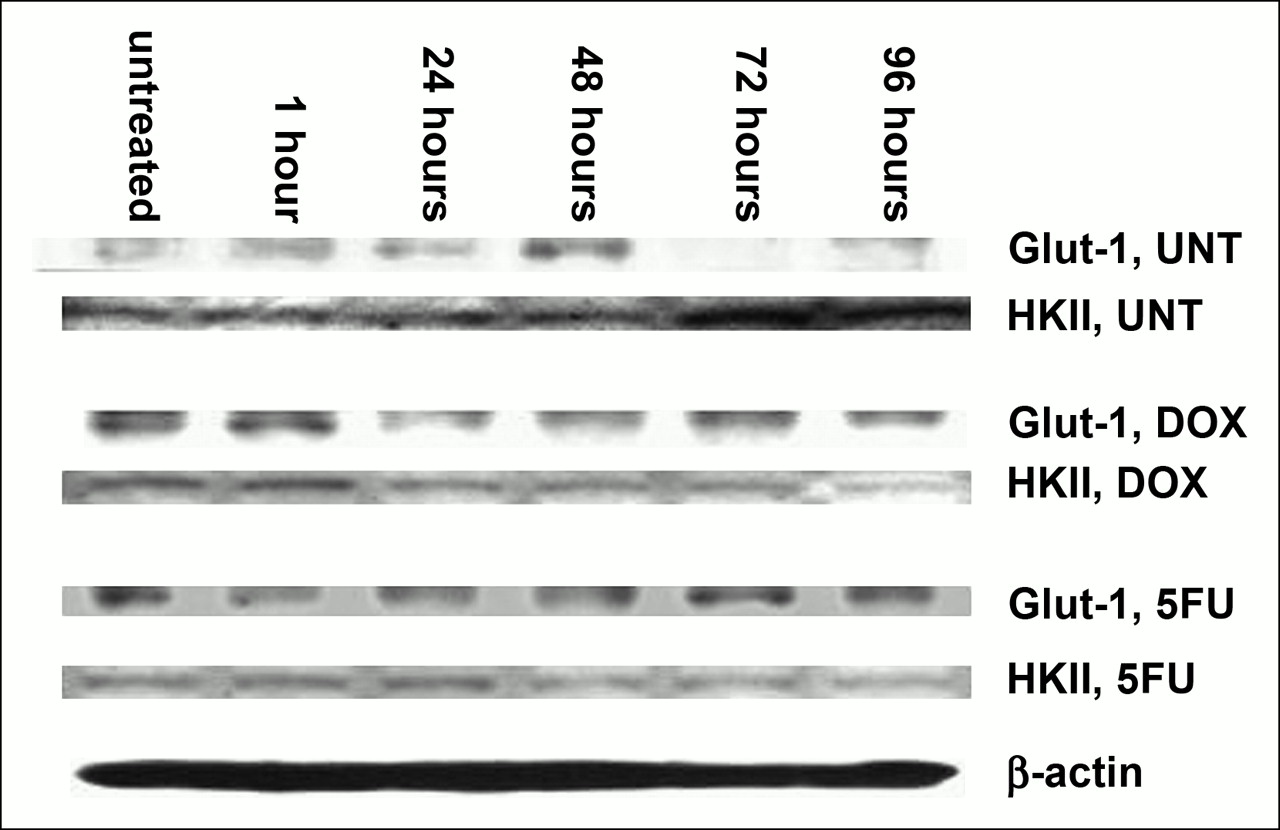
The results of Western blotting on untreated and DOX- and 5FU-treated MCF-7 cells are shown in Figure 7. These results show, much as for the mRNA studies, that HKII levels rise in untreated cells and decline in treated cells. Glut-1 protein levels did not rise in parallel to the increase in Glut-1 mRNA levels in treated cells. At the time of DOX-induced “stunning,” both Glut-1 and HKII protein levels appeared somewhat decreased.
