


Abstract
Further development of nuclear medicine for imaging and internal radiotherapy demands a precise knowledge of the tissue and cellular distribution of radiopharmaceuticals. Ion microscopy (secondary ion mass spectrometry [SIMS]) may be particularly useful in this respect. We used SIMS to study the biodistribution of the melanoma-targeting molecule N-(2-diethylaminoethyl)-4-iodobenzamide (I-BZA), both in its native state and radiolabeled with 14C. Methods: C57BL6/J1/co mice bearing pulmonary colonies of B16 melanoma cells were injected with I-BZA or 14C-I-BZA. Appropriate tissues were fixed and included in epoxy embedding resin for SIMS studies. The distribution of unlabeled I-BZA was studied by detecting its stable iodine atom (127I). 14C-I-BZA distribution was studied by dual detection of 127I and 14C. The time course of I-BZA concentrations at sites of tissue fixation was studied by measuring the signal ratio of 14C and the naturally occurring isotope 13C. Results: SIMS showed that I-BZA concentrated in the cytoplasm of tumoral melanocytes (melanoma cells) and in the cytoplasm of tumor-infiltrating macrophages (melanophages). I-BZA was also detected in the cytoplasm of normal melanocytes in the pigmented structures of skin and eye. Interpretation of I-BZA distribution by using electron micrographs of adjacent sections showed that the intracytoplasmic melanin-rich organelles (melanosomes) were responsible for I-BZA retention. The distributions of 127I and 14C after 14C-I-BZA injection were identical, even when I-BZA was separately labeled with 14C at 2 different positions, indicating the stability of the amide bond of I-BZA. The time course of the 14C/13C ratio in the melanosomes of melanoma cells suggested a retention half-life of about 38 h. Conclusion: Contrary to previous suggestions that I-BZA fixes principally to σ-1 membrane receptors, our results strongly indicate that I-BZA associates with intracytoplasmic melanin pigments. Early I-BZA accumulation, in both melanocytes and melanophages, suggests that this compound fixes to preformed melanin rather than being incorporated during de novo melanin synthesis. These quantitative and qualitative data obtained with I-BZA illustrate the excellent potential of SIMS for studying the biologic fate of radiopharmaceuticals.
Further therapeutic and diagnostic developments in nuclear medicine demand a better knowledge of radiopharmaceutical distribution (1,2). Classic autoradiography and, more recently, β-imagers can be used to study radiopharmaceutical distribution in a given organ, with a resolution close to 50 μm. However, studies of cellular or subcellular distribution require the use of techniques with even higher resolution, such as microautoradiography (1,3) and, more recently, ion microscopy (secondary ion mass spectrometry [SIMS]).
Based on mass spectrometric detection, SIMS can offer images of the tissue or cellular distribution of both stable and radioactive atoms (4–6). Molecules containing a native halogen atom (fluorine, iodine, or bromine) can be detected without radiolabeling (7–9), whereas other molecules must first be labeled with either a stable isotope such as 15N (10–12) or a radioactive isotope such as 14C (10,13,14).
We used SIMS to study the cellular distribution of N-(2-diethylaminoethyl)-4-iodobenzamide (I-BZA), a compound targeting malignant melanoma. Initial studies on animals showed a high concentration ratio between grafted melanomas and various normal tissues (15). These results were confirmed in patients with cutaneous or ocular melanoma in a phase II trial with 123I-BZA (16,17). Subsequent studies further supported the potential value of I-BZA (18–20) and related compounds (21–26) for the pathologic staging of melanoma, together with other tracers such as 18F-FDG (27).
The high selectivity of I-BZA raises the possibility of therapeutic use in patients with metastatic melanoma, a disease with limited treatment options (28,29). However, a precise knowledge of the sites of accumulation in both tumoral and healthy tissues is a prerequisite for clinical use.