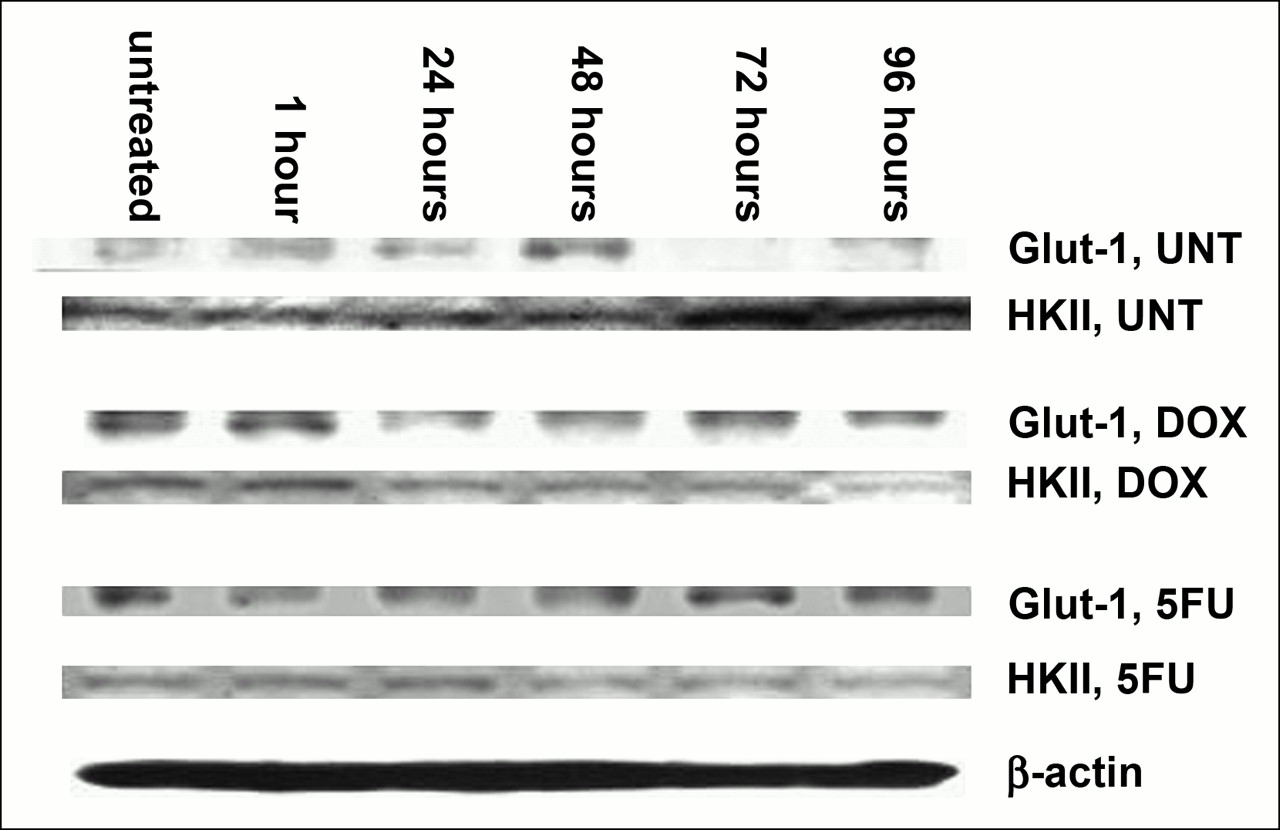
Western blot analysis of total protein extractions from untreated and DOX- and 5FU-treated MCF-7 cells. Comparable amounts of total protein are loaded in each lane, as shown by β-actin. HKII protein rises in untreated cells and falls in treated cells. Glut-1 protein levels change less than does mRNA. UNT = untreated.
DISCUSSION
DOX was the more effective chemotherapy agent used in our study at the doses given. Its two-pronged method of cell killing works first with the inducement of the reactive oxygen species, which increases mitochondrial permeability, releasing cytochrome c and initiating apoptosis. This effect is compounded by the ability of DOX to intercalate DNA, affecting its synthesis and that of RNA. Structurally, HKII associates with the cytoplasmic face of the outer mitochondrial membrane through an amino-terminal hydrophobic domain and uses the abundant adenosine triphosphate for the phosphorylation of glucose (19). Though not tested in this study, it is likely that the disruption of the mitochondrial membrane by DOX would be significantly detrimental to this process.
For the duration of DOX exposure, Glut-1 mRNA levels increase by as much as 2.5 times over untreated levels and then return to a relatively normal range within 24 h after removal of the drug. However, the maximum effects of DOX on Glut-1 protein expression are not as dramatic at 24 h of exposure as is seen in the mRNA data, likely demonstrating the role of posttranslational alterations in the final protein expression. Indeed, Glut-1 protein appears diminished slightly after 24 h of DOX treatment. Conversely, HKII mRNA levels decrease throughout DOX exposure, and the same recovery effect is seen on its removal. Interestingly, the decrease in HKII seen in the mRNA data is also apparent in the protein data, particularly during the 24 h of DOX treatment.