The I-BZA fixation site is controversial. John et al. (21–22) and others (20,30) have suggested that I-BZA and the related analog 2-piperidinylaminoethyl-4-iodobenzamide accumulate in melanomas by binding to σ-receptors on the cell membranes (20–22,30). Competitive binding studies using guinea pig brain membranes and (3H)(+)-pentazocine, a selective σ-1 ligand, showed that both I-BZA and the related analog had high affinity for σ-1 sites. However, Chehade et al. (31) and others (23,24,32) have suggested that I-BZA fixes preferentially to melanin granules. Proponents of this second hypothesis disagree on the underlying mechanism. Dittmann suggests that I-BZA is incorporated during melanin synthesis (23), whereas Labarre suggests that I-BZA binds to preformed melanin (32).
The SIMS images presented here were obtained with various tissue samples from mice bearing pulmonary colonies of melanoma B16 cells. The study comprised 3 steps: We first used an IMS-4F scanning ion microscope (CAMECA) to map the distribution of unlabeled I-BZA in tumor tissue and normally pigmented tissues (eye and skin) by detecting the stable 127I atom. Our initial observations (31,33), obtained with a first-generation low-resolution ion microscope (SMI-300; CAMECA), allowed us to select the tissue regions of interest for this study. We then compared images of 127I distribution with those of a 14C radiolabel. Separate experiments were performed with I-BZA labeled by 14C at 2 different positions, to obtain further information on the structure of the molecule at its tissue-binding site. Finally, we studied the time course of 14C-I-BZA concentrations at cellular retention sites.
MATERIALS AND METHODS
This study was performed in keeping with French legislation on animal experimentation.
Tumor Model, 127I-BZA Administration, and Tissue Preparation
B16 murine melanoma cells were injected intravenously into adult male C57BL6/J1/co mice (Iffa Credo) to obtain tumor cell colonies mimicking pulmonary micrometastases (33). Briefly, transplantable B16 mouse melanoma cells, obtained from Institut de Cancérologie et Immuno-Génétique, were maintained as monolayers in minimal essential medium supplemented with 10% fetal bovine serum and antibiotics and passaged by trypsinization. For transplantation, melanoma cells were trypsinized and washed with phosphate-buffered saline, then resuspended in phosphate-buffered saline (1.5 × 106 viable cells per milliliter); 0.2 mL of this suspension was injected intravenously. Multiple tumor colonies developed in the lungs within 3 wk.
I-BZA was injected into tumor-bearing and control animals via a tail vein (2.6 μmol per animal). Three groups of tumor-bearing and control animals were sacrificed by CO2 inhalation at different times. The lungs, eyes, and tail skin were removed, fixed with glutaraldehyde, and embedded in epoxy resin. To verify that the fixation steps do not lead to significant release of the compound, we measured radioactivity in the solutions during each step of tissue preparation. This was done in our early animal studies using 125I-BZA (15) and confirmed with 14C-I-BZA.
Serial 2-μm-thick slices were placed on gold plates for SIMS and on glass slides for light microscopy. Samples were also prepared for electron microscopy.
SIMS
The basic principles of this method are described in detail elsewhere (4). In a SIMS device, the tissue section is bombarded with ions with an energy of about 15 keV, known as the primary ion beam. This bombardment breaks most chemical bonds, and the atoms constituting the sample are ejected (“sputtered”). The sample is gradually sputtered, starting with the most superficial layers of atoms. Atoms are ejected from the sample surface as charged particles and are guided by an electrostatic field, resulting in a secondary ion beam. The latter is directed toward a mass spectrometer tuned to detect a given ion species, yielding an image that reflects the distribution of the selected ion species.
The ejected iodine atoms are detected by SIMS in the form of negatively charged ions (127I−). The resulting map of 127I distribution reflects the tissue and cellular distribution of I-BZA or its metabolites. The distribution of 14C-I-BZA is visualized by dual detection of 127I− and 14C− ions (10,14). Tissue structures can also be identified by SIMS, by detecting highly emissive CN− polyatomic ions, which reflect tissue and intracellular nitrogen content, or by imaging the distribution of sulfur ions (32S−). Imaging of phosphorus ions (31P−), which are mainly emitted from nucleic acids, can be used to identify the cell nucleus.
The instrument used for this study was an IMS-4F scanning ion microscope. Samples were bombarded with a primary beam of positively charged cesium ions (Cs+), offering high yields of 127I− and 14C−. The minimum diameter of the Cs+ beam in this device is 0.3 μm, offering a resolution of 0.5–1 μm on the recorded images. Analyses were performed at a high mass resolution (M/ΔM = 3,000) to avoid interference by polyatomic groups. Indeed, a mass resolution of M/ΔM ≥ 1,750 is necessary to separate 14C− (14.0032 amu) from the closest interference due to 13C1H− (14.0112 amu) (10,14). The selected ions are counted by the detector (an electron multiplier), directly yielding digital distribution images.
14C Labeling and Comparison of 127I and 14C Distribution
To assess the stability of the amide bond (CO-NH), we separately labeled I-BZA with 14C at 2 positions on either side of this bond—precisely, on the carbonyl group I-φ-14CO-NH-CH2-CH2-N[C2H5]2 (14C-BZA-carbonyl, 151.7 kBq/μmol) and the N-diethylamine group I-φ-CO-NH-CH2-14CH2-N[C2H5]2 (14C-BZA-amine, 188.7 kBq/μmol). The status of the carbon chain (intact/broken) was determined by comparing the images of 127I and 14C distribution.
Estimation of Retention Half-Life of I-BZA at Cellular Level
14C labeling was also used to study the time course of I-BZA concentrations at the sites of retention.
The iodine signal cannot be used to compare I-BZA concentrations from one field to another or from one sample to another, because it is susceptible to the experimental conditions. In contrast, this comparison is possible for a 14C-labeled compound (14), by measuring the local isotopic ratio between 14C and 1 of the 2 stable, naturally occurring carbon isotopes, 12C or 13C. We chose to measure the 14C/13C ratio rather than the 14C/12C ratio because 12C, being highly abundant, can saturate the detector. The isotopic abundance of 13C is close to 1.1%. Naturally occurring 14C, which has an isotopic abundance of 1.2 × 10−12, yields no signal under our analytic conditions (data not shown).
RESULTS
SIMS images were obtained with tissues sampled 3 h after I-BZA injection. (According to Michelot et al., uptake is maximal as early as the first hour (15).)
I-BZA Distribution in Epidermal Layer of Skin
Figure 1 shows SIMS images of mouse tail skin (field, 50 × 50 μm). Part A shows the distribution of CN− ions (mass, 26), revealing various tissue structures, including, from top to bottom, the stratum corneum, the other epidermal layers, and the superficial dermis. Part B shows the distribution of phosphorus (mass, 31), clearly identifying the cell nuclei, especially in the deeper epidermis. The most emissive structures correspond to nuclear membranes (heterochromatin) and nucleoli. Part C shows the distribution of iodine (mass, 127). Several distinct foci of uptake are visible, each with a size approaching 1 μm. These foci are especially abundant in the deeper epidermis. Part D shows superimposed artificially colored images, in which green corresponds to phosphorus and red to iodine. The red foci in the basal epidermis show an intracytoplasmic, perinuclear distribution.

SIMS images of a 50 × 50 μm field of skin: distribution of 26CN− ions (A), 31P− ions (B), and 127I− ions (C) and superimposition of iodine signal in red and of phosphorus signal in green (D).