It is at this 24-h time point that a phenomenon we describe as a “stunning” effect can be seen. In this phenomenon, the relationship between 3H-FDG uptake and viable cell number becomes disjointed after 24 h of treatment with DOX. 3H-FDG uptake becomes less than expected for the number of viable cells, despite the fact that in all other cases in untreated tumors this relationship has proven to be very consistent. With DOX treatment, this phenomenon is seen only at this time point—the time point at which, despite increased Glut-1 mRNA levels, Glut-1 and HKII protein levels decline. Thus, DOX appears to be interfering with the successful translation of Glut-1 mRNA to Glut-1 protein, with a relevant decline in 3H-FDG uptake. After the maximum decrease of 3H-FDG uptake, observed at 24 h, the MCF-7 cells appear incapable of recovery.
5FU is an antineoplastic agent and works in a simpler manner, mainly through the inhibition of thymidylate synthase and, ultimately, of cell growth through inhibition of thymidylate, an essential precursor for DNA synthesis. Like DOX, 5FU also elevates Glut-1 mRNA, only the effects are longer lasting. The increase in Glut-1, particularly at later time points, is evident in the protein analysis. HKII mRNA levels increase after 1 h of treatment, and possibly another, milder, “stun” is observed at that time. The decrease in 3H-FDG uptake is not as pronounced as is that seen with DOX, and the cells are much better able to recover from this treatment under the conditions in which it was given. With the removal of 5FU, we see a steady increase in 3H-FDG uptake that is mirrored in the MTT data. The relationship between 5FU treatment and 3H-FDG uptake is less clear than that between DOX treatment and 3H-FDG uptake, suggesting that some cellular component other than Glut-1 or HKII plays a larger role in the accumulation of FDG-6-phosphate. Changes in mitochondrial HKII levels might be responsible, but this possibility was not evaluated in our study.
The MTT cell viability assay determines cellular levels of MTT, which produces the formazan that is used as an indicator of cellular metabolism. The presence of formazan indicates metabolic reduction, and the amount of formazan relates to the number of metabolic reducing equivalents produced, such as reduced nicotinamide adenine dinucleotide or reduced nicotinamide adenine dinucleotide phosphate. Although metabolism is an indicator of cell viability, we believed it necessary to clarify our measurement approach. Of concern is the ability of succinate dehydrogenase, a necessary component of cellular metabolism, to act as an electron donor and therefore affect MTT results. Berridge (20) has shown that succinate contributes minimally to MTT reduction and that the terms metabolism and viability can be used synonymously.
Our Western blot analysis was performed using total protein lysates. At the molecular level there are other considerations, such as intracellular localization and compartmentation of Glut-1 and HKII, that one must take into account when addressing the metabolic roles of these proteins. Aloj et al. (21) addressed this notion for Glut-1 and HKII by analyzing lysates from both whole-cell extracts and isolated plasma membrane extracts. With chemotherapy, this type of study would provide more insight into the role of Glut-1, which is membranous. Additionally, obtaining fractionalized samples of mitochondria and mitochondrially attached proteins would help in the determination of HKII protein levels actively involved in glycolysis, as opposed to nascent, unattached HKII. This subcellular localization is a topic of interest and should be addressed in future work.
CONCLUSION
Our data demonstrate that 3H-FDG accumulates less in MCF-7 cells treated with either DOX or 5FU than in untreated MCF-7 cells and that, during treatment, 3H-FDG uptake may decline more than viable cell number (“stunning”). In the first 24 h of treatment with either drug, Glut-1 mRNA levels rise substantially, and Glut-1 protein levels show a much more modest change. HKII appears to have a more direct impact on 3H-FDG uptake, particularly in DOX-treated cells, in which declines in HKII mRNA expression, HKII protein, and 3H-FDG uptake per cell seem to move in parallel. Despite these treatment effects, a generally strong relationship between tumor cell 3H-FDG uptake and viable cell number remains after treatment, supporting, on a basic level, the use of 3H-FDG to monitor cancer treatment at most times after chemotherapy.
References
- Received for publication December 20, 2005.
- Accepted for publication December 23, 2005.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Comparison of EORTC Criteria and PERCIST for PET/CT Response Evaluation of Patients with Metastatic Colorectal Cancer Treated with Irinotecan and Cetuximab
- Interim FDG-PET in Hodgkin lymphoma: a compass for a safe navigation in clinical trials?
- PET/CT for Therapy Response Assessment in Lymphoma
- From RECIST to PERCIST: Evolving Considerations for PET Response Criteria in Solid Tumors
- 18F-FDG PET/CT for Monitoring the Response of Lymphoma to Radioimmunotherapy
- Colorectal Tumor Cells Treated with 5-FU, Oxaliplatin, Irinotecan, and Cetuximab Exhibit Changes in 18F-FDG Incorporation Corresponding to Hexokinase Activity and Glucose Transport
- Integrating PET and PET/CT into the Risk-Adapted Therapy of Lymphoma