I-BZA Distribution in Choroid and Pigmented Retinal Epithelium
Figures 2A and 2B are SIMS images of an ocular section (field, 200 × 200 μm). Part A shows the tissue distribution of 127I, with 2 parallel bands. Part B, a superimposition of the 127I signal (red) on the 26CN signal (gray), shows that the broadest (outer) band corresponds to the choroid and the thinnest band to the pigmented retinal epithelium. The other ocular layers seen on the 26CN map (the outer sclerotic and the inner layers corresponding to the cones and rods and to the nuclei of these cells) show no I-BZA uptake.

(A and B) SIMS images of a 200 × 200 μm field of eye: distribution of 127I (A) and superimposition of iodine signal in red and of CN signal in gray (B). (C) Superimposition of iodine signal in red and of phosphorus signal in green in a 50 × 50 μm field centered on choroid and pigmentary epithelium. (D) Electron micrograph of choroidal melanocytes in a 6 × 6 μm field.
We then focused on a 50 × 50 μm field that included the choroid and pigmented retinal epithelium. Figure 2C shows superimposed 31P (green) and 127I (red) signals. The 31P signal shows 2 nuclear aspects: The elongated nuclei correspond to the choroid, and the more rounded nuclei to the underlying pigmented epithelium. As in the skin, the 127I uptake foci are located in the cytoplasm.
Figure 2D is an electron micrograph of the principal cell—melanocytes—constituting the choroid. Comparison of this image with the ion micrograph suggests that the multiple bright cytoplasmic foci of 127I correspond to melanosomes, the intracytoplasmic organelles that synthesize melanin (34). Melanosomes are abundant in the cytoplasm of choroidal melanocytes, explaining why the iodine signal is intense and nearly confluent in these cells.
I-BZA Distribution in Pulmonary Melanoma Cell Colonies
Figures 3A and 3B are SIMS images of a murine B16 melanoma cell colony developing in the alveolar parenchyma (field, 60 × 60 μm). Part A shows that the tissue distribution of 127I is highly heterogeneous. There are multiple small, scattered foci measuring 1 μm or less. There is also a larger, 127I-rich, site about 15 μm in diameter. Part B shows the superimposition of the 31P image (green, representing nuclei) and the 127I image (red): The 2 signals are distinct, confirming that I-BZA has no nuclear location.
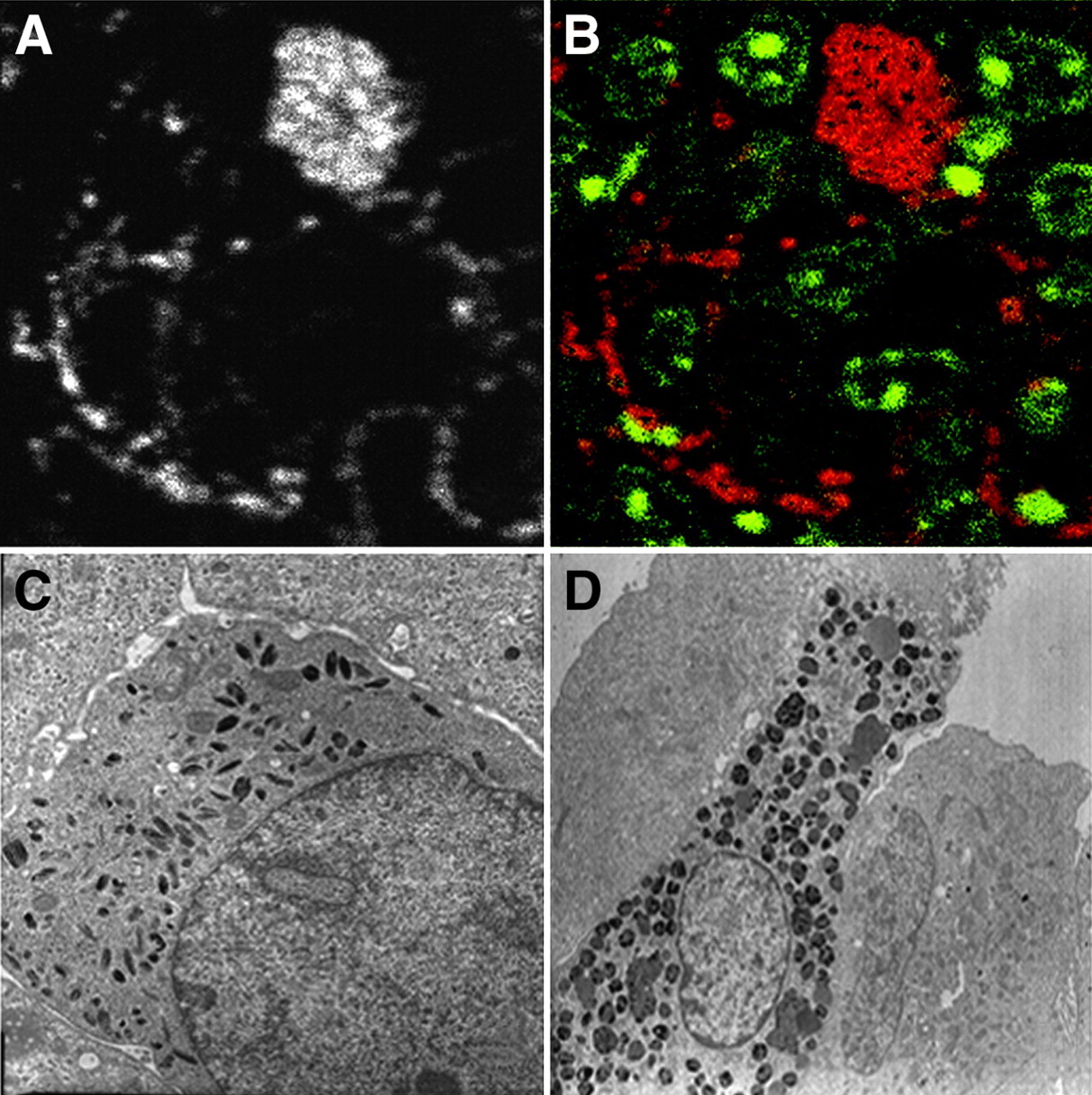
(A and B) SIMS images of a 60 × 60 μm field focusing on part of a pulmonary colony of B16 melanoma: distribution of 127I (A) and superimposition of iodine signal in red and of phosphorus signal in green (B). (C) Electron micrograph of a melanoma cell in an 8 × 8 μm field. (D) Electron micrograph of a tumor-infiltrating macrophage (melanophage) in a 23 × 23 μm field.
Comparison with the electron micrographs (Figs. 3C and 3D) facilitates cell identification and helps explain the iodine distribution. Most of the cells composing the tumor are tumoral melanocytes, and their cytoplasm contains small melanosomes of variable shape (Fig. 3C). The cell containing abundant iodine corresponds to a tumor-infiltrating macrophage (melanophage). These cells show multiple intracytoplasmic complexes, containing melanosomes and other melanin-rich phagocytotic products (Fig. 3D).
Comparison of 127I and 14C Distribution
After Injection of 14C-I-BZA Amine.
SIMS images obtained 1, 14, and 72 h after injection showed a similar distribution of 14C and 127I, both in the eye and in pulmonary colonies of B16 melanoma cells. Figure 4 shows SIMS images of the eye 1 h after injection. Part A shows superimposed images of the phosphorus signal (green) and the iodine signal (red). Part B shows superimposed images of the 31P signal (green) and the 14C signal (violet). These images confirm the similar topography of 14C and 127I, both of which are found in the cytoplasm of melanocytes present in the choroid and retinal pigmented epithelium. The 127I images are more strongly contrasted than the 14C images because 127I, a readily ionized halogen, has a stronger SIMS yield than does 14C. The specificity of I-BZA and 14C-I-BZA imaging was assessed by studying controls prepared under the same conditions.

SIMS images of a 50 × 50 μm field centered on choroid and retinal pigmented epithelium, 1 h after 14C-I-BZA amine injection: superimposition of iodine signal in red and of phosphorus signal in green (A), and superimposition of 14C signal in violet and of phosphorus signal in green (B).
After Injection of 14C-I-BZA-Carbonyl.
Here too, the analysis of images obtained at 2 different times (2 and 14 h after injection) showed that the distributions of 127I and 14C were identical in the pigmented eye layers and in pulmonary colonies of B16 melanoma cells.
Biologic Half-Life of I-BZA in Melanosomes of Melanoma Cells
In view of the imaging results, we studied the time course of the 14C/13C ratio in the cytoplasm of tumoral melanocytes. Intracytoplasmic measurements were focused on regions of interest (1.6 × 1.6 μm) encompassing foci of 14C (melanosomes and intracytoplasmic melanin granules). At each time point (1, 14, and 72 h) after injection of 14C-I-BZA amine, an average of 170 measurements was made.
The 14C/13C ratio gradually fell in tumoral melanosomes, from a mean value of 13 × 10−5 (±12 × 10−5) at the first hour to 10 × 10−5 (±6 × 10−5) at 14 h and then 3.6 × 10−5 (±2.3 × 10−5) at 72 h. On the basis of these data, the estimated retention half-life of 14C-I-BZA in tumoral melanosomes was about 38 h.
DISCUSSION
Site of I-BZA Retention
This study settles the controversy over the I-BZA retention site in melanoma cells—that is, membrane σ-1 receptors (20–22,30) or melanin (23,24,32). The high I-BZA concentration in the cytoplasm of normal and tumoral melanocytes and its pattern of distribution on SIMS images clearly favor the second hypothesis. Thus, when melanin is present, the melanin binding is the one apparent. Any specific binding to σ-membrane receptors would contribute little to the total amount of I-BZA retained in melanomas or normal pigmented tissues. We had early raised the hypothesis that I-BZA was specifically located in melanin granules (31). In that earlier work, however, we had to combine 2 microanalytic techniques: SIMS analysis with the low-resolution ion microscope SMI-300, and electron probe x-ray microanalysis. The SMI-300 had shown that I-BZA was specifically located in the choroidal melanocytes and retinal pigment cells. Then, when the beam of the electron microscope was focused on various intracellular structures, the characteristic Lα1 x-rays of iodine were specifically detected in the melanin granules of the melanosomes (31). The results obtained here with the better-resolution IMS-4F ion microscope on the eye, skin, and melanoma tissues corroborate our previous findings. Further improvement in spatial resolution is now possible with the advent of a new generation of ion microscopes, such as the NanoSIMS-50 (CAMECA) (35), as borne out during work on a prototype (10,11) and also during preliminary work on the commercialized instrument (36).
Melanins are polymers synthesized by melanocytes (34). Many compounds accumulate in pigmented tissues, either by incorporation in nascent melanin (34) or by binding to preformed melanin (37). The significant accumulation of I-BZA observed here, not only in melanoma cells but also in melanoma-infiltrating macrophages (melanophages), as early as 1 h after injection favors binding to preformed melanin. Indeed, melanophages do not synthesize melanin but phagocytose melanin pigment and melanosome bodies released by melanocytes. The presence of I-BZA in the pigmented layers of the retina also supports this view, because this tissue contains mainly mature melanosomes, with little or no melanin turnover after birth (34).
Comparison of 127I and 14C Images
SIMS offered far more information than did a simple distribution map. The distribution of 127I and that of the 2 I-BZA species labeled with 14C at different molecular sites were identical, indicating the stability of the amide bond of I-BZA. To our knowledge, this is the first SIMS work assessing the intracellular metabolic state of a molecule by comparing images of atoms present at different sites of the studied molecule.
As with any tracer study, one should consider the possibility of redistribution of the compound during the fixation steps. However, the images obtained of various tissues (normal and tumoral), consistently showing I-BZA uptake by intracytoplasmic melanosomes, strongly suggest a specific uptake mechanism. Moreover, our results agree with those of other authors, who used either cell cultures (23) or centrifugation techniques (24).
Some authors have applied cryogenic techniques successfully to SIMS using, for example, a cryogenic sandwich-fracture method for cell cultures followed by freeze-drying (5).
Quantitative Cellular Data
The 14C/13C ratio can be used to study concentration variations in different cells and to calculate the biologic half-life at retention sites. Our measurements of the 14C/13C ratio in samples taken at different times after 14C-I-BZA injection yielded a calculated biologic half-life of 38 h in tumor-cell melanosomes.
SIMS and Microdosimetry
When presenting MIRD pamphlet no. 17, Bolch et al. underlined the limitations of macroscopic dosimetry, especially in applications such as internal cancer radiotherapy (1). Indeed, the therapeutic efficacy of new radiopharmaceuticals can be difficult to predict with this method because of a lack of information on the precise tissue distribution.
The digital images provided by SIMS can be used directly to assess the distribution of the radiopharmaceutical as basic data for dose calculations on the cell and tissue level. We found that I-BZA was present both in tumor-cell melanocytes and in melanophages. The latter are less numerous but contain higher I-BZA concentrations. Monte Carlo techniques can now be used to determine the dose distribution for a given isotope, taking into account the distribution as imaged by SIMS (2,38,39).
SIMS circumvents certain limitations of microautoradiography, such as the difficulty of matching the position in the emulsion with the cellular site of radioactive emission, the difficulty of assessing local concentrations (yield quantification is imprecise in microautoradiography), and the poor sensitivity of microautoradiography for 14C (physical half-life, 5,730 y).
Implications of Our Results
Although our results cannot be directly extrapolated to humans, they show that I-BZA may be a useful compound for internal radiotherapy. I-BZA is incorporated by melanoma tumor tissue, within melanocytes and melanophages. Its half-life at its cellular retention site was approximately 38 h in the model used here. Thus, internal radiotherapy seems to be a promising approach for melanoma treatment.
The I-BZA uptake ratio between tumoral and healthy tissues (blood, muscle, brain, lung, and liver) is high (15,18). However, because I-BZA is also taken up by the melanocytes of normal pigmented tissues, studies of the possible impact on these tissues will be needed.
CONCLUSION
This study contributes to the debate on the retention site of I-BZA, a compound of potential interest for the imaging and treatment of malignant melanoma. Our results also show that ionic microscopy (SIMS) is a potent tool for the development and assessment of labeled and unlabeled radiopharmaceuticals.
Acknowledgments
We thank Pr. Georges Slodzian and Royer Dennebouy for having hosted imaging studies on the IMS-4F ion microscope, installed at Laboratoire de Physique des Solides (Paris XI University).
Footnotes
Received Feb. 7, 2005; revision accepted Jun. 9, 2005.
For correspondence contact: Elif Hindie, MD, PhD, Service de Médecine Nucléaire, Hôpital Saint Antoine, 184 rue du Faubourg Saint Antoine, 75012 Paris, France.
E-mail: elif.hindie{at}sat.ap-hop-paris.fr
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Dose Deposits from 90Y, 177Lu, 111In, and 161Tb in Micrometastases of Various Sizes: Implications for Radiopharmaceutical Therapy
- Lognormal Distribution of Cellular Uptake of Radiopharmaceuticals: Implications for Biologic Response in Cancer Treatment
- Reply: 131I Radiation Dose Distribution in Metastases of Thyroid Carcinoma
- CELLDOSE: A Monte Carlo Code to Assess Electron Dose Distribution S Values for 131I in Spheres of Various Sizes
- Preparation and Biologic Evaluation of a Novel Radioiodinated Benzylpiperazine, 123I-MEL037, for Malignant Melanoma